Цианобактериальная морфология

Морфология цианобактерий относится к форме или форме цианобактерий. Цианобактерии — это большой и разнообразный тип бактерий, определяемый уникальной комбинацией пигментов и способностью осуществлять кислородный фотосинтез . [ 2 ] [ 3 ]
Цианобактерии часто живут колониальными скоплениями , которые могут принимать множество форм. [ 3 ] Особый интерес среди многих видов цианобактерий представляют те, которые живут колониально в удлиненных волоскообразных структурах, известных как трихомы . Эти нитчатые виды могут содержать от сотен до тысяч клеток. [ 3 ] Они часто доминируют в верхних слоях микробных матов, встречающихся в экстремальных условиях, таких как горячие источники , гиперсоленая вода , пустыни и полярные регионы. [ 4 ] а также широко распространены в более обыденной среде. [ 3 ]
Многие нитчатые виды также подвижны , скользят вдоль своей длинной оси и демонстрируют фотодвижение , с помощью которого трихома модулирует свое скольжение в зависимости от падающего света. Было обнаружено, что последний играет важную роль в обеспечении трихом оптимальных условий освещения, которые могут либо подавлять клетки, если падающий свет слишком слабый, либо повреждать клетки, если слишком сильный. [ 3 ]
Обзор
[ редактировать ]Для эффективной работы клеточных функций требуется хорошо организованная и скоординированная внутренняя структура. Клеткам необходимо создавать, поддерживать, а иногда и изменять свою форму, что позволяет им быстро менять свое поведение в ответ на внешние факторы. На разных стадиях жизненного цикла, таких как рост клеток , деление клеток или дифференцировка клеток , внутренние структуры должны динамически адаптироваться к текущим требованиям. У эукариот эти разнообразные задачи выполняются цитоскелетом : белковыми полимерами , которые собираются в стабильные или динамические нити или канальцы in vivo и in vitro . Эукариотический цитоскелет исторически делится на три класса: актиновые филаменты (состоящие из актина мономеров ), микротрубочки (состоящие из субъединиц тубулина ) и промежуточные филаменты (ПФ), хотя в последние годы были идентифицированы и другие классы цитоскелета. [ 5 ] [ 6 ] Только совместная работа всех трех цитоскелетных систем обеспечивает правильную клеточную механику. [ 7 ] [ 8 ]
Длительная догма о том, что прокариоты, в силу своей простой формы клеток, не нуждаются в элементах цитоскелета, была окончательно опровергнута открытием FtsZ прокариотического тубулина , гомолога , [ 9 ] [ 10 ] [ 11 ] и MreB , гомолог бактериального актина. [ 12 ] [ 13 ] Эти открытия положили начало интенсивным поискам других белков цитоскелета у бактерий и архей, что в конечном итоге привело к идентификации бактериальных IF-подобных белков, таких как Crescentin из Caulobacter crescentus. [ 14 ] и даже классы бактериально-специфичных цитоскелетных белков, включая бактофилины . [ 15 ] Постоянный приток новых данных, наконец, установил, что многочисленные прокариотические клеточные функции, включая деление клеток, удлинение клеток или сегрегацию бактериальных микрокомпартментов, регулируются прокариотическим цитоскелетом. [ 16 ] [ 17 ] [ 8 ]
Цианобактерии — единственные известные на сегодняшний день прокариоты, способные осуществлять кислородный фотосинтез . На основании наличия внешней мембраны цианобактерии обычно относят к грамотрицательным бактериям . Однако, в отличие от других грамотрицательных бактерий, цианобактерии содержат необычно толстый слой пептидогликана (PG) между внутренней и внешней мембраной, таким образом, сохраняя черты обоих грамфенотипов. [ 18 ] [ 19 ] [ 20 ] Кроме того, степень сшивки PG у цианобактерий намного выше, чем у других грамотрицательных бактерий, хотя тейхоевые кислоты, обычно присутствующие у грамположительных бактерий, отсутствуют. [ 21 ] [ 8 ]
Хотя цианобактерии монофилетичны , [ 22 ] их клеточная морфология чрезвычайно разнообразна и варьируется от одноклеточных видов до сложных многоклеточных видов, дифференцирующих клетки. На основании этого наблюдения цианобактерии классически были разделены на пять подразделов. [ 23 ] Цианобактерии подраздела I ( Chroococcales ) одноклеточные и делятся путем бинарного деления или почкования, тогда как цианобактерии подраздела II ( Pleurocapsales ) также являются одноклеточными, но могут подвергаться множественным делениям, давая начало множеству мелких дочерних клеток, называемых баеоцитами . Подраздел III включает многоклеточные, недифференцирующие клетки цианобактерии ( Oscillatoriales ), а подразделы IV и V цианобактерии ( Nostocales и Stigonematales ) — многоклеточные, клеточно-дифференцирующие цианобактерии, образующие специализированные типы клеток в отсутствие связанного азота ( гетероцисты ), при неблагоприятных условиях ( акинеты) . ) или для распространения и инициирования симбиоза ( гормогония ). В то время как подразделы III и IV образуют линейные клеточные нити (называемые трихомами ), окруженные общей оболочкой, подраздел V может давать боковые ветви и/или делиться в нескольких плоскостях, образуя многорядные трихомы. [ 23 ] Учитывая эту сложную морфологию, было предположено, что определенные специфичные для подраздела V (цитоскелетные) белки могут быть ответственны за этот фенотип. Однако не было идентифицировано ни одного конкретного гена, распределение которого специфически коррелировало бы с морфологией клеток среди различных подсекций цианобактерий. [ 24 ] [ 25 ] Таким образом, представляется более вероятным, что за морфологическое разнообразие цианобактерий ответственна дифференциальная экспрессия генов роста и деления клеток, а не наличие или отсутствие одного гена. [ 24 ] [ 26 ] [ 8 ]
Морфогенез
[ редактировать ]
Морфогенез – это биологический процесс, который заставляет организм приобретать свою форму. Цианобактерии демонстрируют высокую степень морфологического разнообразия и могут подвергаться множеству процессов клеточной дифференцировки, чтобы адаптироваться к определенным условиям окружающей среды. Это помогает им процветать практически во всех средах обитания на Земле, от пресноводных до морских и наземных, включая даже симбиотические взаимодействия. [ 27 ] [ 8 ]
Одним из факторов, который может вызывать морфологические изменения цианобактерий, является свет. Поскольку цианобактерии — это бактерии, которые используют свет в качестве топлива для своих фотосинтетических механизмов, производящих энергию, они зависят от восприятия света, чтобы оптимизировать свою реакцию и избегать вредного света, который может привести к образованию активных форм кислорода и впоследствии к их гибели. [ 28 ] Оптимальные условия освещения можно определить по количеству ( освещенности ), продолжительности ( цикл день-ночь ) и длине волны ( цвет света ). Пригодный для фотосинтеза световой диапазон солнечного спектра обычно называют ФАР ( фотосинтетически активное излучение ), но некоторые цианобактерии могут расширяться за счет ФАР, поглощая не только видимый спектр, но и ближний инфракрасный световой спектр. При этом используются различные хлорофиллы и обеспечивается фототрофный рост до длины волны 750 нм. [ 29 ] Чтобы воспринимать свет в этом диапазоне длин волн, цианобактерии обладают различными суперсемейства фитохромов фоторецепторами . [ 30 ] [ 8 ]
Морфологическая пластичность , или способность одной клетки менять разные формы, является общей стратегией многих бактерий в ответ на изменения окружающей среды или как часть их нормального жизненного цикла. [ 31 ] [ 32 ] [ 33 ] Бактерии могут изменять свою форму путем более простых переходов от палочек к кокковидным (и наоборот), как у Escherichia coli . [ 34 ] путем более сложных переходов при установлении многоклеточности [ 31 ] или путем развития специализированных клеток, структур или придатков, когда популяция ведет плеоморфный образ жизни . [ 35 ] Точные молекулярные цепи, которые управляют этими морфологическими изменениями, еще предстоит определить, однако постоянным фактором является то, что форма клеток определяется жестким PG-саккулюсом, который состоит из нитей гликанов, сшитых пептидами. Чтобы расти, клетки должны синтезировать новый PG, разрушая существующий полимер, чтобы вставить вновь синтезированный материал. То, как клетки растут и удлиняются, тщательно изучалось на модельных организмах как палочковидных, так и палочковидных. [ 36 ] [ 37 ] и коккоидные бактерии. [ 38 ] Однако молекулярная основа морфологической пластичности и плеоморфизма у более сложных бактерий также медленно выясняется. [ 33 ] [ 8 ]
Несмотря на свою морфологическую сложность, цианобактерии содержат все консервативные и известные к настоящему времени бактериальные морфогены . [ 8 ] Понимание морфогенеза цианобактерий является сложной задачей, поскольку среди таксонов цианобактерий существует множество морфотипов , которые также могут варьироваться в пределах данного штамма в течение его жизненного цикла. [ 23 ] Изменения в морфологии клеток или даже трихом представляют собой задачи, требующие активного ремоделирования клеточной стенки, и до сих пор у цианобактерий не было идентифицировано генов, относящихся к различным морфотипам. [ 24 ] Следовательно, наиболее вероятный сценарий заключается в том, что гены или их продукты по-разному регулируются во время этих морфологических изменений клеток. [ 26 ] как это предполагалось для большинства бактерий. [ 33 ] У многоклеточных цианобактерий разделение труда между клетками внутри трихомы достигается за счет различных стратегий клеточного программирования. Таким образом, регуляция генов происходит по-разному в этих конкретных типах клеток [30,97,98]. [ 8 ]
Разнообразие форм
[ редактировать ]Цианобактерии демонстрируют поразительную морфологическую изменчивость: от одноклеточных и колониальных до многоклеточных нитчатых форм. Размер их клеток варьирует от менее 1 мкм в диаметре ( пикоцианобактерии ) до 100 мкм (некоторые тропические формы рода Oscillatoria ). [ 39 ] [ 40 ] [ 41 ]
Нитчатые формы демонстрируют функциональную дифференцировку клеток, такую как гетероцисты (для фиксации азота), акинеты (клетки на стадии покоя) и гормогонии (репродуктивные, подвижные нити). Вместе с имеющимися у них межклеточными связями они считаются первыми признаками многоклеточности. [ 31 ] [ 42 ] [ 43 ] [ 44 ]
Многие цианобактерии образуют подвижные клеточные нити, называемые гормогониями , которые удаляются от основной биомассы, чтобы отпочковаться и сформировать новые колонии в другом месте. [ 45 ] [ 46 ] Клетки гормогония часто тоньше, чем в вегетативном состоянии, а клетки на обоих концах подвижной цепи могут быть суженными. Чтобы отделиться от родительской колонии, гормогонию часто приходится разрывать более слабую клетку нити, называемую некридием.

масштабные линейки около 10 мкм



Schizothrix Calcicola , Gloeocapsa , Coccofloris , Microcoleus vaginatus и Rivularia.
Колониальные и одноклеточные
[ редактировать ]В водной среде обитания одноклеточные цианобактерии считаются важной группой с точки зрения численности, разнообразия и экологического характера. [ 47 ] Одноклеточные цианобактерии имеют сферические, яйцевидные или цилиндрические клетки, которые могут объединяться в колонии неправильной или правильной формы, связанные вместе слизистым матриксом ( слизью ), выделяемым во время роста колонии. [ 48 ] В зависимости от вида количество клеток в каждой колонии может варьироваться от двух до нескольких тысяч. [ 47 ] [ 1 ]
Каждая отдельная клетка (каждая отдельная цианобактерия) обычно имеет толстую студенистую клеточную стенку . [ 49 ] У них отсутствуют жгутики , но гормогонии некоторых видов могут передвигаться, скользя по поверхности. [ 50 ]
-
 Мериспедия образует прямоугольные колонии, скрепленные слизистой матрицей . Виды этого рода делятся только в двух направлениях, создавая характерный сетчатый узор, расположенный рядами и плоскостями. [ 51 ]
Мериспедия образует прямоугольные колонии, скрепленные слизистой матрицей . Виды этого рода делятся только в двух направлениях, создавая характерный сетчатый узор, расположенный рядами и плоскостями. [ 51 ]

-
 Колонии Nostoc pruniforme «желейные шарики».
Колонии Nostoc pruniforme «желейные шарики». -
 Колониальные цианобактерии Stratonostoc
Колониальные цианобактерии Stratonostoc -
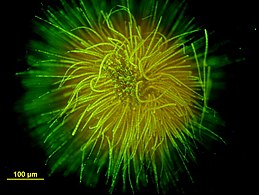 Шаровидная колония Gloeotrichia echinulata.
Шаровидная колония Gloeotrichia echinulata. -
 Цианобактериальная колония Lyngbya majuscula
Цианобактериальная колония Lyngbya majuscula




Нитчатые и многоклеточные
[ редактировать ]-
![Пример структуры нитчатых цианобактерий (Oscillatoria lutea) с сетчатым узором [52]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b9/Filamentous_cyanobacteria_structure_of_Oscillatoria_lutea.jpg/359px-Filamentous_cyanobacteria_structure_of_Oscillatoria_lutea.jpg) Пример структуры нитчатых цианобактерий ( Oscillatoria lutea ) с сетчатым узором. [ 52 ]
Пример структуры нитчатых цианобактерий ( Oscillatoria lutea ) с сетчатым узором. [ 52 ]
![Пример структуры нитчатых цианобактерий (Oscillatoria lutea) с сетчатым узором [52]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b9/Filamentous_cyanobacteria_structure_of_Oscillatoria_lutea.jpg/359px-Filamentous_cyanobacteria_structure_of_Oscillatoria_lutea.jpg)
Некоторые нитчатые виды могут дифференцироваться в несколько различных клеток типов :
- вегетативные клетки – нормальные фотосинтезирующие клетки, образующиеся при благоприятных условиях роста.
- акинеты - устойчивые к климату споры, которые могут образовываться, когда условия окружающей среды становятся суровыми.
- толстостенные гетероцисты , содержащие фермент нитрогеназу, необходимый для фиксации азота. [ 53 ] [ 54 ] [ 55 ] в анаэробной среде из-за чувствительности к кислороду. [ 55 ]
Многие многоклеточные нитчатые формы Oscillatoria способны к волнообразным движениям; нить колеблется взад и вперед. В толщах воды некоторые цианобактерии плавают, образуя газовые пузырьки , как у архей . [ 56 ] Эти везикулы не являются органеллами как таковыми. Они ограничены не липидной мембраной , а белковой оболочкой.

A–C: Microcoleus steenstrupii D–E: Tolypothrix Desertorum F: Scytonema cf. Calcicola G: S. ср. Calcicola H: S. ср. с алкоголем
Масштабная линейка = 10 мкм
-
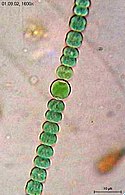
-

-
 Спиральные нити цианобактерий
Спиральные нити цианобактерий -
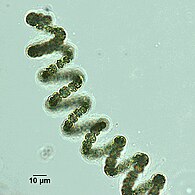 Спиральная нить долихоспермума
Спиральная нить долихоспермума -
 Виды Lyngbya образуют длинные неразветвленные нити внутри жестких слизистых оболочек, которые могут образовывать клубки или маты, смешиваясь с другими фитопланктона. видами
Виды Lyngbya образуют длинные неразветвленные нити внутри жестких слизистых оболочек, которые могут образовывать клубки или маты, смешиваясь с другими фитопланктона. видами





Разветвленный
[ редактировать ]-

-
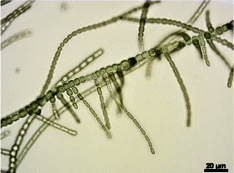 Истинный фенотип ветвления Fischerella Thermalis. колонии
Истинный фенотип ветвления Fischerella Thermalis. колонии


Гетероцисты
[ редактировать ]Гетероцисты — специализированные азотфиксирующие клетки, образующиеся при азотном голодании некоторыми нитчатыми цианобактериями, такими как Nostoc punctiforme , Cylindrospermum stagnale и Anabaena sphaerica . [ 58 ] Они фиксируют азот из атмосферного N 2 с помощью фермента нитрогеназы , чтобы обеспечить клетки нити азотом для биосинтеза. [ 59 ]
Движение
[ редактировать ]
Цианобактерии распространены повсеместно, находя среду обитания в большинстве водоемов и в экстремальных условиях, таких как полярные регионы, пустыни, соленые озера и горячие источники. [ 60 ] [ 61 ] [ 62 ] У них также развилось удивительно сложное коллективное поведение, лежащее на границе между одноклеточной и многоклеточной жизнью. Например, нитчатые цианобактерии живут в длинных цепочках клеток, которые объединяются в более крупные структуры, включая биопленки , биоматы и строматолиты . [ 63 ] [ 64 ] Эти большие колонии обеспечивают жесткую, стабильную и долговременную среду для сообществ бактерий. Кроме того, биопленки на основе цианобактерий можно использовать в качестве биореакторов для производства широкого спектра химических веществ, включая биотопливо, такое как биодизель и этанол. [ 65 ] Однако, несмотря на их важность для истории жизни на Земле, а также их коммерческий и экологический потенциал, остаются основные вопросы о том, как нитчатые цианобактерии перемещаются, реагируют на окружающую среду и самоорганизуются в коллективные закономерности и структуры. [ 52 ]
У всех известных цианобактерий отсутствуют жгутики ; [ 66 ] однако многие нитчатые виды передвигаются по поверхности путем скольжения - формы передвижения, при которой не наблюдается никаких физических придатков, помогающих движению. [ 67 ] Фактический механизм планирования до конца не изучен, хотя с момента его открытия прошло более ста лет. [ 68 ] [ 69 ] Одна теория предполагает, что скользящее движение у цианобактерий опосредовано непрерывной секрецией полисахаридов через поры отдельных клеток. [ 70 ] [ 71 ] [ 72 ] Другая теория предполагает, что скользящее движение включает использование пилей IV типа , полимерных ансамблей белка пилина . [ 73 ] как движущие двигатели движения. [ 74 ] [ 75 ] [ 76 ] Однако неясно, как действие этих пилей приведет к движению, и некоторые предполагают, что они втягиваются. [ 77 ] в то время как другие предлагают нажать, [ 76 ] генерировать силы. Другие ученые предположили, что поверхностные волны, генерируемые сокращением слоя фибрилл, являются механизмом скользящего движения у Oscillatoria . [ 78 ] [ 79 ] Недавние исследования также предполагают, что в скользящем движении могут быть задействованы колебания формы и капиллярные силы. [ 80 ] [ 52 ]
Благодаря коллективному взаимодействию нитчатые цианобактерии самоорганизуются в колонии или биопленки — симбиотические сообщества, встречающиеся в самых разных экологических нишах. Их более крупномасштабные коллективные структуры характеризуются разнообразными формами, включая пучки, вихри и сетчатые узоры. [ 81 ] [ 82 ] Подобные закономерности наблюдались в записях окаменелостей. [ 83 ] [ 82 ] [ 84 ] Известно, что у нитчатых цианобактерий механика нитей способствует самоорганизации, например, при определении того, как одна нить будет изгибаться при контакте с другими нитями или препятствиями. [ 85 ] Кроме того, биопленки и биоматы демонстрируют некоторые удивительно консервативные макромеханические свойства, обычно ведут себя как вязкоупругие материалы со временем релаксации около 20 минут. [ 86 ] [ 52 ]
Цианобактерии предъявляют строгие требования к свету. Слишком мало света может привести к недостаточному производству энергии, а у некоторых видов может привести к тому, что клетки прибегнут к гетеротрофному дыханию. [ 4 ] Слишком много света может подавлять клетки, снижать эффективность фотосинтеза и вызывать повреждение в результате обесцвечивания. УФ-излучение особенно смертельно опасно для цианобактерий, при этом нормальный уровень солнечной радиации в некоторых случаях оказывается для этих микроорганизмов значительно вредным. [ 87 ] [ 88 ] [ 3 ]
Нитчатые цианобактерии, живущие в микробных матах, часто мигрируют внутри мата вертикально и горизонтально, чтобы найти оптимальную нишу, которая уравновешивает их потребности в свете для фотосинтеза и их чувствительность к фотоповреждениям. Например, нитчатые цианобактерии Oscillatoria sp. и Spirulina subsalsa, обнаруженные в гиперсоленых донных матах Герреро-Негро, Мексика, в течение дня мигрируют вниз, в нижние слои, чтобы избежать интенсивного солнечного света, а затем поднимаются на поверхность в сумерках. [ 89 ] Напротив, популяция Microcoleus chthonoplastes, обнаруженная в гиперсоленых матах в Сален-де-Жиро , Камарг, Франция, мигрирует в верхний слой мата в течение дня и равномерно распределяется по мату ночью. [ 90 ] Эксперимент in vitro с использованием P. uncinatum также продемонстрировал склонность этого вида мигрировать, чтобы избежать вредного воздействия радиации. [ 87 ] [ 88 ] Эти миграции обычно являются результатом какого-то фотодвижения, хотя свою роль могут сыграть и другие формы такси. [ 91 ] [ 3 ]
Многие виды цианобактерий способны планировать. Скольжение — это форма движения клеток, которая отличается от ползания или плавания тем, что не зависит от какого-либо очевидного внешнего органа или изменения формы клетки и происходит только в присутствии субстрата . [ 92 ] [ 93 ] Скольжение нитчатых цианобактерий, по-видимому, осуществляется за счет механизма «струи слизи», при котором клетки выдавливают гель, который быстро расширяется по мере гидратации, создавая движущую силу. [ 94 ] [ 95 ] хотя некоторые одноклеточные цианобактерии используют пили IV типа для скольжения. [ 96 ] Отдельные клетки трихомы имеют два набора пор для выделения слизи. Каждый набор организован в кольцо у клеточных перегородок и выбрасывает слизь под острым углом. [ 97 ] Наборы выдавливают слизь в противоположных направлениях, поэтому во время планирования может активироваться только один набор. Альтернативная гипотеза состоит в том, что клетки используют сократительные элементы, которые создают волны, пробегающие по поверхности внутри трубки со слизью, как у дождевого червя. [ 98 ] Трихомы вращаются по спирали, угол которых соответствует Угол наклона сократительных трихом Кастенгольца. [ 3 ]


Кажется, что клетки координируют направление своего скольжения с помощью электрического потенциала, который устанавливает полярность в трихомах и, таким образом, образует «голову» и «хвост». [ 99 ] Трихомы обычно меняют свою полярность случайным образом со средним периодом от нескольких минут до часов. [ 100 ] [ 101 ] Многие виды также образуют полужесткую оболочку, которая остается в виде полой трубки по мере продвижения трихомы вперед. Когда трихома меняет направление, она может вернуться в оболочку или вырваться наружу. [ 102 ] [ 3 ]
Oscillatoria — род нитчатых цианобактерий, названный в честь колебаний в их движении. Нити в колониях скользят вперед и назад друг относительно друга, пока вся масса не переориентируется на источник света. Осциллатории преимущественно сине-зеленого или коричнево-зеленого цвета и обычно встречаются в поилках. Он размножается путем фрагментации, образуя длинные нити клеток, которые могут распадаться на фрагменты, называемые гормогониями . Затем гормогонии могут вырасти в новые, более длинные нити.
Эксперимент Хэдера с цианографом
[ редактировать ]
В 1987 году Хедер продемонстрировал, что трихомы могут довольно точно позиционировать себя в окружающей среде посредством фотодвижения . В эксперименте Хедера с цианографом фотографический негатив проецируется на чашку Петри, содержащую культуру Phormidium uncinatum . [ 103 ] [ 104 ] Через несколько часов трихомы перемещаются из более темных участков в более светлые, образуя фотографический позитив на культуре. Эксперимент показывает, что фотодвижение эффективно не только для дискретных световых ловушек, но и для мелкоструктурированных, непрерывно дифференцированных световых полей. [ 3 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Мехдизаде Аллаф, Малихе; Перхоссаини, Хасан (24 марта 2022 г.). «Цианобактерии: модельные микроорганизмы и не только» . Микроорганизмы . 10 (4). MDPI AG: 696. doi : 10.3390/microorganisms10040696 . ISSN 2076-2607 . ПМК 9025173 . ПМИД 35456747 .
 Измененный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Измененный материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Уиттон, Брайан А.; Поттс, Малькольм (2012). «Знакомство с цианобактериями». Экология цианобактерий II . стр. 1–13. дои : 10.1007/978-94-007-3855-3_1 . ISBN 978-94-007-3854-6 .
- ^ Перейти обратно: а б с д и ж г час я дж к л Тамулонис, Карлос; Постма, Мартен; Каандорп, Яап (2011). «Моделирование нитчатых цианобактерий раскрывает преимущества длинных и быстрых трихом для оптимизации светового воздействия» . ПЛОС ОДИН . 6 (7): e22084. Бибкод : 2011PLoSO...622084T . дои : 10.1371/journal.pone.0022084 . ПМЦ 3138769 . ПМИД 21789215 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Перейти обратно: а б Сталь, Лукас Дж. (5 июля 2012 г.). «Цианобактериальные маты и строматолиты» . В Уиттоне, Брайан А. (ред.). Экология цианобактерий II . Спрингер. стр. 65–126. ISBN 9789400738553 .
- ^ Мозли, Джеймс Б. (2013). «Расширенный взгляд на цитоскелет эукариот» . Молекулярная биология клетки . 24 (11). Американское общество клеточной биологии (ASCB): 1615–1618. дои : 10.1091/mbc.e12-10-0732 . ISSN 1059-1524 . ПМЦ 3667716 . ПМИД 23722945 .
- ^ Альбертс, Брюс; Хилд, Ребекка; Джонсон, Александр; Морган, Дэвид Оуэн; Рафф, Мартин С.; Робертс, К.; Уолтер, Питер (2022). Молекулярная биология клетки . Нью-Йорк, штат Нью-Йорк. ISBN 978-0-393-88482-1 . OCLC 1276902141 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Хубер, Флориан; Буаре, Аделина; Лопес, Магдалена Пресиадо; Кендеринк, Гейсье Х (2015). «Цитоскелетные перекрестные помехи: когда объединяются три разные личности». Современное мнение в области клеточной биологии . 32 . Эльзевир Б.В.: 39–47. дои : 10.1016/j.ceb.2014.10.005 . ISSN 0955-0674 . ПМИД 25460780 . S2CID 40360166 .
- ^ Перейти обратно: а б с д и ж г час я дж Спрингштейн, Бенджамин Л.; Нюрнберг, Деннис Дж.; Вайс, Грегор Л.; Пилхофер, Мартин; Стукен, Карина (17 декабря 2020 г.). «Структурные детерминанты и их роль в морфогенезе цианобактерий» . Жизнь . 10 (12). MDPI AG: 355. Бибкод : 2020Life...10..355S . дои : 10.3390/life10120355 . ISSN 2075-1729 . ПМЦ 7766704 . ПМИД 33348886 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Би, Эрфей; Луткенхаус, Джо (1991). «Кольцевая структура FtsZ, связанная с делением Escherichia coli». Природа . 354 (6349). ООО «Спрингер Сайенс энд Бизнес Медиа»: 161–164. Бибкод : 1991Natur.354..161B . дои : 10.1038/354161a0 . ISSN 0028-0836 . ПМИД 1944597 . S2CID 4329947 .
- ^ де Бур, Пит; Кроссли, Робин; Ротфилд, Лоуренс (1992). «Основной белок деления бактериальных клеток FtsZ представляет собой ГТФазу». Природа . 359 (6392). ООО «Спрингер Сайенс энд Бизнес Медиа»: 254–256. Бибкод : 1992Natur.359..254D . дои : 10.1038/359254a0 . ISSN 0028-0836 . ПМИД 1528268 . S2CID 2748757 .
- ^ Лёве, Ян; Амос, Линда А. (1998). «Кристаллическая структура белка деления бактериальных клеток FtsZ». Природа . 391 (6663). ООО «Спрингер Сайенс энд Бизнес Медиа»: 203–206. Бибкод : 1998Natur.391..203L . дои : 10.1038/34472 . ISSN 0028-0836 . ПМИД 9428770 . S2CID 4330857 .
- ^ Борк, П; Сандер, К; Валенсия, А (15 августа 1992 г.). «АТФазный домен, общий для белков клеточного цикла прокариот, сахарных киназ, актина и белков теплового шока hsp70» . Труды Национальной академии наук . 89 (16): 7290–7294. Бибкод : 1992PNAS...89.7290B . дои : 10.1073/pnas.89.16.7290 . ISSN 0027-8424 . ПМК 49695 . ПМИД 1323828 .
- ^ ван ден Энт, Фусинита; Амос, Линда А.; Лёве, Ян (2001). «Прокариотическое происхождение актинового цитоскелета». Природа . 413 (6851). ООО «Спрингер Сайенс энд Бизнес Медиа»: 39–44. Бибкод : 2001Natur.413...39V . дои : 10.1038/35092500 . ISSN 0028-0836 . ПМИД 11544518 . S2CID 4427828 .
- ^ Аусмиес, Нора; Кун, Джеффри Р.; Джейкобс-Вагнер, Кристина (2003). «Бактериальный цитоскелет» . Клетка . 115 (6). Эльзевир Б.В.: 705–713. дои : 10.1016/s0092-8674(03)00935-8 . ISSN 0092-8674 . ПМИД 14675535 . S2CID 14459851 .
- ^ Кюн, Юлиана; Бригель, Ариана; Мёршель, Эрхард; Кант, Йорг; Лезер, Катя; Вик, Стефани; Дженсен, Грант Дж; Танбихлер, Мартин (3 декабря 2009 г.). «Бактофилины, вездесущий класс цитоскелетных белков, опосредующих полярную локализацию синтазы клеточной стенки у Caulobacter crescentus» . Журнал ЭМБО . 29 (2). Уайли: 327–339. дои : 10.1038/emboj.2009.358 . ISSN 0261-4189 . ПМЦ 2824468 . ПМИД 19959992 .
- ^ Лин, Лин; Танбихлер, Мартин (2013). «Нуклеотид-независимые цитоскелетные каркасы у бактерий». Цитоскелет . 70 (8). Уайли: 409–423. дои : 10.1002/см.21126 . ISSN 1949-3584 . ПМИД 23852773 . S2CID 40504066 .
- ^ Вагстафф, Джеймс; Лёве, Ян (22 января 2018 г.). «Прокариотические цитоскелеты: белковые нити, организующие мелкие клетки». Обзоры природы Микробиология . 16 (4). ООО «Спрингер Сайенс энд Бизнес Медиа»: 187–201. дои : 10.1038/nrmicro.2017.153 . ISSN 1740-1526 . ПМИД 29355854 . S2CID 3537215 .
- ^ Видео, Патрик; Риверс, Орион С.; Ушиджима, Блейк; Осиро, Рид Т.; Ким, Мин Джу; Филм, Бенджамин; Коузи, Лоралин М. (15 апреля 2016 г.). «Мутация генов murC и murB нарушает дифференцировку гетероцист у штамма Anabaena sp. PCC 7120» . Журнал бактериологии . 198 (8). Американское общество микробиологии: 1196–1206. дои : 10.1128/jb.01027-15 . ISSN 0021-9193 . ПМЦ 4859589 . ПМИД 26811320 .
- ^ Хойчик, Э; Баумайстер, В. (1995). «Структура оболочки четырех скользящих нитчатых цианобактерий» . Журнал бактериологии . 177 (9). Американское общество микробиологии: 2387–2395. дои : 10.1128/jb.177.9.2387-2395.1995 . ISSN 0021-9193 . ПМК 176896 . ПМИД 7730269 .
- ^ Гумбарт, Джеймс К.; Биби, Морган; Дженсен, Грант Дж.; Ру, Бенуа (20 февраля 2014 г.). «Структура и механика пептидогликана Escherichia coli, предсказанная моделированием в атомном масштабе» . PLOS Вычислительная биология . 10 (2). Публичная научная библиотека (PLoS): e1003475. Бибкод : 2014PLSCB..10E3475G . дои : 10.1371/journal.pcbi.1003475 . ISSN 1553-7358 . ПМЦ 3930494 . ПМИД 24586129 .
- ^ Хойчик, Эгберт; Гензель, Альфред (2000). «Стенки цианобактерий: новости из необычной прокариотической оболочки» . Журнал бактериологии . 182 (5). Американское общество микробиологии: 1191–1199. дои : 10.1128/jb.182.5.1191-1199.2000 . ISSN 0021-9193 . ПМК 94402 . ПМИД 10671437 .
- ^ Ширмейстер, Беттина Э; Антонелли, Александр; Багери, Хомаюн С (14 февраля 2011 г.). «Происхождение многоклеточности у цианобактерий» . Эволюционная биология BMC . 11 (1). Springer Science and Business Media LLC: 45. doi : 10.1186/1471-2148-11-45 . ISSN 1471-2148 . ПМЦ 3271361 . ПМИД 21320320 .
- ^ Перейти обратно: а б с Риппка, Розмари; Станье, Роджер Ю.; Дерюэль, Жозетт; Хердман, Майкл; Уотербери, Джон Б. (1 марта 1979 г.). «Родовые назначения, история штаммов и свойства чистых культур цианобактерий» . Микробиология . 111 (1). Общество микробиологов: 1–61. дои : 10.1099/00221287-111-1-1 . ISSN 1350-0872 .
- ^ Перейти обратно: а б с Даган, Таль; Реттгер, Мэйо; Стукен, Карина; Лэндан, Гидди; Кох, Робин; Майор, Питер; Гулд, Свен Б.; Горемыкин Вадим Викторович; Риппка, Розмари; Тандо де Марсак, Николь; Гуггер, Мюриэл; Локхарт, Питер Дж.; Аллен, Джон Ф.; Брюн, Ирис; Маус, Ирена; Пюлер, Альфред; Мартин, Уильям Ф. (7 декабря 2012 г.). «Геномы стигонематальных цианобактерий (подраздел V) и эволюция кислородного фотосинтеза от прокариот к пластидам» . Геномная биология и эволюция . 5 (1). Издательство Оксфордского университета (OUP): 31–44. дои : 10.1093/gbe/evs117 . ISSN 1759-6653 . ПМК 3595030 . ПМИД 23221676 .
- ^ Ши, Патрик М.; У, Дунъин; Латифи, Амель; Аксен, Сет Д.; Меньше, Дэвид П.; Талла, Эммануэль; Кальто, Александра; Цай, Фэй; Тандо де Марсак, Николь; Риппка, Розмари; Хердман, Майкл; Сивонен, Каарина; Курсен, Тереза; Лоран, Тьерри; Гудвин, Линн; Нолан, Мэтт; Давенпорт, Карен В.; Хан, Клифф С.; Рубин, Эдвард М.; Эйзен, Джонатан А.; Войке, Таня; Гуггер, Мюриэл; Керфельд, Шерил А. (31 декабря 2012 г.). «Улучшение охвата типа цианобактерий с помощью секвенирования генома, основанного на разнообразии» . Труды Национальной академии наук . 110 (3): 1053–1058. дои : 10.1073/pnas.1217107110 . ISSN 0027-8424 . ПМЦ 3549136 . ПМИД 23277585 .
- ^ Перейти обратно: а б Кох, Робин; Купчок, Анна; Стукен, Карина; Ильхан, Джудит; Хаммершмидт, Катрин; Даган, Таль (31 августа 2017 г.). «Пластичность прежде всего: молекулярные признаки сложной морфологической особенности нитчатых цианобактерий» . Эволюционная биология BMC . 17 (1). Springer Science and Business Media LLC: 209. doi : 10.1186/s12862-017-1053-5 . ISSN 1471-2148 . ПМК 5580265 . ПМИД 28859625 .
- ^ Гайсина, Лира А.; Сараф, Аникет; Сингх, Прашант (2019). «Цианобактерии в разнообразных средах обитания». Цианобактерии . Эльзевир. стр. 1–28. дои : 10.1016/b978-0-12-814667-5.00001-5 . ISBN 9780128146675 . S2CID 135429562 .
- ^ Поспишил, Павел (26 декабря 2016 г.). «Продуцирование активных форм кислорода Фотосистемой II как ответ на световой и температурный стресс» . Границы в науке о растениях . 7 . Frontiers Media SA: 1950. doi : 10.3389/fpls.2016.01950 . ISSN 1664-462X . ПМК 5183610 . ПМИД 28082998 .
- ^ Ларкум, полный привод; Ричи, Р.Дж.; Рэйвен, JA (2018). «Жизнь за счет Солнца: хлорофиллы, бактериохлорофиллы и родопсины». Фотосинтетика . 56 : 11–43. дои : 10.1007/s11099-018-0792-x . S2CID 4907693 .
- ^ Уилтбанк, Лиза Б.; Кехо, Дэвид М. (08 ноября 2018 г.). «Разнообразные световые реакции цианобактерий, опосредованные фоторецепторами суперсемейства фитохромов». Обзоры природы Микробиология . 17 (1). ООО «Спрингер Сайенс энд Бизнес Медиа»: 37–50. дои : 10.1038/s41579-018-0110-4 . ISSN 1740-1526 . ПМИД 30410070 . S2CID 256744429 .
- ^ Перейти обратно: а б с Классен, Деннис; Розен, Дэниел Э.; Койперс, Оскар П.; Сёгаард-Андерсен, Лотте; Ван Везель, Жиль П. (2014). «Бактериальные решения проблемы многоклеточности: рассказ о биопленках, нитях и плодовых телах» . Обзоры природы Микробиология . 12 (2): 115–124. дои : 10.1038/nrmicro3178 . hdl : 11370/0db66a9c-72ef-4e11-a75d-9d1e5827573d . ПМИД 24384602 . S2CID 20154495 .
- ^ Чжу, Цзайчунь, Минени, Ранга Б.; Цзэн, Чжэньчжун; Жозеп Г.; Ситч, Пьер; Арнет, Цао, Чуньсян; Лей, Эцуши, Чарльз Ли; Лиу, Юнвэнь; Мао, Яочжун; Пенуэлас, Хосеп, Бенджамин; Хуэй, Цзэн, Нин (25 апреля 2016 г. . Д.; Виви, Николас; Ван, Сяо, Ян , ) Изменение .6 8 ( . )
- ^ Перейти обратно: а б с Каккамо, Пол Д.; Брун, Ив В. (2018). «Молекулярные основы неканонической бактериальной морфологии» . Тенденции в микробиологии . 26 (3). Эльзевир Б.В.: 191–208. дои : 10.1016/j.tim.2017.09.012 . ISSN 0966-842X . ПМЦ 5834356 . ПМИД 29056293 .
- ^ Ланге, Р; Хенгге-Аронис, Р. (1991). «Регулируемая фазой роста экспрессия bolA и морфология клеток Escherichia coli в стационарной фазе контролируются новым сигма-фактором sigma S» . Журнал бактериологии . 173 (14). Американское общество микробиологии: 4474–4481. дои : 10.1128/jb.173.14.4474-4481.1991 . ISSN 0021-9193 . ПМК 208111 . ПМИД 1648559 .
- ^ Янг, Кевин Д. (2006). «Селективная ценность бактериальной формы» . Обзоры микробиологии и молекулярной биологии . 70 (3). Американское общество микробиологии: 660–703. дои : 10.1128/mmbr.00001-06 . ISSN 1092-2172 . ПМЦ 1594593 . ПМИД 16959965 .
- ^ Типас, Афанасий; Банжаф, Мануэль; Гросс, Кэрол А.; Фоллмер, Вальдемар (28 декабря 2011 г.). «От регуляции синтеза пептидогликана к росту и морфологии бактерий» . Обзоры природы Микробиология . 10 (2). ООО «Спрингер Сайенс энд Бизнес Медиа»: 123–136. дои : 10.1038/nrmicro2677 . ISSN 1740-1526 . ПМЦ 5433867 . ПМИД 22203377 .
- ^ Иган, Александр Дж. Ф.; Эррингтон, Джефф; Фоллмер, Вальдемар (18 мая 2020 г.). «Регуляция синтеза и ремоделирования пептидогликана». Обзоры природы Микробиология . 18 (8). Springer Science and Business Media LLC: 446–460. дои : 10.1038/s41579-020-0366-3 . ISSN 1740-1526 . ПМИД 32424210 . S2CID 256745837 .
- ^ Пиньо, Мариана Г.; Кьос, Мортен; Вининг, Ян-Виллем (16 августа 2013 г.). «Как обойти: механизмы, контролирующие рост и деление коккоидных бактерий» (PDF) . Обзоры природы Микробиология . 11 (9). ООО «Спрингер Сайенс энд Бизнес Медиа»: 601–614. дои : 10.1038/nrmicro3088 . ISSN 1740-1526 . ПМИД 23949602 . S2CID 205498610 .
- ^ Уиттон, Брайан А. (1992). «Разнообразие, экология и систематика цианобактерий». Фотосинтезирующие прокариоты . Бостон, Массачусетс: Springer US. стр. 1–51. дои : 10.1007/978-1-4757-1332-9_1 . ISBN 978-1-4757-1334-3 .
- ^ Шульц-Фогт, Хайде Н; Ангерт, Эстер Р; Гарсиа-Пишель, Ферран (28 сентября 2007 г.), «Гигантские бактерии», eLS , Wiley, doi : 10.1002/9780470015902.a0020371 , ISBN 9780470016176
- ^ Джассер, Ивона; Каллиери, Кристиана (11 февраля 2017 г.). «Пикоцианобактерии». Справочник по цианобактериальному мониторингу и анализу цианотоксинов . Чичестер, Великобритания: John Wiley & Sons, Ltd., стр. 19–27. дои : 10.1002/9781119068761.ch3 . ISBN 9781119068761 .
- ^ Нюрнберг, Деннис Дж.; Марискаль, Висенте; Паркер, Джейми; Мастроянни, Джулия; Флорес, Энрике; Муллино, Конрад В. (2014). «Ветвление и межклеточная коммуникация у цианобактерии Раздела V Mastigocladus laminosus, сложного многоклеточного прокариота». Молекулярная микробиология . 91 (5): 935–949. дои : 10.1111/mmi.12506 . hdl : 10261/99110 . ПМИД 24383541 . S2CID 25479970 .
- ^ Эрреро, Антония; Ставанс, Джоэл; Флорес, Энрике (2016). «Многоклеточная природа нитчатых гетероцистообразующих цианобактерий». Обзоры микробиологии FEMS . 40 (6): 831–854. дои : 10.1093/femsre/fuw029 . hdl : 10261/140753 . ПМИД 28204529 .
- ^ Орел, Аннабелла; Клеменич, ВМФ; Зарплата, Даниэла Дж.; Ржимский, Петр; Джаннуцци, Леда; Мартин, Мэри Виктория (2021). «Клеточная смерть цианобактерий: современное понимание и рекомендации для достижения консенсуса по ее номенклатуре » Границы микробиологии . 12 :631654 дои : 10.3389/fmicb.2021.631654 . ПМЦ 7965980 . ПМИД 33746925 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Риссер Д.Д., Чу В.Г., Микс Дж.К. (апрель 2014 г.). «Генетическая характеристика локуса hmp, кластера генов, подобного хемотаксису, который регулирует развитие и подвижность гормогоний у Nostoc punctiforme» . Молекулярная микробиология . 92 (2): 222–33. дои : 10.1111/mmi.12552 . ПМИД 24533832 . S2CID 37479716 .
- ^ Хайатан Б., Бэйнс Д.К., Ченг М.Х., Чо Ю.В., Хуинь Дж., Ким Р., Оморуи О.Х., Пантоджа А.П., Пак Дж.С., Пэн Дж.К., Сплитт С.Д., Тиан М.Ю., Риссер Д.Д. (май 2017 г.). «Предполагаемая O-связанная β-N-ацетилглюкозаминтрансфераза необходима для развития и подвижности гормонов у нитчатых цианобактерий Nostoc punctiforme» . Журнал бактериологии . 199 (9): e00075–17. дои : 10.1128/JB.00075-17 . ПМК 5388816 . ПМИД 28242721 .
- ^ Перейти обратно: а б Дворжак, Петр; Казаматта, Дейл А.; Хаслер, Петр; Ягодаржова, Ева; Норвич, Элисон Р.; Пуличкова, Алоизия (2017). «Разнообразие цианобактерий». Современные проблемы фототрофных прокариот . Чам: Международное издательство Springer. стр. 3–46. дои : 10.1007/978-3-319-46261-5_1 . ISBN 978-3-319-46259-2 .
- ^ Хор, Ингрид; Бартрам, Джейми (1999). Токсичные цианобактерии в воде: руководство по их последствиям для общественного здравоохранения, мониторингу и управлению . Лондон: E & FN Spon. ISBN 0-419-23930-8 . OCLC 40395794 .
- ^ Сингх. Учебник по ботанике «Разнообразие микробов и криптогам» . Публикации Растоги. ISBN 978-8171338894 .
- ^ «Различия между бактериями и цианобактериями» . Заметки по микробиологии . 29 октября 2015 г. Проверено 21 января 2018 г.
- ^ Палинска, Катажина А.; Лизак, Вернер; Риэль, Эрхард; Крумбейн, МЫ (17 октября 1996 г.). «Фенотипическая изменчивость идентичных генотипов: необходимость комбинированного подхода в таксономии цианобактерий, продемонстрированная на изолятах, подобных мерисопедиа». Архив микробиологии . 166 (4): 224–233. дои : 10.1007/s002030050378 . ПМИД 8824145 . S2CID 3022844 .
- ^ Перейти обратно: а б с д и Фалувеки, Миксон К.; Геринг, Лукас (2022). «Структурная механика нитчатых цианобактерий» . Журнал интерфейса Королевского общества . 19 (192). Королевское общество. дои : 10.1098/rsif.2022.0268 . ISSN 1742-5662 . ПМЦ 9326267 . ПМИД 35892203 . Измененный текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Микс Дж.К., Элхай Дж., Тиль Т., Поттс М., Лаример Ф., Ламердин Дж., Предки П., Атлас Р. (2001). «Обзор генома Nostoc punctiforme, многоклеточной симбиотической цианобактерии». Исследования фотосинтеза . 70 (1): 85–106. дои : 10.1023/А:1013840025518 . ПМИД 16228364 . S2CID 8752382 .
- ^ Golden JW, Юн Х.С. (декабрь 1998 г.). «Формирование гетероцист в Анабаене». Современное мнение в микробиологии . 1 (6): 623–9. дои : 10.1016/s1369-5274(98)80106-9 . ПМИД 10066546 .
- ^ Перейти обратно: а б Фэй П. (июнь 1992 г.). «Кислородные отношения азотфиксации у цианобактерий» . Микробиологические обзоры . 56 (2): 340–73. дои : 10.1128/MMBR.56.2.340-373.1992 . ПМЦ 372871 . ПМИД 1620069 .
- ^ Уолсби А.Е. (март 1994 г.). «Газовые пузырьки» . Микробиологические обзоры . 58 (1): 94–144. дои : 10.1128/ММБР.58.1.94-144.1994 . ПМК 372955 . ПМИД 8177173 .
- ^ Шапиро, Игорь (май 2014 г.). «Сверхбыстрая фотохимия сенсорного родопсина Anabaena: эксперимент и теория» . Biochimica et Biophysica Acta (BBA) — Биоэнергетика . 1837 (5): 589–597. дои : 10.1016/j.bbabio.2013.09.014 . ПМИД 24099700 .
- ^ Фундаментальная биология (18 марта 2016 г.). «Бактерии» .
- ^ Волк, К. Питер; Эрнст, Аннализа; Эльхай, Джефф (1994). «Метаболизм и развитие гетероцист». В Дональде А. Брайанте (ред.). Молекулярная биология цианобактерий . Достижения в области фотосинтеза и дыхания. стр. 769–823. дои : 10.1007/978-94-011-0227-8_27 . ISBN 978-0-7923-3273-2 .
- ^ Уолтер, MR; Баулд, Дж.; Брок, Т.Д. (1976). «Глава 6.2 Микробиология и морфогенез столбчатых строматолитов (Conophyton, Vacerrilla) из Хот-Спрингс в Йеллоустонском национальном парке». Строматолиты . Развитие седиментологии. Том. 20. стр. 273–310. дои : 10.1016/S0070-4571(08)71140-3 . ISBN 9780444413765 .
- ^ Джонс, Б.; Ренаут, RW; Розен, MR; Ансделл, К.М. (2002). «Кониформные строматолиты из геотермальных систем, Северный остров, Новая Зеландия». ПАЛЕОС . 17 (1): 84. Бибкод : 2002Palai..17...84J . doi : 10.1669/0883-1351(2002)017<0084:CSFGSN>2.0.CO;2 . ISSN 0883-1351 . S2CID 130120737 .
- ^ Уортон, Роберт А.; Паркер, Брюс С.; Симмонс, Джордж М. (1983). «Распространение, видовой состав и морфология водорослевых матов в озерах сухих долин Антарктики». Психология . 22 (4): 355–365. дои : 10.2216/i0031-8884-22-4-355.1 .
- ^ Уиттон, Брайан А., изд. (2012). Экология цианобактерий II . дои : 10.1007/978-94-007-3855-3 . ISBN 978-94-007-3854-6 . S2CID 46736903 .
- ^ Сталь, Лукас Дж. (2012). «Цианобактериальные маты и строматолиты». Экология цианобактерий II . стр. 65–125. дои : 10.1007/978-94-007-3855-3_4 . ISBN 978-94-007-3854-6 .
- ^ Фаррох, Париса; Шейхпур, Мойган; Касаеян, Алибахш; Асади, Хасан; Баванди, Ройя (2019). «Цианобактерии как экологически чистый ресурс для производства биотоплива: критический обзор». Биотехнологический прогресс . 35 (5): e2835. дои : 10.1002/btpr.2835 . ПМИД 31063628 . S2CID 147705730 .
- ^ Риппка, Розмари; Станье, Роджер Ю.; Дерюэль, Жозетт; Хердман, Майкл; Уотербери, Джон Б. (1979). «Родовые назначения, история штаммов и свойства чистых культур цианобактерий» . Микробиология . 111 : 1–61. дои : 10.1099/00221287-111-1-1 .
- ^ Хойчик, Э. (2000). «Скользящая подвижность цианобактерий: наблюдения и возможные объяснения». Архив микробиологии . 174 (1–2): 11–17. дои : 10.1007/s002030000187 . ПМИД 10985737 . S2CID 9927312 .
- ^ Хансгирг А. (1883) «Замечания о движениях осциллариев». Бот. , 41 : 831.
- ^ Дрюс Г. (1959) «Вклад в знание фототаксических реакций Cyanophycea». Арх. Протистенк . 104 : 389-430.
- ^ Акимицу (1951). Выделение вещества слизи в осциллаториях в зависимости от его движения» Shokubutsugaku Zasshi . 64 (751–752): 14–17 . Хосой , « .
- ^ Уолсби, А.Э. (1968). «Выделение слизи и движение сине-зеленых водорослей». Протоплазма . 65 (1–2): 223–238. дои : 10.1007/BF01666380 . S2CID 20310025 .
- ^ Хойчик, Эгберт; Баумайстер, Вольфганг (1998). «Комплекс соединительных пор, прокариотическая органелла секреции, является молекулярным мотором, лежащим в основе скользящей подвижности у цианобактерий» . Современная биология . 8 (21): 1161–1168. дои : 10.1016/S0960-9822(07)00487-3 . ПМИД 9799733 . S2CID 14384308 .
- ^ Крейг, Лиза; Пике, Майкл Э.; Тайнер, Джон А. (2004). «Структура пилуса IV типа и патогенность бактерий». Обзоры природы Микробиология . 2 (5): 363–378. дои : 10.1038/nrmicro885 . ПМИД 15100690 . S2CID 10654430 .
- ^ Дагган, Паула С.; Готтарделло, Присцилла; Адамс, Дэвид Г. (2007). «Молекулярный анализ генов Nostoc punctiforme, участвующих в биогенезе пилуса и инфицировании растений» . Журнал бактериологии . 189 (12): 4547–4551. дои : 10.1128/JB.01927-06 . ЧВК 1913353 . ПМИД 17416648 .
- ^ Риссер, Дуглас Д.; Чу, Уильям Г.; Микс, Джон К. (2014). «Генетическая характеристика thehmplocus, кластера генов, подобного хемотаксису, который регулирует развитие и подвижность гормогоний у Nostoc punctiforme» . Молекулярная микробиология . 92 (2): 222–233. дои : 10.1111/mmi.12552 . ПМИД 24533832 . S2CID 37479716 .
- ^ Перейти обратно: а б Хайатан, Бехзад; Микс, Джон К.; Риссер, Дуглас Д. (2015). «Доказательства того, что модифицированная пилоподобная система IV типа обеспечивает скользящую подвижность и секрецию полисахаридов у нитчатых цианобактерий» . Молекулярная микробиология . 98 (6): 1021–1036. дои : 10.1111/mmi.13205 . ПМИД 26331359 . S2CID 8749419 .
- ^ Шугерс, Нильс; Нюрнберг, Деннис Дж.; Валлнер, Томас; Муллино, Конрад В.; Уайльд, Аннегрет (2015). «Локализация PilB коррелирует с направлением подергивания подвижности у цианобактерии Synechocystis sp. PCC 6803» . Микробиология . 161 (5): 960–966. дои : 10.1099/mic.0.000064 . ПМИД 25721851 .
- ^ Халфен, Лоуренс Н.; Кастенхольц, Ричард В. (1970). «Скольжение сине-зеленой водоросли: возможный механизм». Природа . 225 (5238): 1163–1165. Бибкод : 1970Natur.225.1163H . дои : 10.1038/2251163a0 . ПМИД 4984867 . S2CID 10399610 .
- ^ Халфен, Лоуренс Н.; Кастенхольц, Ричард В. (1971). «Скользящая подвижность сине-зеленых водорослей Oscillatoria Princeps 1». Журнал психологии . 7 (2): 133–145. дои : 10.1111/j.1529-8817.1971.tb01492.x . S2CID 86115246 .
- ^ Чуфаг, Жоэль; Гош, Пушпита; Пог, Коннор Б.; Нан, Бэйян; Мандадапу, Кранти К. (2019). «Механизмы скользящей подвижности бактерий на мягких субстратах» . Труды Национальной академии наук . 116 (50): 25087–25096. arXiv : 1807.07529 . Бибкод : 2019PNAS..11625087T . дои : 10.1073/pnas.1914678116 . ПМК 6911197 . ПМИД 31767758 .
- ^ Шепард, Р.Н.; Самнер, ДЮ (2010). «Ненаправленное движение нитчатых цианобактерий образует сетчатые маты» . Геобиология . 8 (3): 179–190. Бибкод : 2010Gbio....8..179S . дои : 10.1111/j.1472-4669.2010.00235.x . ПМИД 20345889 . S2CID 24452272 .
- ^ Перейти обратно: а б Дэвис, Нил С.; Лю, Александр Г.; Гиблинг, Мартин Р.; Миллер, Рэндалл Ф. (2016). «Разрешение концепций и заблуждений MISS: геологический подход к текстурам поверхности осадочных пород, создаваемым микробными и абиотическими процессами» . Обзоры наук о Земле . 154 : 210–246. Бибкод : 2016ESRv..154..210D . doi : 10.1016/j.earscirev.2016.01.005 . hdl : 1983/bd67cb45-b022-4db0-be3d-b2977d2b81ab . S2CID 56345018 .
- ^ Оллвуд, Эбигейл К.; Уолтер, Малкольм Р.; Камбер, Бальц С.; Маршалл, Крейг П.; Берч, Ян В. (2006). «Строматолитовый риф раннеархейской эпохи Австралии». Природа . 441 (7094): 714–718. Бибкод : 2006Natur.441..714A . дои : 10.1038/nature04764 . ПМИД 16760969 . S2CID 4417746 .
- ^ Самнер, Дон Ю. (1997). «Позднеархейские взаимодействия кальцита и микробов: два морфологически различных микробных сообщества, которые по-разному влияли на зародышеобразование кальцита». ПАЛЕОС . 12 (4): 302–318. Бибкод : 1997Палай..12..302S . дои : 10.2307/3515333 . JSTOR 3515333 .
- ^ Тамулонис, Карлос; Каандорп, Яап (2014). «Модель нитчатых цианобактерий, приводящая к образованию сетчатого узора» . Жизнь . 4 (3): 433–456. Бибкод : 2014Life....4..433T . дои : 10.3390/life4030433 . ПМК 4206854 . ПМИД 25370380 .
- ^ Шоу, Т.; Уинстон, М.; Рупп, CJ; Клэппер, И.; Студли, П. (2004). «Общность времен упругой релаксации в биопленках» . Письма о физических отзывах . 93 (9): 098102. Бибкод : 2004PhRvL..93i8102S . doi : 10.1103/PhysRevLett.93.098102 . ПМИД 15447143 .
- ^ Перейти обратно: а б Тамулонис, Карлос; Постма, Мартен; Каандорп, Яап (2011). «Моделирование нитчатых цианобактерий раскрывает преимущества длинных и быстрых трихом для оптимизации светового воздействия» . ПЛОС ОДИН . 6 (7): e22084. Бибкод : 2011PLoSO...622084T . дои : 10.1371/journal.pone.0022084 . ПМЦ 3138769 . ПМИД 21789215 .
- ^ Перейти обратно: а б Донкор, Виктория А.; Амевовор, Дамина ХАК; Хадер, Донат-П. (1993). «Влияние тропической солнечной радиации на подвижность нитчатых цианобактерий» . ФЭМС Микробиология Экология . 12 (2): 143–147. дои : 10.1111/j.1574-6941.1993.tb00026.x .
- ^ Гарсиа-Пишель, Ферран; Мехлинг, Маргарет; Кастенхольц, Ричард В. (1994). «Суточная миграция микроорганизмов в бентосном сообществе гиперсоленого мата» . Прикладная и экологическая микробиология . 60 (5): 1500–1511. Бибкод : 1994ApEnM..60.1500G . doi : 10.1128/aem.60.5.1500-1511.1994 . ПМК 201509 . ПМИД 16349251 .
- ^ Фуранс, Од; Сола, Антони; Диестра, Элла; Раншу-Пейрус, Энтони; Эстев, Изабель; Кометт, Пьер; Дюран, Роберт (2006). «Вертикальная миграция фототрофных бактериальных популяций в гиперсоленом микробном мате из Сален-де-Жиро (Камарг, Франция)» . ФЭМС Микробиология Экология . 57 (3): 367–377. дои : 10.1111/j.1574-6941.2006.00124.x . ПМИД 16907751 .
- ^ Ричардсон, Лори Л.; Кастенхольц, Ричард В. (1987). «Вертикальные движения цианобактерии Oscillatoria terebriformis в богатом сульфидами микробном мате горячих источников» . Прикладная и экологическая микробиология . 53 (9): 2142–2150. Бибкод : 1987ApEnM..53.2142R . дои : 10.1128/aem.53.9.2142-2150.1987 . ПМК 204072 . ПМИД 16347435 .
- ^ Макбрайд, Марк Дж. (2001). «Скользящая подвижность бактерий: множественные механизмы движения клеток по поверхностям». Ежегодный обзор микробиологии . 55 : 49–75. дои : 10.1146/аннурев.микро.55.1.49 . ПМИД 11544349 .
- ^ Райхенбах, Х. (1981). «Таксономия планирующих бактерий». Ежегодный обзор микробиологии . 35 : 339–364. дои : 10.1146/annurev.mi.35.100181.002011 . ПМИД 6794424 .
- ^ Хойчик, Эгберт; Баумайстер, Вольфганг (1998). «Комплекс соединительных пор, прокариотическая органелла секреции, является молекулярным мотором, лежащим в основе скользящей подвижности у цианобактерий» . Современная биология . 8 (21): 1161–1168. дои : 10.1016/S0960-9822(07)00487-3 . ПМИД 9799733 . S2CID 14384308 .
- ^ Хойчик, Э. (2000). «Скользящая подвижность цианобактерий: наблюдения и возможные объяснения». Архив микробиологии . 174 (1–2): 11–17. дои : 10.1007/s002030000187 . ПМИД 10985737 . S2CID 9927312 .
- ^ Бхайя, Д.; Ватанабэ, Н.; Огава, Т.; Гроссман, Арканзас (1999). «Роль альтернативного сигма-фактора в подвижности и образовании ворсинок у цианобактерии Synechocystis sp. Штамм PCC6803» . Труды Национальной академии наук . 96 (6): 3188–3193. Бибкод : 1999PNAS...96.3188B . дои : 10.1073/pnas.96.6.3188 . ЧВК 15917 . ПМИД 10077659 .
- ^ Хойчик, Э.; Баумайстер, В. (1995). «Структура оболочки четырех скользящих нитчатых цианобактерий» . Журнал бактериологии . 177 (9): 2387–2395. дои : 10.1128/jb.177.9.2387-2395.1995 . ПМК 176896 . ПМИД 7730269 .
- ^ Халфен, Лоуренс Н.; Кастенхольц, Ричард В. (1971). «Скользящая подвижность сине-зеленых водорослей Oscillatoria Princeps 1». Журнал психологии . 7 (2): 133–145. дои : 10.1111/j.1529-8817.1971.tb01492.x . S2CID 86115246 .
- ^ «Усовершенствованная модель фотофобных реакций сине-зеленой водоросли, <italic>Phormidium uncinatum</italic>». Физиология растений и клеток . 1982. doi : 10.1093/oxfordjournals.pcp.a076487 .
- ^ Хедер, Донат-П. (1987). «ВЛИЯНИЕ УФ-Б-ОБЛУЧЕНИЯ НА ФОТОДВИЖЕНИЕ В ДЕСМИДЕ Cosmarium cucumis». Фотохимия и фотобиология . 46 : 121–126. дои : 10.1111/j.1751-1097.1987.tb04745.x . S2CID 97100233 .
- ^ Габай, В.Л. (1985). «Одномгновенный механизм фототаксиса у цианобактерии Phormidium uncinatum» . Письма FEMS по микробиологии . 30 (1–2): 125–129. дои : 10.1111/j.1574-6968.1985.tb00998.x .
- ^ Хэдер, Д.П. (1987) "Фотодвижение". Цианобактерии : 325-345.
- ^ Кастенхольц, Ричард В. (14 сентября 2015 г.), « Цианобактерии », Cyanobacteria , Wiley, стр. 1–2, doi : 10.1002/9781118960608.pbm00010 , ISBN 9781118960608
- ^ Ангартер, Роджер П.; Гест, Ховард (2004). «Живописные демонстрации фотосинтеза». Исследования фотосинтеза . 80 (1–3). ООО «Спрингер Сайенс энд Бизнес Медиа»: 421–425. doi : 10.1023/b:pres.0000030426.98007.6a . ISSN 0166-8595 . ПМИД 16328838 . S2CID 9453250 .