Каспарская полоса
Каспарова полоска — лентообразное утолщение в центре корня эндодермы (радиальной и тангенциальной стенок клеток энтодермы) сосудистых растений ( Pteridophytes [1] и сперматофиты ). В состав региона входят в основном суберин , лигнин и некоторые структурные белки, которые способны уменьшать диффузионный апопластный поток воды и растворенных веществ в стелу , а его ширина варьируется у разных видов. [2] [3] Каспарова полоска непроницаема для воды, поэтому может контролировать транспортировку воды и неорганических солей между корой и сосудистым пучком , предотвращая транспортировку воды и неорганических солей к стеле через апопласт , так что они должны проникать в клеточную мембрану и перемещаться. к стеле через симпластический путь , блокируя внутренние и внешние объекты клетки. [ нужны разъяснения ] Функция массового транспорта аналогична функции тканей животных. [ нужны разъяснения ] . [4] [5] Развитие полоски Каспария регулируется такими факторами транскрипции, как SHORT-ROOT (SHR), SCARECROW (SCR) и MYB36 , а также полипептидным гормоном, синтезируемым клетками среднего столбика. [6] [7]


Химический состав полоски Каспарова был описан как состоящий из суберина . Согласно некоторым исследованиям, [8] полоса Каспарова начинается с локализованного отложения фенольных и ненасыщенных жировых веществ в средней пластинке между радиальными стенками в виде частично окисленных пленок. Первичная стенка покрывается коркой, а затем утолщается отложениями подобных веществ на внутренней стороне этой стенки. Покрытие клеточной стенки материалом, составляющим полоску Каспарова, предположительно закупоривает поры, которые в противном случае позволяли бы воде и питательным веществам перемещаться по этому пути за счет капиллярного действия . [9] Цитоплазма поэтому она не может легко отделиться от полоски , энтодермальной клетки прочно прикреплена к полоске Каспарова , когда клетки подвергаются сокращению протопластов . В корне полоска Каспарова встроена в клеточную стенку энтодермальных клеток в нерастущей области корня за кончиком корня. [10] Здесь полоска Каспарова служит пограничным слоем, отделяющим апопласт коры от апопласта сосудистой ткани, тем самым блокируя диффузию материала между ними. [11] Это разделение заставляет воду и растворенные вещества проходить через плазматическую мембрану симпластическим путем, чтобы пересечь слой эндодермы. [10]
роста коры Полоски Каспарова дифференцируются после завершения наружу. На этом уровне развития корня первичная ксилема его сосудистого цилиндра развита лишь частично. У голосеменных и покрытосеменных растений, имеющих вторичный рост, в корнях обычно развивается только эндодерма с каспариановыми полосками. У многих из них эндодерма позже отторгается вместе с корой, когда развивается перидерма из перицикла . Если перицикл поверхностный и кора сохранена, то энтодерма либо растягивается, либо сдавливается, либо идет в ногу с расширением сосудистого цилиндра за счет радиальных антиклинальных делений, и на новых стенках развиваются каспариевые полоски в продолжение старых. [12]
В отсутствие вторичного роста (большинство однодольных и несколько эвдикотов ) эндодерма обычно подвергается модификациям стенок. Помимо освоения Каспарийской полосы, существуют две стадии развития. На второй стадии суберин (или энтодерма [9] ) покрывает всю стенку внутри клетки. В результате полоска Каспарова отделяется от цитоплазмы, и связь между ними перестает быть очевидной. На третьей стадии целлюлозы над суберином откладывается толстый слой , иногда преимущественно на внутренних тангенциальных стенках. Утолщенная стенка, как и исходная стенка, в которой расположена полоска Каспарова, может одревеснеть, образуя вторичную клеточную стенку. Каспарову полоску можно идентифицировать после того, как произошло утолщение стенки энтодермы. Утолщенная стенка эндодермы может иметь ямки. У однодольных отчетливо выражено последовательное развитие стенок эндодермы. [13] [14]
Открытие
[ редактировать ]Открытие Каспаровой полосы датируется серединой XIX века и продвинуло вперед понимание эндодермы корней растений. [15] В 1865 году немецкий ботаник Роберт Каспари впервые описал энтодерму корня растений, обнаружил, что ее клеточная стенка утолщена, и назвал ее Шухтцшейде. Позже ученые назвали его утолщенную часть «Поясом Карла», названным в честь Кэсбери. [ нужны разъяснения ] . [5] [16] Термин «Caspary'schen fleck» ( нем . Caspary'schen fleck ) появился в литературе 1870-х годов, [17] [18] а после 20 века ее часто называли полосой Каспарова. В 1922 году исследователи впервые оставили каспарианскую полоску от корня растения, чтобы изучить ее состав. [ нужны разъяснения ] [19] [20]
Состав
[ редактировать ]Химический состав Каспарийской полосы долгое время вызывал споры. Кэсбери указал, что эта структура может состоять из лигнина или суберина . Более поздние ученые в основном думали, что это суберин. [21] Лишь в 1990-х годах после анализа Каспаровой полосы нескольких растений было обнаружено, что основным компонентом является лигнин, но многие учебники не обновлялись. [4] Хотя клеточная стенка эндотелия богата вудболитами , это является результатом подуровневой дифференцировки эндотелия. [примечание 1] Раньше некоторые ученые считали, что формирование эндодермы Каспаровой полоски является началом подуровневой дифференциации, однако прямой связи между ними нет. Каспарова полоска образовалась после первичной дифференцировки, а вторичная дифференцировка начинается с косого среза корня, а не там, где находится Каспарова полоска. [1]
Функция
[ редактировать ]Каспарова полоска полностью заполнена промежутком между эндотелиальными клетками, включая среднюю пластинку , в результате чего клеточные стенки двух клеток практически слиты. [1] При транспортировке воды и неорганических питательных веществ в корне растений полоска Каспарова влияет преимущественно на транспорт первичных in vitro, то есть на транспорт воды и неорганических солей через интерстициальные клетки эпидермиса и клетки коры. Когда вода и неорганическая соль поступают к эндотелиальным клеткам, им необходимо проникнуть в клетку через клеточную мембрану , поскольку каспариева полоска не является водопроницаемой, а затем транспортироваться по внутреннему пути протоплазмы, чтобы достичь лигнановых клеток стелы , а затем к другим органам, таким как стебли и листья. [16] При неблагоприятной среде роста каспариановая полоска может выступать в качестве барьера между растительными клетками и внешним миром, предотвращая попадание ионов или отток собственных ионов в окружающую среду. [7] Кроме того, утолщение пояса тушки и коры также предотвращает проникновение токсичных веществ или патогенов, а также выполняет функцию предотвращения рассеивания воды. [22] Некоторые исследования показали, что растения могут образовывать более толстую полосу Каспария в среде с высоким содержанием соли и на участках ближе к кончикам корней, что может быть адаптацией к окружающей среде. [23] [24] но по сравнению с эндотелиальным подуровнем, дифференцированным деревянными болтовыми стенками, которые значительно утолщаются при высоком солевом воздействии, изменения Каспаровой полосы меньшие. [25]
Каспарова полоска расположена преимущественно в эндодерме корня. [26] но у некоторых растений также есть полоска Каспарова во внешней коре на внешней стороне коры корня, стебля или листа . [27] Например, хвойные деревья Pinus Bungeana и стебли пеларгонии имеют каспариевую полосу, что может быть связано с предотвращением распространения воды или проникновения патогенов. [28] [29]
Разработка
[ редактировать ]
Развитие Каспаровой полоски начинается после полной задержки эндогенных клеток, [21] [30] и в настоящее время существует два новостных сигнала трансдукции, которые способствуют образованию эндотелиальных клеток полоски Каспарова. Первым является транскрипционный фактор Short-root (SHR). Активируются дополнительные два транскрипционных фактора Myb36 и Scarecrow (SCR), первый может стимулировать белки полос Каспари (Casp1-5), пероксидазу (PER64) и ESB1 (улучшенный) суберин 1) и т. д. ., последнее влияет на положение полоски Каспарова во внутренней клетке кожи, из-за чего положение полоски Каспарова оказывается слишком близким к стеле; [6] второй - средний фактор целостности полоски Каспария (CIF1-2) и рецепторы GSO1 (SGN3) и GSO2, связанные с радиальной стенкой эндотелиальных клеток, и рецептор GSO2 в боковой стенке. CASP в клетках концентрируется в области клеточной мембраны, соответствующей положению полоски Каспарова, образуя мембранный домен полоски Каспария (CSD), и CSD включается в эту область. Рецептор GS01 окружен краем каждой области CSD, способствуя слиянию CSD в область непрерывной полоски, то есть область, где должна формироваться полоска Каспарова. [7] [31]
Белок полоски Каспария представляет собой мембранный белок , который взаимодействует друг с другом и может связываться с белками, необходимыми для синтеза лигнина, такими как PER64, ESB1 и гомолог F респираторной оксидазы (RBOHF), для активации последующей реакции развития полоски Каспария. [1] [5] У мутантных растений, лишенных рецепторов GSO1 или одновременно лишенных полипептидов CIF1 и CIF2, CASP1 аномально распределяется на мембране эндотелиальных клеток, и CSD не может нормально сливаться в непрерывную и полную полосчатую структуру, таким образом, в конечном итоге образуя разорванную и прерывистую полоску Каспарова. . [7] [31]
Факторы окружающей среды, такие как свет, засоленность почвы и дефицит воды, могут повлиять на развитие Каспарийской полосы. [28]
Фото
[ редактировать ] Сосудистые пучки корня рода Acorus ( однодольные ) можно увидеть в эндотелии и каспаревой полоске вокруг среднего столбика. Полоса Каспарова окрашена в красный цвет, поскольку содержит лигнин.
Сосудистые пучки корня рода Acorus ( однодольные ) можно увидеть в эндотелии и каспаревой полоске вокруг среднего столбика. Полоса Каспарова окрашена в красный цвет, поскольку содержит лигнин. В сосудистом пучке корня рода Ranunculus ( двудольных ) можно увидеть эндотелий и каспариевую полоску вокруг среднего столбика. Полоса Каспарова окрашена в красный цвет, поскольку содержит лигнин.
В сосудистом пучке корня рода Ranunculus ( двудольных ) можно увидеть эндотелий и каспариевую полоску вокруг среднего столбика. Полоса Каспарова окрашена в красный цвет, поскольку содержит лигнин.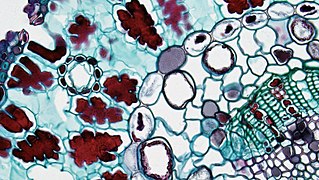 Хвойные листья сосновых голосеменных ( ) . Периферическая эндотелиальная эндодерма и полоска Каспарова видимых сосудистых пучков.
Хвойные листья сосновых голосеменных ( ) . Периферическая эндотелиальная эндодерма и полоска Каспарова видимых сосудистых пучков.



См. также
[ редактировать ]Примечания
[ редактировать ]- ^ После того, как стенка эндотелиальных клеток в старом корне образует утолщенную древесную эмболию, ее функция может быть изменена с передачи воды на защиту растений, что может еще больше ограничить передачу воды и неорганических солей. Только канальные клетки (несколько подуровневых дифференцированных эндотелиальных клеток) сохраняют транспортную функцию. По мере роста корня некоторые растения теряют канальные клетки в корне. [1]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Гельднер, Н. (2013). «Энтодерма». Ежегодный обзор биологии растений . 64 (1): 531–558. doi : 10.1146/annurev-arplant-050312-120050 . ISSN 1543-5008 . ПМИД 23451777 .
- ^ Чен, Тонг; Цай, Ся; Ву, Сяоцинь; Карахара, Ичиро; Шрайбер, Лукас; Линь, Цзиньсин (октябрь 2011 г.). «Развитие Каспарийской полосы и его потенциальная роль в обеспечении солеустойчивости» . Сигнализация и поведение растений . 6 (10): 1499–1502. дои : 10.4161/psb.6.10.17054 . ISSN 1559-2316 . ПМЦ 3256377 . ПМИД 21904117 .
- ^ Киркхэм, М.Б. (01.01.2005), Киркхэм, М.Б. (редактор), «14 - Анатомия корня и закон Пуазейля для потока воды в корнях» , Принципы взаимоотношений почвы и воды в растениях , Берлингтон: Academic Press, стр. 207 –227, ISBN 978-0-12-409751-3 , получено 22 декабря 2022 г.
- ^ Перейти обратно: а б Гельднер, Н. (2013). «Каспарианские полосы» (PDF) . Современная биология . Том. 23, нет. 23. С. R1025, R1026.
- ^ Перейти обратно: а б с Ропполо, Д.; Де Рибель, Б.; Сухожилие, В.Д.; Пфистер, А.; Алассимоне, Дж.; Вермеер, JEM; Ямадзаки, М.; Штирхоф, Ю.-Д.; Бекман, Т.; Гельднер, Н. (2011). «Новое семейство белков опосредует образование каспариановых полосок в эндодерме». Природа . 473 (7347): 380–383. Бибкод : 2011Natur.473..380R . дои : 10.1038/nature10070 . ISSN 0028-0836 . ПМИД 21593871 . S2CID 4366553 .
- ^ Перейти обратно: а б Ли, П.; Ю, К.; Гу, Х.; Сюй, К.; Ци, С.; Ван, Х.; и др. (2018). «Построение функциональной полоски Каспарова в неэнтодермальных линиях управляется двумя параллельными сигнальными системами у Arabidopsis thaliana» . Современная биология . 28 (17): 2777–2786.e2. дои : 10.1016/j.cub.2018.07.028 . ПМИД 30057307 .
- ^ Перейти обратно: а б с д Накаяма, Т.; Синохара, Х.; Танака, М.; Баба, К.; Огава-Охниши, М.; Мацубаяси, Ю. (2017). «Пептидный гормон, необходимый для формирования диффузионного барьера полос Каспари в корнях арабидопсиса». Наука . 355 (6322): 284–286. Бибкод : 2017Sci...355..284N . дои : 10.1126/science.aai9057 . ПМИД 28104889 . S2CID 206653442 .
- ^ Ван Флит, DS (1961). «Гистохимия и функция эндодермы». Ботанический обзор . 27 (2): 165–220. дои : 10.1007/BF02860082 . S2CID 45393531 .
- ^ Перейти обратно: а б Фрей-Висслинг, А.; Его Святейшество Боссхард (1959). Цитология лучевых клеток заболони и сердцевины . Крам.
- ^ Перейти обратно: а б Тайз Л., Зейгер, Эдуардо, Мёллер, Ян Макс и Мерфи, Ангус. (2015). Физиология и развитие растений (Шестое изд.).
- ^ Гребе, М (2011). «Открытие Каспаровой полосы». Природа . 473 (7347): 294–5. дои : 10.1038/473294a . ПМИД 21593860 . S2CID 873056 .
- ^ фон Гуттенберг, Х. (1943). Физиологические оболочки . Борнтрегер.
- ^ ОГУРА, Ю. (1938). «Задачи морфологии (13)». Ботаника и зоология . 6 : 139–148.
- ^ Напп-Зинн, А.Ф. (1953). 100 лет рейнскому пароходству Кёльн-Дюссельдорф: особенности разрушения и реконструкции 1939-1953 гг . Рейс на пароходе Кёльн-Дюссельдорф по Рейну.
- ^ Сиго, Джей Л. младший (2020). «Возврат к возникновению и обнаружению эндодермы в побегах покрытосеменных» . Флора . 273 : 151709. doi : 10.1016/j.flora.2020.151709 .
- ^ Перейти обратно: а б Гребе, М. (2011). «Открытие Каспаровой полосы». Природа . 473 (7347): 294–295. дои : 10.1038/473294a . ISSN 0028-0836 . ПМИД 21593860 . S2CID 873056 .
- ^ Руссов, Э. (1872). «Rhizocarpeae. I. Осевые органы: А. Ствол: а. Марсилия (Drummondii, elata, salvatrix)» . Сравнительные исследования по вопросам гистиологии (гистиографии и гистиогении) вегетативных и спорообразующих органов и развития спор сосудистых криптогам: с учетом гистиологии фанерогам, начиная с рассмотрения Marsiliaceae . Комиссары Имперской академии наук. стр. 1–12.
- ^ Мюллер, К. (1884). «Морфология тканей» . 12 (1): 234–342.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Пристли, Дж. Х.; Норт, EE (1922). «Физиологические исследования в анатомии растений III. Структура эндодермы в зависимости от ее функции» . Новый фитолог . 21 (3): 113–139. дои : 10.1111/j.1469-8137.1922.tb07593.x . JSTOR 2428118 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Песня, К.; Шен, В.; Ду, Л.; Вэнь, Дж.; Лин, Дж.; Ли, Р. (2019). «Развитие и химическая характеристика полосок Каспария в корнях пихты китайской (Cunninghamia lanceolata)» . Деревья . 33 (3): 827–836. дои : 10.1007/s00468-019-01820-x . ISSN 0931-1890 .
- ^ Перейти обратно: а б Насер, С.; Ли, Ю.; Лапьер, К.; Франке, Р.; Наврат, К.; Гельднер, Н. (2012). «Диффузионный барьер полоски Каспария у Arabidopsis состоит из полимера лигнина без суберина» . Труды Национальной академии наук . 109 (25): 10101–6. Бибкод : 2012PNAS..10910101N . дои : 10.1073/pnas.1205726109 . ПМЦ 3382560 . ПМИД 22665765 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Роббинс, штат Невада; Тронтин, К.; Дуань, Л.; Диннени, младший (2014). «За барьером: связь в корне через эндодерму» . Физиология растений . 166 (2): 551–9. дои : 10.1104/стр.114.244871 . ПМК 4213087 . ПМИД 25125504 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Карахара, И.; Икеда, А.; Кондо, Т.; Уэтаке, Ю. (2004). «Развитие Каспаровой полосы в первичных корнях кукурузы в условиях солевого стресса» . Планта . 219 (1): 41–7. дои : 10.1007/s00425-004-1208-7 . ПМИД 14986139 . S2CID 792699 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Чен, Т.; Цай, X.; Ву, Х.; Карахара, И.; Шрайбер, Л.; Лин, Дж. (2011). «Развитие Каспарийской полосы и его потенциальная роль в обеспечении солеустойчивости» . Сигнализация и поведение растений . 6 (10): 1499–502. дои : 10.4161/psb.6.10.17054 . ПМЦ 3256377 . ПМИД 21904117 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Палмгрен, М. (2018). «Растительный эпителий: какова роль раствора в стене?» . ПЛОС Биология . 16 (12): e3000073. дои : 10.1371/journal.pbio.3000073 . ПМК 6296743 . ПМИД 30517104 .
- ^ Эверт, РФ (28 августа 2006 г.). Анатомия растений Исава: меристемы, клетки и ткани тела растения: их строение, функции и развитие . Джон Уайли и сыновья. п. 6. ISBN 978-0-470-04737-8 . Архивировано из оригинала 01 декабря 2020 г. Проверено 17 апреля 2021 г.
- ^ Лерстен, НР (1997). «Появление эндодермы с каспариановой полоской в стебле и листе». Ботаническое обозрение . 63 (3): 265–272. дои : 10.1007/BF02857952 . ISSN 0006-8101 . S2CID 45859773 .
- ^ Перейти обратно: а б Ву, Х.; Лин, Дж.; Лин, К.; Ван, Дж.; Шрайбер, Л. (2005). «Каспариевые полоски в хвое более проницаемы для растворов, чем энтодермальные транспортные барьеры в корнях Pinus Bungeana» . Физиология растений и клеток . 46 (11): 1799–1808. дои : 10.1093/pcp/pci194 . ISSN 1471-9053 . ПМИД 16170202 .
- ^ Мейер, CJ; Петерсон, Калифорния (2011). «Каспариевые полосы встречаются на перидерме стебля и корня Pelargonium hortorum» . Анналы ботаники . 107 (4): 591–8. дои : 10.1093/aob/mcq267 . ПМК 3064534 . ПМИД 21239408 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Барберон, М. (2017). «Энтодерма как контрольный пункт питательных веществ» . Новый фитолог . 213 (4): 1604–1610. дои : 10.1111/nph.14140 . ПМИД 27551946 .
- ^ Перейти обратно: а б Пфистер, А.; Барберон, М.; Алассимоне, Дж.; Кальмбах, Л.; Ли, Ю.; Вермеер, JEM; и др. (2014). «Рецептороподобный киназный мутант с отсутствующим энтодермальным диффузионным барьером демонстрирует селективные дефекты гомеостаза питательных веществ» . электронная жизнь . 3 : e03115. doi : 10.7554/eLife.03115 . ПМК 4164916 . ПМИД 25233277 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )
- Исав, Кэтрин (1965). Анатомия растений . Джон Уайли и сыновья. п. 767 . ISBN 9780471244554 .