Прокариотический цитоскелет
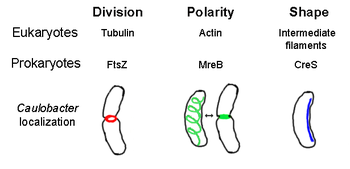
Прокариотический цитоскелет является коллективным названием для всех структурных нитей в прокариотах . Однажды считалось, что прокариотические клетки не обладали цитоскелетами , но достижения в области технологии визуализации и определения структуры привели к открытию филаментов в этих клетках в начале 1990 -х годов. [ 2 ] Также были обнаружены аналоги для всех основных цитоскелетных белков у эукариот цитоскелетные белки без известных эукариотических гомологов . , также были обнаружены [ 3 ] [ 4 ] [ 5 ] [ 6 ] Цитоскелетные элементы играют важную роль в делении клеток , защите, определении формы и определении полярности в различных прокариотах. [ 7 ] [ 8 ]
Тубулин суперсемейство
[ редактировать ]FTSZ
[ редактировать ]FTSZ, первый идентифицированный прокариотический цитоскелетный элемент, образует нитевидную кольцевую структуру, расположенную в середине клетки, называемого Z-кольцом, которое сжимает во время деления клеток , аналогично актино-миозиновому сократительному кольцу у эукариот. [ 2 ] Z-кольцо-это очень динамическая структура, которая состоит из многочисленных пучков протофиламентов, которые расширяются и сокращаются, хотя механизм сокращения Z-кольца и количество вовлеченных протофиламентов неясно. [ 1 ] FTSZ действует как белок организатора и необходим для деления клеток. Это первый компонент перегородки во время цитокинеза , и он рекрутирует все другие известные белки клеточных делений в участок деления. [ 9 ]
Несмотря на это функциональное сходство с актином , FTSZ гомологичен эукариальному тубулину . Хотя сравнение первичных структур FTSZ и тубулина выявляет слабую связь, их трехмерные структуры удивительно похожи. Кроме того, как тубулин, мономерный FTSZ связан с GTP и полимеризуется с другими мономерами FTSZ с гидролизом GTP в механизме, сходном с димеризацией тубулина . [ 10 ] Поскольку FTSZ имеет важное значение для деления клеток у бактерий, этот белок является мишенью для дизайна новых антибиотиков . [ 11 ] В настоящее время существует несколько моделей и механизмов, которые регулируют образование Z-кольца, но эти механизмы зависят от видов. Несколько видов в форме стержня, в том числе Escherichia coli и Caulobacter Crescentus , используют один или несколько ингибиторов сборки FTSZ, которые образуют биполярный градиент в клетке, усиливая полимеризацию FTSZ в клеточном центре. [ 12 ] Одна из этих систем градиента состоит из белков minCDE (см. Ниже).
Актин суперсемейство
[ редактировать ]Манеб
[ редактировать ]MREB является бактериальным белком, который, как полагают, является гомологичен эукариальным актином . MREB и актин имеют слабую первичную структуру , но очень похожи с точки зрения трехмерной структуры и полимеризации филаментов.
Почти все несферические бактерии полагаются на MREB, чтобы определить их форму. MREB собирается в спиральную сеть нитевидных структур, недалеко от цитоплазматической мембраны , покрывая всю длину клетки. [ 13 ] MREB определяет форму клеток, опосредуя положение и активность ферментов, которые синтезируют пептидогликан и действуя как жесткая нить под клеточной мембраной, которая оказывает наружное давление, чтобы лепить и поддерживать клетку. [ 1 ] MREB конденсируется из своей обычной спиральной сети и образует плотное кольцо на перегородке в Caulobacter Crescentus прямо перед делением клеток, механизм, который, как полагают, помогает найти его вне центра перегородки. [ 14 ] MREB также важен для определения полярности у полярных бактерий, поскольку он отвечает за правильное расположение как минимум четырех разных полярных белков в C. crescentus . [ 14 ]
Парм и Сопа
[ редактировать ]PARM - это цитоскелетный элемент, который обладает аналогичной структурой с актином , хотя он ведет себя функционально как тубулин . Кроме того, это полимеризует двунаправленную и демонстрирует динамическую нестабильность , которая оба являются поведением, характерными для полимеризации тубулина. [ 4 ] [ 15 ] Он образует систему с PARR и PARC , которая отвечает за разделение плазмиды R1 . PARM прикрепляется к PARR, ДНК-связывающему белу , который специфически связывается с 10 прямыми повторениями в области PARC на плазмиде R1. Это связывание происходит на обоих концах филамента PARM. Затем эта нить расширяется, разделяя плазмиды. [ 16 ] Система аналогична сегрегации эукариотической хромосом, поскольку PARM действует как эукариотический в митотическом веретке , PARR действует как кинетохорский комплекс, а PARC действует как центромер хромосомы тубулин . [ 17 ]
F -плазмида сегрегация происходит в аналогичной системе, где SOPA действует как цитоскелетная нить, а SOPB связывается с последовательности SOPC в плазмиде F, таких как кинетохор и центромер соответственно. [ 17 ] В последнее время актин-подобный гомолог PARM был обнаружен в грамположительной бактерии Bacillus thuringiensis , которая собирается в микротрубокоподобную структуру и участвует в сегрегации плазмиды . [ 18 ]
Архаальный актин
[ редактировать ]Crenactin архаального королевства - это актиновый гомолог, уникальный для термопротеты (ранее Crenarchaeota), который был обнаружен в Орденах Thermoproteales и Candidatus corarchaeum . [ 19 ] Во время его открытия в 2009 году он имеет самое высокое сходство последовательностей с эукариотическими актинами любого известного гомолога актина. [ 20 ] Кренактин был хорошо охарактеризован в Pyryobaculum calidifontis ( A3MWN5 ) и показал, что обладает высокой специфичностью для АТФ и GTP. [ 19 ] Виды, содержащие кенактин, - это стержень или игла в форме. У P. calidifontis было показано, что Crenactin образует спиральные структуры, которые охватывают длину клетки, что предполагает роль определения Crenactin в форме, аналогичной MREB у других прокариот. [ 19 ] [ 21 ]
Даже ближе к эукариотической актиновой системе обнаружено в предлагаемом суперфилу Асгардархиты . Они используют примитивные версии профилина , гельсолина и кофилина для регулирования цитоскелета. [ 22 ]
Уникальные группы
[ редактировать ]Полумесяца
[ редактировать ]Crescentin (кодируемый геном CRES ) является аналогом эукариотических промежуточных филаментов (IFS). В отличие от других аналогичных отношений, обсуждаемых здесь, Crescentin имеет довольно большую первичную гомологию с белками IF в дополнение к трехмерному сходству - последовательность CRE имеет 25% идентичность и 40% сходство с цитокератином 19 и 24% идентично 40% сходство с ядерным ламином а . Кроме того, филиалы полумесяца имеют примерно 10 нм в диаметре и, таким образом, находятся в диаметре диаметра для Eukaryal IF (8-15 нм). [ 23 ] Crescentin образует непрерывную нить от полюса к полюсу рядом с внутренней вогнутой стороной бактерий Caulobacter Crescentus в форме полумесяца . Как MREB, так и Crescentin необходимы для C. crescentus , чтобы существовать в своей характерной форме; Считается, что MREB формирует клетку в форму стержня, а полумесятель изгибает эту форму в полумесяц. [ 1 ]
Система mincde
[ редактировать ]Система MinCDE представляет собой систему филаментов, которая должным образом позиционирует перегородку в середине клетки в Escherichia coli . Согласно Shih et al., MINC ингибирует образование перегородки, запрещая полимеризацию z-кольца. MINC, MIND и MINE образуют структуру спирали, которая заманивается вокруг ячейки и привязана к мембране. Mincde Helix занимает полюс и заканчивается нитевидной структурой, называемой электронным кольцом, сделанным из моего на среднем краю полярной зоны. Из этой конфигурации электронное кольцо будет сжиматься и двигаться к этому полюсу, разбирая спираль Mincde, когда он движется вдоль. Одновременно разборные фрагменты будут пересматривать на противоположном полярном конце, реформируя катушку Mincde на противоположном полюсе, в то время как нынешняя спираль Mincde разбита. Затем этот процесс повторяется, когда спираль Mincde колеблются от полюса к полюсу. Эти колебания возникают неоднократно во время клеточного цикла, тем самым сохраняя MINC (и его ингибирующий эффект перегородки) в более низкой концентрации, усредненной по времени в середине клетки, чем на концах клетки. [ 24 ]
Динамическое поведение белков MIN было восстановлено in vitro с использованием искусственного липидного бислоя в качестве имитации для клеточной мембраны. Мои и разум самоорганизовались в параллельные и спиральные белковые волны с помощью реакционной диффузии, такой как механизм. [ 25 ]
Бактофилин
[ редактировать ]Bactofilin ( InterPro : IPR007607 ) представляет собой β-спиральный цитоскелетный элемент, который образует филаменты во всех клетках стержня Proteobacterium myxococcus xanthus . [ 26 ] Белок бактофилина, BACM, необходим для правильного поддержания формы клеток и целостности клеточной стенки. Клетки M. xanthus, в которых отсутствует бакма, имеют деформированную морфологию, характеризующуюся изогнутым клеточным телом, а мутанты BacM имеют снижение устойчивости к антибиотикам, нацеленным на бактериальную клеточную стенку. М. Ксантус Бакм белок расщепляется от его полноразмерной формы, чтобы обеспечить полимеризацию. Бактофилины были вовлечены в регуляцию формы клеток у других бактерий, включая кривизну протеус -мирабилис . [ 27 ] образование стебля Caulobacter crescentus , [ 28 ] и спиральная форма Helicobacter pylori . [ 29 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый Gitai Z (март 2005 г.). «Новая биология бактериальных клеток: движущиеся части и субклеточная архитектура» . Клетка . 120 (5): 577–86. doi : 10.1016/j.cell.2005.02.026 . PMID 15766522 . S2CID 8894304 .
- ^ Jump up to: а беременный Bi EF, Lutkenhaus J (ноябрь 1991 г.). «Структура кольца FTSZ, связанная с разделением в Escherichia coli». Природа . 354 (6349): 161–4. Bibcode : 1991natur.354..161b . doi : 10.1038/354161A0 . PMID 1944597 . S2CID 4329947 .
- ^ Gunning PW, Ghoshdastider U, Whitaker S, Popp D, Robinson RC (июнь 2015 г.). «Эволюция композиционно и функционально различных актиновых филаментов» . Журнал сотовой науки . 128 (11): 2009–19. doi : 10.1242/jcs.165563 . PMID 25788699 .
- ^ Jump up to: а беременный Попп Д., Нарита А., Ли Л.Дж., Гошдастидер У, Сюэ Б., Сринивасан Р., Баласубраманян М.К., Танака Т., Робинсон Р.К. (июнь 2012 г.). «Новая актиноподобная структура нити от Clostridium tetani» . Журнал биологической химии . 287 (25): 21121–9. doi : 10.1074/jbc.m112.341016 . PMC 3375535 . PMID 22514279 .
- ^ Попп Д., Нарита А., Гошдастидер У, Маэда К., Маэда Й., Ода Т., Фудзисава Т., Одноши Х, Ито К., Робинсон Р.К. (апрель 2010). «Полимерные структуры и динамические свойства бактериального актина альфа». Журнал молекулярной биологии . 397 (4): 1031–41. doi : 10.1016/j.jmb.2010.02.010 . PMID 20156449 .
- ^ Wickstead B, Gull K (август 2011 г.). «Эволюция цитоскелета» . Журнал клеточной биологии . 194 (4): 513–25. doi : 10.1083/jcb.201102065 . PMC 3160578 . PMID 21859859 .
- ^ Shih YL, Rothfield L (сентябрь 2006 г.). «Бактериальный цитоскелет» . Микробиология и молекулярная биология обзоры . 70 (3): 729–54. doi : 10.1128/mmbr.00017-06 . PMC 1594594 . PMID 16959967 .
- ^ Мичи К.А., Лёве Дж. (2006). «Динамические филаменты бактериального цитоскелета» (PDF) . Ежегодный обзор биохимии . 75 : 467–92. doi : 10.1146/annurev.biochem.75.103004.142452 . PMID 16756499 . Архивировано из оригинала (PDF) 17 ноября 2006 года.
- ^ Graumann PL (декабрь 2004 г.). «Цитоскелетные элементы у бактерий». Текущее мнение о микробиологии . 7 (6): 565–71. doi : 10.1016/j.mib.2004.10.010 . PMID 15556027 .
- ^ Десаи А., Митчисон Т.Дж. (июль 1998 г.). «Структуры тубулина и FTSZ: функциональные и терапевтические последствия». Биологии . 20 (7): 523–7. doi : 10.1002/(SICI) 1521-1878 (199807) 20: 7 <523 :: AID-bies1> 3.0.co; 2-l . PMID 9722999 .
- ^ Haydon DJ, Stocks NR, Ure R, Galbraith G, Bennett JM, Brown DR, Baker PJ, Barryin VV, Rice DW, Sedelnikova SE, Heal JR, Sheridan JM, Aiwale St, Chauhan PK, Srivastava A, Taneja A, Collins I. , Errington J, Czaplewski LG (сентябрь 2008 г.). «Ингибитор FTSZ с мощной и селективной антистафилококковой активностью» Наука . 321 (5896): 1673–5 Bibcode : 2008Sci ... 321.1673H Doi : 10.1126/ science.1 18801997PMID 7878853S2CID
- ^ Heausser DP, Margolin W (апрель 2016 г.). «Splitsville: структурное и функциональное понимание динамического бактериального Z -кольца» . Природные обзоры. Микробиология . 14 (5): 305–19. doi : 10.1038/nrmicro.2016.26 . PMC 5290750 . PMID 27040757 .
- ^ Kürner J, Medalia O, Linaroudis AA, Baumeister W (ноябрь 2004 г.). «Новое понимание структурной организации эукариотических и прокариотических цитоскелетов с использованием криоэлектронной томографии». Экспериментальные исследования клеток . 301 (1): 38–42. doi : 10.1016/j.yexcr.2004.08.005 . PMID 15501443 .
- ^ Jump up to: а беременный Gitai Z, Dye N, Shapiro L (июнь 2004 г.). «Актин-подобный ген может определять полярность клеток у бактерий» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (23): 8643–8. doi : 10.1073/pnas.0402638101 . PMC 423248 . PMID 15159537 .
- ^ Garner EC, Campbell CS, Mullins Rd (ноябрь 2004 г.). «Динамическая нестабильность в германизирующем прокариотическом гомологе актина». Наука . 306 (5698): 1021–5. Bibcode : 2004sci ... 306.1021G . doi : 10.1126/science.1101313 . PMID 15528442 . S2CID 14032209 .
- ^ Møller-Jensen J, Jensen RB, Löwe J, Gerdes K (июнь 2002 г.). «Прокариотическая сегрегация ДНК актин-подобной нити» . Embo Journal . 21 (12): 3119–27. doi : 10.1093/emboj/cdf320 . PMC 126073 . PMID 12065424 .
- ^ Jump up to: а беременный Gitai Z (февраль 2006 г.). «Сегрегация плазмиды: появляется новый класс цитоскелетных белков» . Текущая биология . 16 (4): R133-6. Bibcode : 2006cbio ... 16.r133g . doi : 10.1016/j.cub.2006.02.007 . PMID 16488865 .
- ^ Цзян С., Нарита А., Попп Д., Гошдастидер У, Ли Л.Дж., Шринивасан Р., Баласубраманян М.К., Ода Т., Ко Ф., Ларссон М., Робинсон Р.К. (март 2016 г.). «Новые актиновые филаменты из Bacillus thuringiensis образуют нанотрубочки для сегрегации плазмидной ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 113 (9): E1200-5. BIBCODE : 2016PNAS..113E1200J . doi : 10.1073/pnas.1600129113 . PMC 4780641 . PMID 26873105 .
- ^ Jump up to: а беременный в Ettema TJ, Lindås AC, Bernander R (май 2011 г.). «Цитоскелет на основе актина в археи» . Молекулярная микробиология . 80 (4): 1052–61. doi : 10.1111/j.1365-2958.2011.07635.x . PMID 21414041 .
- ^ Ютин Н., Вольф Мей, Вольф Йи, Кунин Э.В. (февраль 2009 г.). «Происхождение фагоцитоза и эукариогенеза» . Биология прямой . 4 : 9. doi : 10.1186/1745-6150-4-9 . PMC 2651865 . PMID 19245710 .
- ^ Ghoshdastider U, Jiang S, Popp D, Robinson RC (июль 2015 г.). «В поисках изначальной накаливания актина» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (30): 9150–1. doi : 10.1073/pnas.1511568112 . PMC 4522752 . PMID 26178194 .
- ^ Akıl, Caner; Тран, Линх Т.; Orhant-Prioux, Magali; Баскаран, Йохендран; Мансер, Эдвард; Бланхон, Лоран; Робинсон, Роберт С. (18 августа 2020 г.). «Понимание эволюции регулируемой динамики актина посредством характеристики примитивных белков гельсолина/кофилина из Asgard Archaea» . ПНА . 117 (33): 19904–19913. Bibcode : 2020pnas..11719904a . Biorxiv 10.1101/768580 . doi : 10.1073/pnas.2009167117 . PMC 7444086 . PMID 32747565 .
- ^ Ausmees N, Kuhn Jr, Jacobs-Wagner C (декабрь 2003 г.). «Бактериальный цитоскелет: промежуточная функция, похожая на нити в форме клеток» . Клетка . 115 (6): 705–13. doi : 10.1016/s0092-8674 (03) 00935-8 . PMID 14675535 . S2CID 14459851 .
- ^ Shih YL, Le T, Rothfield L (июнь 2003 г.). «Выбор сайта дивизии в Escherichia coli включает в себя динамическое перераспределение мин белков внутри спиральных структур, которые простираются между двумя полюсами клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (13): 7865–70. Bibcode : 2003pnas..100.7865S . doi : 10.1073/pnas.1232225100 . PMC 164679 . PMID 12766229 .
- ^ Ship M, Fischer-Friedrich E, Ries J, Kruse K, Schwille P (май 2008 г.). «Пространственные регуляторы для деления бактериальных клеток самоорганизуются в поверхностные волны in vitro». Наука . 320 (5877): 789–92. Bibcode : 2008Sci ... 320..789L . doi : 10.1126/science.11544413 . PMID 18467587 . S2CID 27134918 .
- ^ Koch Mk, McHugh CA, Hoiczyk E (май 2011 г.). «BACM, N-концевой обработанный бактофилин Myxococcus xanthus, имеет решающее значение для правильной формы клеток» . Молекулярная микробиология . 80 (4): 1031–51. doi : 10.1111/j.1365-2958.2011.07629.x . PMC 3091990 . PMID 21414039 .
- ^ Hay NA, Tipper DJ, Gygi D, Hughes C (апрель 1999 г.). «Новый мембранный белок, влияющий на форму клеток и многоклеточное роя Proteus mirabilis» . Журнал бактериологии . 181 (7): 2008–16. doi : 10.1128/jb.181.7.2008-2016.1999 . PMC 93611 . PMID 10094676 .
- ^ Kühn J, Briegel A, Mörschel E, Kahnt J, Leser K, Wick S, Jensen GJ, Thanbichler M (январь 2010 г.). «Бактофилины, вездесущий класс цитоскелетных белков, опосредующих полярную локализацию синтазы клеточной стенки в Caulobacter Crescentus» . Embo Journal . 29 (2): 327–39. doi : 10.1038/emboj.2009.358 . PMC 2824468 . PMID 19959992 .
- ^ Sycuro LK, Pincus Z, Gutierrez KD, Biboy J, Stern CA, Vollmer W, Salama NR (май 2010 г.). «Пептидогликан сшивающая релаксация способствует спиральной форме Helicobacter Pylori и колонизации желудка» . Клетка . 141 (5): 822–33. doi : 10.1016/j.cell.2010.03.046 . PMC 2920535 . PMID 20510929 .