Комплекс МЕЧТА
Партнер по димеризации, RB-подобный комплекс E2F и мультивульвального класса B (DREAM), представляет собой белковый комплекс, ответственный за регуляцию экспрессии генов, зависящей от клеточного цикла . [1] [2] Комплекс эволюционно консервативен, хотя некоторые его компоненты варьируются от вида к виду. У человека ключевыми белками комплекса являются RBL1 (p107) и RBL2 (p130), оба из которых являются гомологами RB (p105) и связывают репрессивные факторы транскрипции E2F E2F4 и E2F5 ; DP1 , DP2 и DP3, партнеры по димеризации E2F; и MuvB, который представляет собой комплекс LIN9 /37/52/54 и RBBP4 . [1]
Открытие
[ редактировать ]Гены, кодирующие комплекс MuvB, были первоначально идентифицированы в результате исследований мутаций с потерей функции у C. elegans . В результате мутации эти гены дали червям множество органов, похожих на вульву, отсюда и название «Muv». и DP1 млекопитающих Были классифицированы три класса генов Muv: гены класса B, кодирующие гомологи RB, E2F , а также другие, такие как LIN-54, LIN-37, LIN-7 и LIN-52, функции которых еще не изучены. [3] [4]
Исследования на клетках фолликула яичников Drosophila melanogaster выявили белковый комплекс, который связывается с многократно амплифицирующими генами хориона. В состав комплекса вошли гены, имеющие близкую гомологию с генами MuvB, такие как Mip130, Mip120 и Mip40. Эти гены Mip были идентифицированы как гомологи генов MuvB LIN9, LIN54 и LIN37 соответственно. [5] Дальнейшие исследования ядерных экстрактов эмбрионов мух подтвердили сосуществование этих белков с другими, такими как гомологи RB Rbf1 и Rbf2, а также с другими, такими как E2f и Dp. Таким образом, белковый комплекс был назван дрозофилы комплексом RBF, E2f2 и Mip (dREAM) . Разрушение комплекса dREAM посредством нокдауна RNAi компонентов комплекса dREAM привело к более высокой экспрессии генов, регулируемых E2f, которые обычно подавляются, что указывает на роль dREAM в подавлении генов. [6] Позже у Drosophila melanogaster был также обнаружен семенниково-специфичный паралог комплекса Myb-MuvB/DREAM, известный как tMAC (testis-специфический комплекс остановки мейоза), который участвует в остановке мейоза. [7]
Белковый комплекс, аналогичный dREAM, был впоследствии идентифицирован в экстракте C. elegans, содержащем DP, RB и MuvB, и получил название DRM. Этот комплекс включал гомологи RB и DP млекопитающих и других членов комплекса MuvB. [8]
Комплекс DREAM млекопитающих был идентифицирован после иммунопреципитации р130 с помощью масс-спектрометрического анализа. Результаты показали, что p130 связан с E2F4, E2F5, партнером по димеризации DP, а также LIN9, LIN54, LIN37, LIN52 и RBBP4, которые составляют комплекс MuvB. Иммунопреципитация факторов MuvB также выявила ассоциацию BMYB. Последующая иммунопреципитация с помощью BMYB дала все основные белки MuvB, но не других членов комплекса DREAM – p130, p107, E2F4/5 и DP. Это указывало на то, что MuvB связывался с BMYB с образованием комплекса BMYB-MuvB или с p130/p107, E2F4/5 и DP с образованием комплекса DREAM. Комплекс DREAM был обнаружен преобладающим в покоящихся или голодающих клетках, а комплекс BMYB-MuvB был обнаружен в активно делящихся клетках, что указывает на отдельные функциональные возможности этих двух комплексов. [9]
Недавно у Arabidoposis были обнаружены MuvB-подобные комплексы , которые включают ортологи E2F и MYB в сочетании с ортологами LIN9 и LIN54. [10] [11]
Функция
[ редактировать ]Основная функция комплекса DREAM заключается в репрессии G1/S и G2/M экспрессии генов во время покоя (G0 ) . Вступление в клеточный цикл отделяет p130 от комплекса и приводит к последующему привлечению активирующих белков E2F. Это позволяет экспрессировать гены поздней фазы G1 и S, регулируемые E2F. BMYB ( MYBL2 ), который репрессируется комплексом DREAM во время G0 , также способен экспрессироваться в это время и связывается с MuvB во время S-фазы , чтобы способствовать экспрессии ключевых генов G2 -фазы , / M таких как CDK1 и CCNB1 . Затем FOXM1 рекрутируется в G2 для дальнейшего стимулирования экспрессии генов (например, AURKA ). Во время поздней фазы S BMYB разрушается посредством CUL1 ( комплекс SCF ), тогда как FOXM1 разрушается во время митоза с помощью APC/C . [1] [12] Ближе к концу клеточного цикла комплекс DREAM повторно собирается с помощью DYRK1A для репрессии генов G1/S и G2/M.
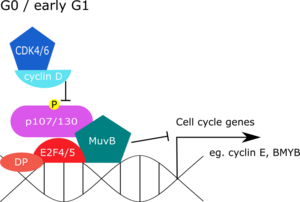
G0
[ редактировать ]В состоянии покоя комплекс DREAM репрессирует экспрессию генов G1/S и G2/M. В системах млекопитающих исследования иммунопреципитации хроматина (ChIP) показали, что компоненты DREAM обнаруживаются вместе на промоторах генов, пик которых приходится на фазу G1/S или G2/M . [9] С другой стороны, отмена комплекса DREAM привела к усилению экспрессии генов, регулируемых E2F, которые обычно репрессируются в фазе G0. [9] [13] В отличие от клеток млекопитающих, комплекс dREAM мух был обнаружен почти в трети всех промоторов, что может отражать более широкую роль dREAM в регуляции генов, например, в запрограммированной гибели нервных клеток-предшественников. [14] [15]
Стыковка комплекса DREAM с промоторами достигается путем связывания LIN-54 с областями, известными как область гомологии генов клеточного цикла (CHR). Это специфическая последовательность нуклеотидов, которая обычно встречается в промоторах генов, экспрессируемых во время поздней фазы S или фазы G2/M. Стыковка также может быть достигнута посредством связывания белков E2F с последовательностями, известными как сайты элементов, зависящих от клеточного цикла (CDE). Были обнаружены некоторые гены, зависящие от клеточного цикла, где CHR и CDE находятся рядом друг с другом. Поскольку p130-E2F4 может образовывать стабильные ассоциации с комплексом MuvB, близость CHR к CDE предполагает, что аффинность связывания комплекса DREAM с генами-мишенями кооперативно улучшается за счет ассоциации с обоими сайтами связывания. [16]
Когда DREAM пристыкован к промотору, p130 связывается с LIN52, и эта ассоциация ингибирует связывание LIN52 с белками-модификаторами хроматина . [17] [18] Следовательно, в отличие от RB-E2F, комплекс DREAM вряд ли напрямую рекрутирует модификаторы хроматина для репрессии экспрессии генов, хотя некоторые ассоциации были предположены. [19] [20] Вместо этого комплекс DREAM может подавлять экспрессию генов, влияя на положение нуклеосом . Уплотненная ДНК в местах начала транскрипции ингибирует экспрессию генов, блокируя стыковку РНК-полимеразы . [21] Например, у червей потеря белка комплекса MuvB, LIN35, приводит к потере репрессивных гистоновых ассоциаций и высокой экспрессии генов, зависимых от клеточного цикла. Однако прямые доказательства связи между репрессивными гистонами и комплексом СОН еще предстоит выяснить. [22]

Г1/С
[ редактировать ]Как и его аналог, RB-E2F, на комплекс DREAM также влияют аналогичные стимулы роста и последующая активность циклин-CDK. Увеличение активности циклина D-CDK4 и циклина E-CDK2 диссоциирует комплекс DREAM от промотора путем фосфорилирования p130. [18] Гиперфосфорилированный р130 впоследствии разрушается. [23] [24] и E2F4 экспортируется из ядра. [25] Как только репрессивные E2F освобождаются, активирующие E2F связываются с промотором, активируя гены G1/S, которые способствуют синтезу ДНК и переходу клеточного цикла. [26] В это время также происходит активация BMYB, который затем связывается с генами, достигающими пика в фазе G2/M. [9] [27] [28] Связывание BMYB с генами позднего клеточного цикла зависит от его ассоциации с ядром MuvB с образованием комплекса BMYB-MuvB, который затем способен активировать гены в фазе G2/M. [12]
Поздний митоз
[ редактировать ]Ближе к концу митоза р130 и р107 дефосфорилируются из гиперфосфорилированного состояния фосфатазой PP2a . [29] [30] Ингибирование активности PP2a снижало связывание промоторов некоторых белков комплекса DREAM в последующей фазе G1 и дерепрессию экспрессии генов. [31]
Было показано, что другие компоненты фосфорилируются для сборки комплекса DREAM. Из них наиболее изучено фосфорилирование LIN52 по остатку S28. Замена этого серина на аланин приводила к снижению связывания ядра MuvB с p130 и нарушала способность клеток входить в состояние покоя. Это указывает на то, что фосфорилирование LIN52 S28 необходимо для правильной ассоциации и функционирования комплекса DREAM посредством связывания с p130. Одним из известных регуляторов фосфорилирования остатка S28 является DYRK1A . Потеря этой киназы приводит к снижению фосфорилирования остатка S28 и ассоциации p130 с MuvB. [13] Также было обнаружено, что DYRK1A разрушает циклин D1, что приводит к повышению уровня p21 – оба из которых способствуют выходу из клеточного цикла. [32]
Также было показано, что комплекс DREAM регулирует цитокинез посредством GAS2L3. [33]
Терапия рака
[ редактировать ]Благодаря его регуляторной роли в клеточном цикле, воздействие на комплекс DREAM может улучшить противораковые методы лечения, такие как иматиниб . [34] [35]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Садашивам, Субхашини; ДеКаприо, Джеймс А. (11 июля 2013 г.). «Комплекс DREAM: главный координатор экспрессии генов, зависящей от клеточного цикла» . Обзоры природы Рак . 13 (8): 585–595. дои : 10.1038/nrc3556 . ПМЦ 3986830 . ПМИД 23842645 .
- ^ Фишер, М; Мюллер, Джорджия (декабрь 2017 г.). «Контроль транскрипции клеточного цикла: комплексы DREAM/MuvB и RB-E2F» . Критические обзоры по биохимии и молекулярной биологии . 52 (6): 638–662. дои : 10.1080/10409238.2017.1360836 . ПМИД 28799433 . S2CID 205695213 .
- ^ Бейтель, Г.Дж.; Ламби, Э.Дж.; Хорвиц, HR (22 августа 2000 г.). «Ген lin-9 C. elegans, который действует на пути, связанном с Rb, необходим для развития клеток оболочки гонад и кодирует новый белок». Джин . 254 (1–2): 253–63. дои : 10.1016/s0378-1119(00)00296-1 . ПМИД 10974557 .
- ^ Томас, Дж. Х.; Сеол, CJ; Шварц, ХТ; Хорвиц, HR (май 2003 г.). «Новые гены, которые взаимодействуют с lin-35 Rb, негативно регулируют путь ras let-60 у Caenorhabditis elegans» . Генетика . 164 (1): 135–51. дои : 10.1093/генетика/164.1.135 . ПМЦ 1462563 . ПМИД 12750327 .
- ^ Билл, Эл.; Манак, младший; Чжоу, С; Белл, М; Липсик, Дж. С. ; Ботчан, MR (19 декабря 2002 г.). «Роль белкового комплекса, содержащего Myb дрозофилы, в сайт-специфической репликации ДНК». Природа . 420 (6917): 833–7. Бибкод : 2002Natur.420..833B . дои : 10.1038/nature01228 . ПМИД 12490953 . S2CID 4425307 .
- ^ Кореняк, М; Тейлор-Хардинг, Б; Бинне, Великобритания; Саттерли, Дж. С.; Стево, О; Осланд, Р; Уайт-Купер, Х; Дайсон, Н.; Брем, А. (15 октября 2004 г.). «Нативные комплексы E2F/RBF содержат белки, взаимодействующие с Myb, и подавляют транскрипцию генов-мишеней E2F, контролируемых развитием» . Клетка . 119 (2): 181–93. дои : 10.1016/j.cell.2004.09.034 . ПМИД 15479636 . S2CID 17989678 .
- ^ Билл, Эл.; Льюис, ПВ; Белл, М.; Роча, М.; Джонс, Д.Л.; Ботчан, М.Р. (15 апреля 2007 г.). «Открытие tMAC: специфичный для семенников мейотический комплекс ареста дрозофилы, паралогичный Myb-Muv B» . Гены и развитие . 21 (8): 904–919. дои : 10.1101/gad.1516607 . ПМК 1847709 . ПМИД 17403774 .
- ^ Харрисон, ММ; Сеол, CJ; Лу, Х; Хорвиц, HR (07 ноября 2006 г.). «Некоторые синтетические белки multivulva класса B C. elegans кодируют консервативный LIN-35 Rb-содержащий комплекс, отличный от NuRD-подобного комплекса» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (45): 16782–7. Бибкод : 2006PNAS..10316782H . дои : 10.1073/pnas.0608461103 . ПМК 1636532 . ПМИД 17075059 .
- ^ Перейти обратно: а б с д Литовчик Л ; Садасивам, С; Флоренция, Л; Чжу, Х; Суонсон, СК ; Велмуруган, С; Чен, Р; Уошберн, член парламента; Лю, XS; ДеКаприо, Дж.А. (25 мая 2007 г.). «Эволюционно консервативный мультисубъединичный белковый комплекс RBL2/p130 и E2F4 подавляет гены, зависящие от клеточного цикла человека, в состоянии покоя» . Молекулярная клетка . 26 (4): 539–51. doi : 10.1016/j.molcel.2007.04.015 . ПМИД 17531812 .
- ^ Кобаяши, К; Сузуки, Т; Ивата, Э; Накамичи, Н; Сузуки, Т; Чен, П; Отани, М; Исида, Т; Хосоя, Х; Мюллер, С; Левицкий, Т; Петтко-Сандтнер, А; Дарула, З; Ивамото, А; Номото, М; Тада, Ю; Хигасияма, Т; Демура, Т; Дунан, Дж. Х.; Хаузер, Монтана; Сугимото, К; Умеда, М; Мадьяр, З; Бёгре, Л; Ито, М (4 августа 2015 г.). «Транкрипционная репрессия белками MYB3R регулирует рост органов растений» . Журнал ЭМБО . 34 (15): 1992–2007. дои : 10.15252/embj.201490899 . ПМЦ 4551348 . ПМИД 26069325 .
- ^ Ланг, Лукас; Петтко-Сандтнер, Аладар; Тунчай Эльбаши, Хасибе; Такацука, Хиротомо; Номото, Юджи; Заки, Ахмад; Дорохов, Стефан; Де Ягер, Герт; Экхаут, Доминик; Ито, Масаки; Мадьяр, Золтан; Бёгре, Ласло; Хиз, Марен; Шнитгер, Арп (декабрь 2021 г.). «Комплекс DREAM подавляет рост в ответ на повреждение ДНК арабидопсиса» . Альянс наук о жизни . 4 (12): e202101141. дои : 10.26508/lsa.202101141 . ПМК 8500230 . ПМИД 34583930 .
- ^ Перейти обратно: а б Садасивам, С.; Дуань, С.; ДеКаприо, JA (5 марта 2012 г.). «Комплекс MuvB последовательно вовлекает B-Myb и FoxM1 для стимулирования митотической экспрессии генов» . Гены и развитие . 26 (5): 474–489. дои : 10.1101/gad.181933.111 . ПМК 3305985 . ПМИД 22391450 .
- ^ Перейти обратно: а б Литовчик, Л; Флоренс, Луизиана; Суонсон, СК; Уошберн, член парламента; ДеКаприо, Дж.А. (15 апреля 2011 г.). «Протеинкиназа DYRK1A способствует покою и старению посредством сборки комплекса DREAM» . Гены и развитие . 25 (8): 801–13. дои : 10.1101/gad.2034211 . ПМК 3078706 . ПМИД 21498570 .
- ^ Джорджетт, Д; Ан, С; Макэлпайн, DM; Чунг, Э; Льюис, ПВ; Билл, Эл.; Белл, СП; Скорость, Т; Манак, младший; Ботчан, MR (15 ноября 2007 г.). «Исследования геномного профилирования и экспрессии выявили как положительную, так и отрицательную активность комплекса Myb MuvB/dREAM дрозофилы в пролиферирующих клетках» . Гены и развитие . 21 (22): 2880–96. дои : 10.1101/gad.1600107 . ПМК 2049191 . ПМИД 17978103 .
- ^ Ровани, Маргритт К.; Брахманн, Кэрри Бейкер; Рамзи, Гэри; Катцен, Алиса Л. (декабрь 2012 г.). «Комплекс dREAM/Myb-MuvB и Grim являются ключевыми регуляторами запрограммированной гибели нервных клеток-предшественников на заднем крае крыла дрозофилы» . Биология развития . 372 (1): 88–102. дои : 10.1016/j.ydbio.2012.08.022 . ПМК 3621911 . ПМИД 22960039 .
- ^ Мюллер, Джорджия; Энгеланд, К. (февраль 2010 г.). «Центральная роль элементов промотора CDE/CHR в регуляции транскрипции генов, зависимой от клеточного цикла». Журнал ФЭБС . 277 (4): 877–93. дои : 10.1111/j.1742-4658.2009.07508.x . ПМИД 20015071 . S2CID 8955433 .
- ^ Форристал, К; Хенли, ЮАР; Макдональд, Дж.И.; Буш-младший; Орт, С; Пассос, DT; Таллури, С; Ишак, Калифорния; Туэйтс, MJ; Норли, CJ; Литовчик, Л; ДеКаприо, Дж.А.; ДиМаттиа, Г; Холдсворт, Д.В.; Бейер, Ф; Дик, ФА (июнь 2014 г.). «Потеря комплекса DREAM у млекопитающих нарушает регуляцию пролиферации хондроцитов» . Молекулярная и клеточная биология . 34 (12): 2221–34. дои : 10.1128/MCB.01523-13 . ПМК 4054284 . ПМИД 24710275 .
- ^ Перейти обратно: а б Гайли, Казахстан; Либан, ТиДжей; Фельтхаузен, Дж.Г.; Раманан, П; Литовчик, Л; Рубин, С.М. (01 мая 2015 г.). «Структурные механизмы сборки и регуляции комплекса СОН» . Гены и развитие . 29 (9): 961–74. дои : 10.1101/gad.257568.114 . ПМК 4421984 . ПМИД 25917549 .
- ^ Сандовал, Р; Пилкинтон, М; Коламоничи, Орегон (15 октября 2009 г.). «Удаление p107/p130-связывающего домена Mip130/LIN-9 обходит требование активности CDK4 для диссоциации Mip130/LIN-9 от комплекса p107/p130-E2F4» . Экспериментальные исследования клеток . 315 (17): 2914–20. дои : 10.1016/j.yexcr.2009.07.014 . ПМЦ 2757496 . ПМИД 19619530 .
- ^ Стиглер, П; Де Лука, А; Багелла, Л; Джордано, А (15 ноября 1998 г.). «СООН-концевая область pRb2/p130 связывается с гистондеацетилазой 1 (HDAC1), усиливая репрессию транскрипции E2F-зависимого промотора циклина А». Исследования рака . 58 (22): 5049–52. ПМИД 9823308 .
- ^ Залог; Морозов А.В. (ноябрь 2010 г.). «Регуляция генов путем расположения нуклеосом». Тенденции в генетике . 26 (11): 476–83. дои : 10.1016/j.tig.2010.08.003 . ПМИД 20832136 .
- ^ Латорре, я; Чесни, Массачусетс; Гарригес, Ж.М.; Стемпор, П; Апперт, А; Франческони, М; Стром, С; Арингер, Дж (01 марта 2015 г.). «Комплекс DREAM стимулирует генное тело H2A.Z для целевой репрессии» . Гены и развитие . 29 (5): 495–500. дои : 10.1101/gad.255810.114 . ПМК 4358402 . ПМИД 25737279 .
- ^ Тедеско, Д; Лукас, Дж; Рид, С.И. (15 ноября 2002 г.). «Родственный pRb белок p130 регулируется посредством фосфорилирования-зависимого протеолиза с помощью протеинубиквитинлигазы SCF(Skp2)» . Гены и развитие . 16 (22): 2946–57. дои : 10.1101/gad.1011202 . ПМК 187481 . ПМИД 12435635 .
- ^ Бхаттачарья, С; Гаррига, Дж; Кальбо, Дж; Йонг, Т; Хейнс, Д.С.; Гранья, X (24 апреля 2003 г.). «SKP2 связывается с p130 и ускоряет убиквитилирование и деградацию p130 в клетках человека» . Онкоген . 22 (16): 2443–51. дои : 10.1038/sj.onc.1206339 . ПМИД 12717421 . S2CID 26125392 .
- ^ Гаубац, С; Лиз, Дж.А.; Линдеман, Дж.Дж.; Ливингстон, DM (февраль 2001 г.). «E2F4 экспортируется из ядра CRM1-зависимым образом» . Молекулярная и клеточная биология . 21 (4): 1384–92. дои : 10.1128/MCB.21.4.1384-1392.2001 . ПМК 99590 . ПМИД 11158323 .
- ^ Такахаши, Ю; Рэйман, Дж.Б.; Динлахт, Б.Д. (1 апреля 2000 г.). «Анализ связывания промотора семействами E2F и pRB in vivo: различные белки E2F опосредуют активацию и репрессию» . Гены и развитие . 14 (7): 804–16. дои : 10.1101/gad.14.7.804 . ПМК 316494 . PMID 10766737 .
- ^ Лам, EW; Робинсон, К; Уотсон, Р.Дж. (сентябрь 1992 г.). «Характеристика и регулируемая клеточным циклом экспрессия мышиного B-myb». Онкоген . 7 (9): 1885–90. ПМИД 1501895 .
- ^ Пилкинтон, М; Сандовал, Р; Сонг, Дж; Несс, ЮАР; Коламоничи, Орегон (5 января 2007 г.). «Mip/LIN-9 регулирует экспрессию B-Myb и индукцию циклина А, циклина B и CDK1» . Журнал биологической химии . 282 (1): 168–75. дои : 10.1074/jbc.M609924200 . ПМИД 17098733 . S2CID 21963932 .
- ^ Колупаева, В; Янссенс, В. (январь 2013 г.). «Фосфатазы PP1 и PP2A — сотрудничающие партнеры в модуляции активации белка ретинобластомы» . Журнал ФЭБС . 280 (2): 627–43. дои : 10.1111/j.1742-4658.2012.08511.x . ПМИД 22299668 . S2CID 46705471 .
- ^ Куримчак, А; Гранья, X (2015). «PP2A: больше, чем просто переключатель сброса для активации белков pRB во время клеточного цикла и в ответ на сигнальные сигналы» . Клеточный цикл . 14 (1): 18–30. дои : 10.4161/15384101.2014.985069 . ПМЦ 4612414 . ПМИД 25483052 .
- ^ Наэтар, Н; Саундарапандиан, В; Литовчик Л ; Гоген, КЛ; Саблина А.А.; Боуман-Колин, К; Сичинский, П; Хан, туалет; ДеКаприо, Дж.А.; Ливингстон, DM (19 июня 2014 г.). «PP2A-опосредованная регуляция передачи сигналов Ras в G2 необходима для стабильного покоя и нормальной длины G1» . Молекулярная клетка . 54 (6): 932–45. doi : 10.1016/j.molcel.2014.04.023 . ПМК 4118046 . ПМИД 24857551 .
- ^ Чен, JY; Лин, младший; Цай, ФК; Мейер, Т. (10 октября 2013 г.). «Дозировка Dyrk1a сдвигает клетки внутри сигнальной карты p21-циклин D1, чтобы контролировать решение о вступлении в клеточный цикл» . Молекулярная клетка . 52 (1): 87–100. doi : 10.1016/j.molcel.2013.09.009 . ПМК 4039290 . ПМИД 24119401 .
- ^ Уолтер, П; Шмитт, К; Факлер, М; Кремлинг, Х; Пробст, Л; Хаузер, С; Грусс, О.Дж.; Гаубац, С. (15 мая 2012 г.). «GAS2L3, ген-мишень комплекса DREAM, необходим для правильного цитокинеза и стабильности генома» . Журнал клеточной науки . 125 (Часть 10): 2393–406. дои : 10.1242/jcs.097253 . ПМИД 22344256 .
- ^ ДеКаприо, Джеймс А.; Дуэнсинг, Анетт (июль 2014 г.). «Комплекс DREAM в противоопухолевой активности мезилата иматиниба при стромальных опухолях желудочно-кишечного тракта» . Современное мнение в онкологии . 26 (4): 415–421. doi : 10.1097/CCO.0000000000000090 . ПМЦ 4236229 . ПМИД 24840522 .
- ^ Бойчук С. ; Парри, Дж.А.; Макиельски, КР ; Литовчик Л. ; Барон, JL; Зеве, Япония; Возняк, А.; Мехалек, КР; Корженевский, Н.; Сухожилия, СД; Шоффски, П.; Дебиец-Рихтер, М.; ДеКаприо, Дж.А.; Дуэнсинг, А. (20 июня 2013 г.). «Комплекс DREAM опосредует покой клеток GIST и является новой терапевтической мишенью для усиления апоптоза, индуцированного иматинибом» . Исследования рака . 73 (16): 5120–5129. дои : 10.1158/0008-5472.CAN-13-0579 . ПМИД 23786773 .