Гетероциста

A–F: Носток обыкновенный G–H: Носток кальцикола
I–M: Tolypothrix Distorta N–R: Scytonema hyalinum.
Масштабная линейка = 10 мкм. Сокращения: hc - гетероциста, ак - акинете, hm - гормогоний, nd - некридии.
Гетероцисты или гетероциты — специализированные азотфиксирующие клетки, образующиеся при азотном голодании некоторыми нитчатыми цианобактериями , такими как Nostoc , Cylindrospermum и Anabaena . [1] Они фиксируют азот из динитрогена (N 2 ) в воздухе с помощью фермента нитрогеназы , чтобы обеспечить клетки нити азотом для биосинтеза. [2]
Нитрогеназа инактивируется кислородом, поэтому гетероцисте необходимо создать микроанаэробную среду. гетероцист Уникальная структура и физиология требуют глобального изменения экспрессии генов . Например, гетероцисты:
- производят три дополнительные клеточные стенки , в том числе одну из гликолипидов , образующих гидрофобный барьер для кислорода.
- вырабатывают нитрогеназу и другие белки, участвующие в фиксации азота.
- разрушать фотосистему II , которая производит кислород
- активировать гликолитические ферменты
- производят белки, которые поглощают оставшийся кислород
- содержат полярные пробки, состоящие из цианофицина , который замедляет диффузию от клетки к клетке.
Цианобактерии обычно получают фиксированный углерод (углевод) путем фотосинтеза . Отсутствие расщепления воды в фотосистеме II не позволяет гетероцистам осуществлять фотосинтез, поэтому вегетативные клетки снабжают их углеводами , которыми, как полагают, является сахароза . Обмен фиксированными источниками углерода и азота осуществляется через каналы между клетками нити. Гетероцисты поддерживают фотосистему I , что позволяет им генерировать АТФ путем циклического фотофосфорилирования .
Одиночные гетероцисты развиваются примерно через каждые 9–15 клеток, образуя одномерный узор вдоль нити. Интервал между гетероцистами остается примерно постоянным, хотя клетки нити делятся. Бактериальную нить можно рассматривать как многоклеточный организм с двумя различными, но взаимозависимыми типами клеток. Такое поведение весьма необычно для прокариот и, возможно, было первым примером формирования многоклеточного паттерна в эволюции . Образовавшаяся гетероциста не может вернуться в вегетативную клетку. Некоторые бактерии, образующие гетероцисты, могут дифференцироваться в спороподобные клетки, называемые акинетами, или подвижные клетки, называемые гормогониями , что делает их наиболее фенотипически универсальными из всех прокариот.
Экспрессия генов
[ редактировать ]
В среде с низким содержанием азота дифференцировка гетероцист запускается регулятором транскрипции NtcA. NtcA влияет на дифференцировку гетероцист посредством сигнальных белков, участвующих в процессе дифференцировки гетероцист. Например, NtcA контролирует экспрессию нескольких генов, включая HetR, который имеет решающее значение для дифференцировки гетероцист. [3] Это имеет решающее значение, поскольку повышает регуляцию других генов, таких как hetR, patS, hepA, связываясь с их промотором и, таким образом, действуя как фактор транскрипции . Также стоит отметить, что экспрессия ntcA и HetR зависят друг от друга и их присутствие способствует дифференцировке гетероцист даже в присутствии азота. Недавно также было обнаружено, что другие гены, такие как PatA, hetP, регулируют дифференцировку гетероцист. [4] PatA формирует гетероцисты вдоль нитей и также важен для клеточного деления . PatS влияет на формирование паттерна гетероцист путем ингибирования дифференцировки гетероцист, когда группа дифференцирующихся клеток объединяется, чтобы сформировать прогетероцисты (незрелые гетероцисты). [5] Поддержание гетероцист зависит от фермента hetN. Образование гетероцист ингибируется присутствием фиксированного источника азота , такого как аммоний или нитрат . [6]
Формирование гетероцист
[ редактировать ]При образовании гетероцист из вегетативной клетки происходят следующие последовательности:
- Клетка увеличивается.
- Зернистые включения уменьшаются.
- Фотосинтетическая пластинка переориентируется.
- Стена наконец становится трехслойной. Эти три слоя развиваются за пределами внешнего слоя клетки.
- Средний слой однородный.
- Внутренний слой ламинирован.
- Стареющая гетероциста подвергается вакуолизации и, наконец, отрывается от нити, вызывая фрагментацию. Эти фрагменты называются гормогониями (единственное число гормогоний ) и подвергаются бесполому размножению.
Цианобактерии, образующие гетероцисты, делятся на отряды Nostocales и Stigonematales , которые образуют простые и ветвящиеся нити соответственно. Вместе они образуют монофилетическую группу с очень низкой генетической изменчивостью .
Симбиотические отношения
[ редактировать ]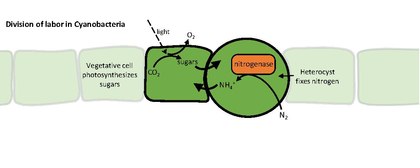
Бактерии также могут вступать в симбиотические отношения с некоторыми растениями. В таких отношениях бактерии реагируют не на наличие азота, а на сигналы, вырабатываемые растением для дифференцировки гетероцист. До 60% клеток могут стать гетероцистами, обеспечивая растение фиксированным азотом в обмен на фиксированный углерод. [6] Сигнал, вырабатываемый растением, и стадия дифференцировки гетероцист, на которую он влияет, неизвестны. По-видимому, симбиотический сигнал, генерируемый растением, действует до активации NtcA, поскольку hetR необходим для симбиотической дифференцировки гетероцист. Для симбиотической ассоциации с растением необходим ntcA, поскольку бактерии с мутировавшим ntcA не могут заражать растения. [7]
Анабаэна-Азолла
[ редактировать ]Примечательными симбиотическими отношениями являются Anabaena цианобактерии с Azolla растениями . Анабаена обитает на стеблях и листьях растений Азоллы . [8] Растение азолла он подвергается фотосинтезу и обеспечивает фиксированным углеродом , который анабаену может использовать в качестве источника энергии для динитрогеназ в клетках гетероцист. [8] В свою очередь, гетероцисты способны снабжать вегетативные клетки и растение Азолла фиксированным азотом в форме аммиака , который поддерживает рост обоих организмов. [8] [9]
Эти симбиотические отношения используются людьми в сельском хозяйстве. В Азии Azolla растения , содержащие виды Anabaena, используются в качестве биоудобрения там, где азот ограничен. [8] а также в кормах для животных . [9] Различные штаммы Azolla-Anabaena подходят для разных условий и могут привести к различиям в урожайности сельскохозяйственных культур. [10] рисовые культуры, выращенные с использованием Azolla-Anabaena в качестве биоудобрения, дают гораздо большее количество и качество продукции по сравнению с культурами без цианобактерий. Было показано, что [9] [11] Растения Azolla-Anabaena выращивают до и после посадки риса. [9] По мере роста растения Azolla-Anabaena накапливают фиксированный азот за счет действия ферментов нитрогеназы и органический углерод в результате фотосинтеза растений Azolla и вегетативных клеток Anabaena . [9] Когда растения Azolla-Anabaena большое количество связанного азота, фосфора , органического углерода и многих других питательных веществ, создавая богатую среду, идеальную для роста рисовых культур. умирают и разлагаются, они выделяют в почву [9]
Взаимоотношения Анабаены из окружающей среды - и Азоллы также изучались как возможный метод удаления загрязнителей процесс, известный как фиторемедиация . [12] Анабаена сп. вместе с Azolla caroliniana Было показано, что успешно удаляет уран , токсичный загрязнитель, вызванный добычей полезных ископаемых , а также тяжелые металлы ртуть (II) , хром (III) и хром (VI) из загрязненных сточных вод. [12] [13]
-
 Азолла Каролиниана Растение
Азолла Каролиниана Растение -
 Anabaena circinalis Нить
Anabaena circinalis Нить -
 цилиндросперма Нить
цилиндросперма Нить



Ссылки
[ редактировать ]- ^ Фундаментальная биология (18 марта 2016 г.). «Бактерии» .
- ^ Волк, КП; Эрнст, А.; Эльхай, Дж. (1994). «Метаболизм и развитие гетероцист». Молекулярная биология цианобактерий . стр. 100-1 769–823. дои : 10.1007/978-94-011-0227-8_27 . ISBN 978-0-7923-3273-2 .
- ^ Эрреро, Антония; Муро-Пастор, Алисия М.; Флорес, Энрике (15 января 2001 г.). «Контроль азота в цианобактериях» . Журнал бактериологии . 183 (2): 411–425. дои : 10.1128/JB.183.2.411-425.2001 . ISSN 0021-9193 . ПМК 94895 . ПМИД 11133933 .
- ^ Хига, Келли С.; Каллахан, Шон М. (1 августа 2010 г.). «Эктопическая экспрессия hetP может частично обойти необходимость в hetR при дифференцировке гетероцист штаммом Anabaena sp. PCC 7120». Молекулярная микробиология . 77 (3): 562–574. дои : 10.1111/j.1365-2958.2010.07257.x . ISSN 1365-2958 . ПМИД 20545862 .
- ^ Ороско, Кристина К.; Риссер, Дуглас Д.; Каллахан, Шон М. (2006). «Анализ эпистаза четырех генов штамма Anabaena sp. PCC 7120 предполагает связь между PatA и PatS в формировании структуры гетероцист» . Журнал бактериологии . 188 (5): 1808–1816. дои : 10.1128/JB.188.5.1808-1816.2006 . ISSN 0021-9193 . ПМЦ 1426565 . ПМИД 16484191 .
- ^ Перейти обратно: а б Ли, Роберт Эдвард. Психология (PDF) . Проверено 9 октября 2017 г.
- ^ Микс, Дж.К.; Эльхай, Дж (2002). «Регуляция клеточной дифференцировки нитчатых цианобактерий в свободноживущих и связанных с растениями состояниях симбиотического роста» . Обзоры микробиологии и молекулярной биологии . 66 (1): 94–121, оглавление. дои : 10.1128/ММБР.66.1.94-121.2002 . ПМК 120779 . ПМИД 11875129 .
- ^ Перейти обратно: а б с д ван Хов, К.; Лежен, А. (2002). «Азолла: симбиоз анабаены». Биология и окружающая среда: Труды Ирландской королевской академии . 102Б (1): 23–26. дои : 10.1353/bae.2002.0036 . JSTOR 20500136 . S2CID 245843704 .
- ^ Перейти обратно: а б с д и ж Вайшампаян, А.; Синха, Р.П.; Хедер, Д.-П.; Дей, Т.; Гупта, А.К.; Бхан, У.; Рао, AL (2001). «Цианобактериальные биоудобрения в рисоводстве». Ботанический обзор . 67 (4): 453–516. дои : 10.1007/bf02857893 . JSTOR 4354403 . S2CID 20058464 .
- ^ Бокки, Стефано; Мальджольо, Антонино (2010). «Азолла-Анабаена как биоудобрение для рисовых полей в долине По, рисовой зоне умеренного пояса в Северной Италии» (PDF) . Международный журнал агрономии . 2010 : 1–5. дои : 10.1155/2010/152158 . ISSN 1687-8159 .
- ^ Сингх, С.; Прасад, Р.; Сингх, Б.В.; Гоял, СК; Шарма, С.Н. (1 июня 1990 г.). «Влияние зеленых удобрений, сине-зеленых водорослей и мочевины, покрытой нимом, на водно-болотный рис (Oryza sativa L.)». Биология и плодородие почв . 9 (3): 235–238. дои : 10.1007/bf00336232 . ISSN 0178-2762 . S2CID 38989291 .
- ^ Перейти обратно: а б Бенничелли, Р.; Стемпневска, З.; Банах, А.; Шайноча, К.; Островский, Дж. (1 апреля 2004 г.). «Способность Azolla caroliniana удалять тяжелые металлы (Hg(II), Cr(III), Cr(VI)) из городских сточных вод». Хемосфера . 55 (1): 141–146. Бибкод : 2004Chmsp..55..141B . doi : 10.1016/j.chemSphere.2003.11.015 . ПМИД 14720557 .
- ^ Пан, Чанчунь; Ху, Нан; Дин, Дексин; Ху, Цзиньсун; Ли, Гуанъюэ; Ван, Юндон (01 января 2016 г.). «Экспериментальное исследование синергетического эффекта между Азоллой и Анабаеной при удалении урана из растворов симбиотической системой Азолла-анабаена». Журнал радиоаналитической и ядерной химии . 307 (1): 385–394. дои : 10.1007/s10967-015-4161-y . ISSN 0236-5731 . S2CID 82545272 .