Гауссовская сетевая модель

Модель гауссовой сети (GNM) представляет собой представление биологической макромолекулы как упругой сети масс и пружин для изучения, понимания и характеристики механических аспектов ее долгосрочной крупномасштабной динамики . Модель имеет широкий спектр применений: от небольших белков, таких как ферменты, состоящие из одного домена , до крупных макромолекулярных ансамблей, таких как рибосома или вирусный капсид . Динамика белковых доменов играет ключевую роль во множестве процессов молекулярного распознавания и передачи сигналов в клетках .Белковые домены, соединенные внутренне неупорядоченными гибкими линкерными доменами, индуцируют дальнюю аллостерию посредством динамики белковых доменов .Получающиеся в результате динамические режимы обычно не могут быть предсказаны на основе статических структур ни всего белка, ни отдельных доменов.
Сетевая модель Гаусса — это минималистский, крупнозернистый подход к изучению биологических молекул. В модели белки представлены узлами, соответствующими α-углеродам аминокислотных остатков. Аналогично структуры ДНК и РНК представлены от одного до трех узлов на каждый нуклеотид . Модель использует гармоническое приближение для моделирования взаимодействий. Такое крупнозернистое представление делает вычисления недорогими в вычислительном отношении.
На молекулярном уровне многие биологические явления, такие как каталитическая активность фермента , происходят в диапазоне от нано-до миллисекунд. Все методы моделирования атомов, такие как моделирование молекулярной динамики , редко достигают микросекундной длины траектории, в зависимости от размера системы и доступных вычислительных ресурсов. Анализ нормального режима в контексте моделей GNM или моделей упругой сети (EN) в целом дает представление о более крупномасштабном функциональном динамическом поведении макромолекул. Здесь модель отражает функциональные движения биомолекулы в естественном состоянии за счет атомных деталей. Выводы, полученные на основе этой модели, дополняют методы моделирования атомарных деталей.
Другая модель динамики белков, основанная на упругих сетях масс и пружин, — это модель анизотропной сети .
Теория модели гауссовской сети
[ редактировать ]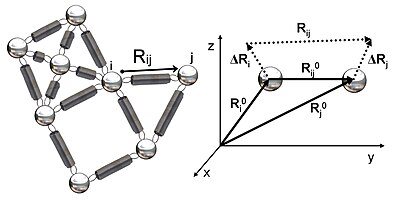
Модель гауссовской сети была предложена Бахаром, Атилганом, Халилоглу и Эрманом в 1997 году. [1] [2] GNM часто анализируется с использованием анализа нормального режима, который предлагает аналитическую формулировку и уникальное решение для каждой структуры. Анализ нормального режима GNM отличается от других анализов нормального режима тем, что он основан исключительно на топологии контакта между остатками, находящейся под влиянием теории упругости Флори. [3] и модель Роуз [4] и не учитывает трехмерную направленность движений.
Представление структуры в виде эластичной сети
[ редактировать ]На рисунке 2 показано схематическое изображение эластичной сети, изучаемой в GNM. Металлические бусины представляют собой узлы этой гауссовой сети (остатки белка), а пружины представляют собой связи между узлами (ковалентные и нековалентные взаимодействия между остатками). Для узлов i и j векторы положения равновесия R 0 я и Р 0 j , вектор равновесного расстояния, R 0 ij , мгновенные векторы колебаний, ΔR i и ΔR j , и мгновенный вектор расстояния, R ij , показаны на рисунке 2. Мгновенные векторы положения этих узлов определяются R i и R j . Разница между вектором положения равновесия и вектором мгновенного положения остатка i дает вектор мгновенных колебаний ΔR i = R i - R. 0 я . Следовательно, мгновенный вектор колебаний между узлами i и j выражается как ΔR ij = ΔR j - ΔR i = R ij - R 0 я .
Потенциал гауссовой сети
[ редактировать ]Потенциальная энергия сети через ΔR i равна
![{\displaystyle V_{GNM}={\frac {\gamma }{2}}\left[\sum _{i,j}^{N}(\Delta R_{j}-\Delta R_{i})^ {2}\right]={\frac {\gamma }{2}}\left[\sum _{i,j}^{N}\Delta R_{i}\Gamma _{ij}\Delta R_{j }\верно]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/d5be28f13b07a00cf754058700cee674e9badb75)
где γ — силовая постоянная, равномерная для всех пружин, а Γ ij — ij -й элемент матрицы Кирхгофа (или матрицы связности) контактов между остатками, Γ , определяемый формулой

r c представляет собой предельное расстояние для пространственных взаимодействий и принимается равным 7 Å для пар аминокислот (представленных их α-углеродами).
Выражая компоненты X, Y и Z векторов колебаний ΔR i как ΔX Т = [ΔX 1 ΔX 2 ..... ΔX N ], ΔY Т = [ΔY 1 ΔY 2 ..... ΔY N ], и ΔZ Т = [ΔZ 1 ΔZ 2 ..... ΔZ N ], приведенное выше уравнение упрощается до
![{\displaystyle V_{GNM}={\frac {\gamma }{2}}[\Delta X^{T}\Gamma \Delta X+\Delta Y^{T}\Gamma \Delta Y+\Delta Z^{T }\Гамма \Дельта Z]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/e69f61521a50a6c344b039f8d8c5e74a141c239c)
Основы статистической механики
[ редактировать ]В GNM распределение вероятностей всех флуктуаций P ( ΔR ) изотропно.

и гауссовский

где k B — постоянная Больцмана, а T — абсолютная температура. p ( ΔY ) и p ( ΔZ ) выражаются аналогично. N-мерная функция плотности вероятности Гаусса с вектором случайной величины x , средним вектором µ и ковариационной матрицей Σ равна

нормализует распределение и |Σ| является определителем ковариационной матрицы.

Подобно распределению Гаусса, нормализованное распределение для ΔX Т = [ΔX 1 ΔX 2 ..... ΔX N ] вокруг положений равновесия можно выразить как

Константа нормализации, а также статистическая сумма Z X , определяется выражением

где в данном случае это ковариационная матрица. Z Y и Z Z выражаются аналогично. Эта формулировка требует обращения матрицы Кирхгофа. В GNM определитель матрицы Кирхгофа равен нулю, поэтому вычисление ее обратного требует разложения по собственным значениям . Γ −1 строится с использованием N-1 ненулевых собственных значений и связанных с ними собственных векторов. Выражения для p ( ΔY ) и p ( ΔZ ) аналогичны выражениям для p ( ΔX ). Распределение вероятностей всех колебаний в GNM становится


Для этой системы масс и пружин константа нормализации в предыдущем выражении представляет собой общую статистическую сумму GNM, Z GNM ,

Ожидаемые значения колебаний и корреляций
[ редактировать ]Ожидаемые значения колебаний остатка, < ΔR i 2 > (также называемые среднеквадратичными флуктуациями, MSF) и их взаимные корреляции < ΔR i · ΔR j > могут быть организованы как диагональные и недиагональные члены, соответственно, ковариационной матрицы. На основе статистической механики ковариационная матрица для ΔX определяется выражением

Последнее равенство получается путем подстановки приведенного выше p( ∆X ) и принятия (обобщенного гауссова) интеграла. С,

< ΔR я 2 > и < ΔR i · ΔR j > следует


Разложение по модам
[ редактировать ]Нормальные моды ГНМ находятся путем диагонализации матрицы Кирхгофа Γ = UΛU Т . Здесь U — унитарная матрица, U Т = У −1 собственных векторов u i Γ Λ и i является диагональной матрицей собственных значений λ . , Частота и форма моды представлены ее собственным значением и собственным вектором соответственно. Поскольку матрица Кирхгофа положительно полуопределена, первое собственное значение λ 1 равно нулю, а все элементы соответствующего собственного вектора равны 1/ √ N . Это показывает, что сетевая модель трансляционно инвариантна.
Взаимные корреляции между флуктуациями остатков можно записать в виде суммы по N-1 ненулевым модам как
![{\displaystyle <\Delta R_{i}\cdot \Delta R_{j}>={\frac {3k_{B}T}{\gamma }}[U\Lambda ^{-1}U^{T}] _{ij}={\frac {3k_{B}T}{\gamma }}\sum _{k=1}^{N-1}\lambda _{k}^{-1}[u_{k} u_{k}^{T}]_{ij}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/cba38d6c9778162a706d15d897151e128670cd23)
Отсюда следует, что [ ΔR i · ΔR j ] вклад отдельной моды выражается как
![{\displaystyle [\Delta R_{i}\cdot \Delta R_{j}]_{k}={\frac {3k_{B}T}{\gamma }}\lambda _{k}^{-1} [u_{k}]_{i}[u_{k}]_{j}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/8284898bdd32ed64bd4ca2eb3c5274540809d2b4)
[ uk ] где i — й элемент uk i - .
Влияние локальной плотности упаковки
[ редактировать ]По определению, диагональный элемент матрицы Кирхгофа, Γ ii , равен степени узла в GNM, который представляет координационное число соответствующего остатка. Это число является мерой локальной плотности упаковки вокруг данного остатка. Влияние локальной плотности упаковки можно оценить путем разложения Γ в ряд −1 матрица. Γ можно записать как сумму двух матриц Γ = D + O , содержащих диагональные элементы и недиагональные элементы Γ .
- С −1 = ( Д + О ) −1 = [ Д ( Я + Д −1 О ) ] −1 = ( Я + Д −1 О ) −1 Д −1 = ( Я - Д −1 О +...) Д −1 = Д −1 - Д −1 Д О −1 + ...
Это выражение показывает, что локальная плотность упаковки вносит существенный вклад в ожидаемые колебания остатков. [5] Члены, которые следуют за обратной диагональной матрицей, представляют собой вклад позиционных корреляций в ожидаемые колебания.
Приложения ГНМ
[ редактировать ]
Колебания равновесия
[ редактировать ]Равновесные колебания биологических молекул можно измерить экспериментально. В рентгеновской кристаллографии B-фактор (также называемый фактором Дебая-Валлера или температурным фактором) каждого атома является мерой его среднеквадратичного отклонения вблизи положения равновесия в собственной структуре. В экспериментах ЯМР эту меру можно получить путем расчета среднеквадратических различий между различными моделями.Во многих приложениях и публикациях, включая оригинальные статьи, было показано, что ожидаемые флуктуации остатков, полученные с помощью GNM, хорошо согласуются с экспериментально измеренными флуктуациями собственного состояния. [6] [7] Например, связь между B-факторами и ожидаемыми колебаниями остатков, полученными из GNM, следующая:

На рисунке 3 показан пример расчета GNM для каталитического домена белка Cdc25B, цикла деления клеток фосфатазы двойной специфичности .

Физический смысл медленных и быстрых режимов
[ редактировать ]Диагонализация матрицы Кирхгофа разлагает конформационные движения на спектр коллективных мод. Ожидаемые значения флуктуаций и взаимных корреляций получаются из линейных комбинаций флуктуаций вдоль этих нормальных мод. Вклад каждой моды масштабируется пропорционально частоте этой моды. Следовательно, медленные (низкочастотные) моды вносят наибольший вклад в ожидаемые флуктуации. Показано, что среди нескольких самых медленных режимов движения являются коллективными и глобальными и потенциально имеют отношение к функциональности биомолекул. С другой стороны, быстрые (высокочастотные) моды описывают некоррелированные движения, не вызывающие заметных изменений в структуре. Методы, основанные на ГНМ, не обеспечивают реальной динамики, а лишь аппроксимацию, основанную на сочетании и интерполяции нормальных режимов. [8] Их применимость сильно зависит от того, насколько коллективным является движение. [8] [9]
Другие специальные приложения
[ редактировать ]Существует несколько основных областей, в которых модель гауссовой сети и другие модели эластичных сетей оказались полезными. [10] К ним относятся:
- Сетевая модель на основе пружинных шариков . В сетевой модели на основе пружинных шариков пружины и шарики используются в качестве компонентов в сшитой сети. Пружины сшиты для представления механического поведения материала и модели молекулярной динамики моста (MD) и модели конечных элементов (FE) (см. Рисунок 5). Шарики представляют собой материальную массу кластерных связей. Каждая пружина используется для обозначения группы полимерных цепей, а не части одной полимерной цепи. Это упрощение позволяет объединить различные модели в разных масштабах и значительно повышает эффективность моделирования. На каждой итерации моделирования силы пружин прикладываются к узлам в центре буртиков и рассчитываются уравновешенные узловые перемещения по всей системе. В отличие от традиционного метода КЭ для получения напряжений и деформаций, модель «пружина-буртик» обеспечивает перемещения узлов и силы в пружинах. Эквивалентную деформацию и энергию деформации модели сети на основе пружины-шарика можно определить и рассчитать с использованием смещений узлов и характеристик пружины. Кроме того, результаты сетевой модели можно масштабировать для получения структурной реакции на макроуровне с использованием анализа конечных элементов. [11] [12]
- Разложение гибких/жестких областей и доменов белков [13] [14] [15]
- Характеристика функциональных движений и функционально важных участков/остатков белков, ферментов и крупных макромолекулярных ансамблей. [16] [11] [17] [18] [19] [20] [21] [22] [23] [24] [25] [26]
- Уточнение и динамика структурных данных с низким разрешением, например, криоэлектронная микроскопия. [27] [28] [29] [30]
- Молекулярная замена для решения рентгеновских структур , когда произошло конформационное изменение по отношению к известной структуре. [31]
- Интеграция с атомистическими моделями и симуляциями [32] [33]
- Исследование путей и кинетики складывания/раскладывания. [34] [35]
- Аннотация функционального значения молекулярной эволюции [36] [37]
Веб-серверы
[ редактировать ]На практике можно выполнить два вида вычислений.Первый тип (GNM как таковой) использует матрицу Кирхгофа . [1] [2] Второй тип (более конкретно называемый либо моделью эластичной сети, либо моделью анизотропной сети) использует матрицу Гессе, связанную с соответствующим набором гармонических пружин. [38] Оба типа моделей можно использовать онлайн, используя следующие серверы.
ГНМ-серверы
[ редактировать ]- iGNM: база данных функциональных движений белков на основе GNM http://ignm.ccbb.pitt.edu [39]
- oGNM: Онлайн-расчет структурной динамики с использованием GNM https://web.archive.org/web/20070516042756/http://ignm.ccbb.pitt.edu/GNM_Online_Calculation.htm
Серверы ENM/ANM
[ редактировать ]- модели анизотропной сети Веб-сервер http://www.ccbb.pitt.edu/anm [40]
- elNemo: веб-интерфейс к модели эластичной сети http://www.sciences.univ-nantes.fr/elnemo/
- AD-ENM: Анализ динамики модели эластичной сети http://enm.lobos.nih.gov/
- WEBnm@: Веб-сервер для анализа белков в нормальном режиме http://apps.cbu.uib.no/webnma/home
Другие соответствующие серверы
[ редактировать ]- ProDy: интерфейс прикладного программирования (API) на Python, который объединяет анализы GNM и ANM, а также несколько инструментов анализа молекулярной структуры и последовательностей, а также инструментов визуализации: http://prody.csb.pitt.edu [41] [42]
- HingeProt: алгоритм прогнозирования белковых шарниров с использованием моделей эластичных сетей http://www.prc.boun.edu.tr/appserv/prc/hingeprot/ или http://bioinfo3d.cs.tau.ac.il/HingeProt/ шарнирпрот.html
- DNABindProt: сервер для определения потенциальных сайтов связывания белков с ДНК http://www.prc.boun.edu.tr/appserv/prc/dnabindprot/
- MolMovDB: База данных макромолекулярных движений: http://www.molmovdb.org/
См. также
[ редактировать ]- Гауссово распределение
- Гармонический осциллятор
- Закон Гука
- Молекулярная динамика
- Обычный режим
- Анализ главных компонентов
- Динамика белка
- Эластичность резины
- Статистическая механика
Ссылки
[ редактировать ]Первоисточники
[ редактировать ]- Бахар, И.; Атилган, Арканзас; Эрман, Б. (1997). «Прямая оценка тепловых колебаний белка с использованием гармонического потенциала одного параметра». Складывание и дизайн . 2 (3): 173–181. дои : 10.1016/s1359-0278(97)00024-2 . ПМИД 9218955 .
- Халилоглу, Т. Бахар; Эрман, Б. (1997). «Гауссова динамика свернутых белков». Физ. Преподобный Летт . 79 (16): 3090–3093. Бибкод : 1997PhRvL..79.3090H . дои : 10.1103/physrevlett.79.3090 .
- Цуй К, Бахар I (2006). Анализ в нормальном режиме: теория и приложения к биологическим и химическим системам, Chapman & Hall/CRC, Лондон, Великобритания.
Конкретные цитаты
[ редактировать ]- ^ Перейти обратно: а б Бахар, И.; Атилган, Арканзас; Эрман, Б. (1997). «Прямая оценка тепловых колебаний белка с использованием гармонического потенциала одного параметра». Складывание и дизайн . 2 (3): 173–181. дои : 10.1016/s1359-0278(97)00024-2 . ПМИД 9218955 .
- ^ Перейти обратно: а б Халилоглу, Т. Бахар; Эрман, Б (1997). «Гауссова динамика свернутых белков». Физ. Преподобный Летт . 79 (16): 3090–3093. Бибкод : 1997PhRvL..79.3090H . дои : 10.1103/physrevlett.79.3090 .
- ^ Флори, Пи Джей (1976). «Статистическая термодинамика случайных сетей». Учеб. Р. Сок. Лонд. А. 351 (1666): 351. Бибкод : 1976RSPSA.351..351F . дои : 10.1098/rspa.1976.0146 . S2CID 122325882 .
- ^ Роуз, ЧП (1953). «Теория линейных вязкоупругих свойств разбавленных растворов спиральных полимеров». Дж. Хим. Физ . 21 (7): 1272. Бибкод : 1953ЖЧФ..21.1272Р . дои : 10.1063/1.1699180 .
- ^ Галле, Б. (2002). «Гибкость и упаковка в белках» . Учеб. Натл. акад. наук. США . 99 (3): 1274–1279. Бибкод : 2002PNAS...99.1274H . дои : 10.1073/pnas.032522499 . ПМК 122180 . ПМИД 11818549 .
- ^ Бахар, И.; Валлквист, А.; Ковелл, Д.Г.; Джерниган, РЛ (1998). «Корреляция между водородным обменом в естественном состоянии и кооперативными колебаниями остатков из простой модели». Биохимия . 37 (4): 1067–1075. CiteSeerX 10.1.1.551.9055 . дои : 10.1021/bi9720641 . ПМИД 9454598 .
- ^ Бахар, И.; Атилган, Арканзас; Демирель, MC; Эрман, Б. (1998). «Колебательная динамика белков: значение медленных и быстрых режимов в отношении функции и стабильности». Физ. Преподобный Летт . 80 (12): 2733–2736. Бибкод : 1998PhRvL..80.2733B . дои : 10.1103/physrevlett.80.2733 . S2CID 1070176 .
- ^ Перейти обратно: а б Кмичик, Себастьян; Коуза, Максим; Бадачевска-Давид, Александра Е.; Клочковски, Анджей; Колинский, Анджей (2018). «Моделирование структурной гибкости белка и крупномасштабной динамики: крупномасштабное моделирование и модели эластичных сетей» . Международный журнал молекулярных наук . 19 (11): 3496. doi : 10.3390/ijms19113496 . ПМК 6274762 . ПМИД 30404229 .
- ^ Ян, Лей; Сун, Гуан; Джерниган, Роберт Л. (1 августа 2007 г.). «Насколько хорошо мы можем понять крупномасштабные движения белков, используя обычные режимы моделей эластичных сетей?» . Биофизический журнал . 93 (3): 920–929. Бибкод : 2007BpJ....93..920Y . дои : 10.1529/biophysj.106.095927 . ISSN 0006-3495 . ЧВК 1913142 . ПМИД 17483178 .
- ^ Ченнубхотла, К; Рейдер, Эй Джей; Ян, ЛВ; Бахар, я (2005). «Модели эластичной сети для понимания биомолекулярных механизмов: от ферментов до супрамолекулярных сборок». Физ. Биол . 2 (4): С173–С180. Бибкод : 2005PhBio...2S.173C . дои : 10.1088/1478-3975/2/4/S12 . ПМИД 16280623 . S2CID 28083452 .
- ^ Перейти обратно: а б Чжан, Цзиньцзюнь (2015). «Оптимизированная модель сшитой сети для моделирования реакции линейно-упругого материала интеллектуального полимера» . Журнал интеллектуальных материальных систем и структур . 27 (11): 1461–1475. дои : 10.1177/1045389X15595292 . S2CID 137709230 .
- ^ Чжан, Цзиньцзюнь (2015). «Новая статистическая сетевая модель на основе пружинных шариков для самочувствительных интеллектуальных полимерных материалов» . Умные материалы и конструкции . 24 (8): 085022. Бибкод : 2015SMaS...24h5022Z . дои : 10.1088/0964-1726/24/8/085022 . hdl : 2286/RI35587 . S2CID 55578558 .
- ^ Хинсен, К. (1999). «Анализ движений доменов путем приближенных расчетов в нормальном режиме». Белки . 33 (3): 417–429. doi : 10.1002/(sici)1097-0134(19981115)33:3<417::aid-prot10>3.0.co;2-8 . ПМИД 9829700 . S2CID 33336590 .
- ^ Рейдер, AJ; Андерсон, Г.; Исин, Б.; Корана, Х.Г.; Бахар, И.; Кляйн-Ситхараман, Дж. (2004). «Идентификация основных аминокислот, стабилизирующих родопсин» . Учеб. Натл. акад. наук. США . 101 (19): 7246–7251. Бибкод : 2004PNAS..101.7246R . дои : 10.1073/pnas.0401429101 . ПМК 409904 . ПМИД 15123809 .
- ^ Кунду, С.; Соренсен, округ Колумбия; Филлипс, Дж. Н. младший (2004). «Автоматическое доменное разложение белков с помощью модели гауссовой сети». Белки . 57 (4): 725–733. дои : 10.1002/прот.20268 . ПМИД 15478120 . S2CID 9600056 .
- ^ Чжан, Цзиньцзюнь (2015). «Новая статистическая сетевая модель на основе пружинных шариков для самочувствительных интеллектуальных полимерных материалов» . Умные материалы и конструкции . 24 (8): 085022. Бибкод : 2015SMaS...24h5022Z . дои : 10.1088/0964-1726/24/8/085022 . hdl : 2286/RI35587 . S2CID 55578558 .
- ^ Кескин, О.; и др. (2002). «Связь молекулярной гибкости с функцией: пример тубулина» . Биофиз. Дж . 83 (2): 663–80. Бибкод : 2002BpJ....83..663K . дои : 10.1016/s0006-3495(02)75199-0 . ПМК 1302177 . ПМИД 12124255 .
- ^ Темиз Н.А. и Бахар I, Связывание ингибитора изменяет направления движения домена в обратной транскриптазе ВИЧ-1, Белки: структура, функции и генетика 49, 61-70, 2002.
- ^ Сюй, К., Тоби, Д. и Бахар, И. 2003 Аллостерические изменения в структуре белка, рассчитанные с помощью простой механической модели: переход гемоглобина T <-> R2, J. Mol. Биол. , 333, 153.
- ^ Дрор Тоби и Ивет Бахар, Структурные изменения, связанные с связыванием белков, коррелируют с внутренними движениями белков в несвязанном состоянии, Proc Natl Acad Sci (США) 102, 18908-18913, 2005.
- ^ Шривастава, Индира Х.; Бахар, Ивет (2006). «Общий механизм открытия пор, общий для пяти различных калиевых каналов» . Биофиз Дж . 90 (11): 3929–3940. Бибкод : 2006BpJ....90.3929S . doi : 10.1529/biophysj.105.080093 . ПМЦ 1459499 . ПМИД 16533848 .
- ^ Ян, ЛВ; Бахар, я (2005). «Связь между каталитическим центром и коллективной динамикой: требование механохимической активности ферментов» . Структура . 13 (6): 893–904. дои : 10.1016/j.str.2005.03.015 . ПМК 1489920 . ПМИД 15939021 .
- ^ Ченнубхотла, Чакра; Бахар, Ивет (2006). «Марковские методы для иерархического грубого анализа динамики крупных белков» . Исследования в области вычислительной молекулярной биологии . Конспекты лекций по информатике. Том. 3909. С. 379–393 . дои : 10.1007/11732990_32 . ISBN 978-3-540-33295-4 .
- ^ Ван, Ю. Рейдер; Эй Джей; Бахар, И.; Джерниган, РЛ. (2004). «Глобальные движения рибосом, выявленные с помощью модели эластичной сети». Дж. Структ. Биол . 147 (3): 302–314. дои : 10.1016/j.jsb.2004.01.005 . ПМИД 15450299 .
- ^ Рейдер, Эй Джей; Влад, Даниил; Бахар, Ивет (2005). «Динамика созревания капсида бактериофага HK97» . Структура . 13 (3): 413–21. дои : 10.1016/j.str.2004.12.015 . ПМИД 15766543 .
- ^ Хамахер, К.; Трильска, Дж.; Маккаммон, Дж. А. (2006). «Карта зависимости белков малой рибосомальной субъединицы» . ПЛОС Компьютер. Биол . 2 (2): е10. Бибкод : 2006PLSCB...2...10H . дои : 10.1371/journal.pcbi.0020010 . ПМК 1364506 . ПМИД 16485038 .
- ^ Мин, Д.; и др. (2002). «Как описать движение белка без аминокислотной последовательности и координат атомов» . Учеб. Натл. акад. наук. США . 99 (13): 8620–8625. Бибкод : 2002PNAS...99.8620M . дои : 10.1073/pnas.082148899 . ПМЦ 124334 . ПМИД 12084922 .
- ^ Тама, Ф.; Риггерс, В.; Брукс III, CL (2002). «Изучение глобальных искажений биологических макромолекул и агрегатов на основе структурной информации низкого разрешения и теории эластичных сетей». Дж. Мол. Биол . 321 (2): 297–305. CiteSeerX 10.1.1.457.8 . дои : 10.1016/s0022-2836(02)00627-7 . ПМИД 12144786 .
- ^ Деларю, М.; Дюма, П. (2004). «Об использовании низкочастотных нормальных мод для усиления коллективных движений при уточнении структурных моделей макромолекул» . Учеб. Натл. акад. наук. США . 101 (18): 6957–6962. Бибкод : 2004PNAS..101.6957D . дои : 10.1073/pnas.0400301101 . ПМК 406448 . ПМИД 15096585 .
- ^ Микелетти, К.; Карлони, П.; Маритан, А. (2004). « Точное и эффективное описание колебательной динамики белков» в сравнении молекулярной динамики и гауссовских моделей». Белки . 55 (3): 635–45. arXiv : cond-mat/0405145 . Бибкод : 2004cond.mat..5145M . дои : 10.1002/прот.20049 . ПМИД 15103627 . S2CID 5348611 .
- ^ Зуре, К.; Санеджуанд, Ю.Х. (2004). «О возможностях анализа в нормальном режиме для решения сложных проблем молекулярного замещения». Акта Кристаллогр. Д. 60 (4): 796–9. дои : 10.1107/s0907444904001982 . ПМИД 15039589 .
- ^ Чжан, ЗЯ; Ши, ГГ; Лю, HY (2003). «Молекулярно-динамическое моделирование пептидов и белков с усиленными коллективными движениями» . Биофиз. Дж . 84 (6): 3583–93. Бибкод : 2003BpJ....84.3583Z . дои : 10.1016/s0006-3495(03)75090-5 . ПМК 1302944 . ПМИД 12770868 .
- ^ Микелетти, К.; Латтанци, Г.; Маритан, А. (2002). «Эластичные свойства белков: понимание процесса сворачивания и эволюционного отбора нативных структур». Дж. Мол. Биол . 321 (5): 909–21. arXiv : cond-mat/0204400 . Bibcode : 2002cond.mat..4400M . дои : 10.1016/s0022-2836(02)00710-6 . ПМИД 12206770 . S2CID 8492131 .
- ^ Микелетти, К.; и др. (2002). «Важнейшие этапы сворачивания белка с помощью решаемой модели: предсказание сайтов-мишеней для препаратов, ингибирующих ферменты» . Белковая наука . 11 (8): 1878–87. arXiv : cond-mat/0209325 . Бибкод : 2002cond.mat..9325M . дои : 10.1110/ps.3360102 . ПМЦ 2373687 . ПМИД 12142442 .
- ^ Портман, Джей-Джей; Такада, С.; Волинс, П.Г. (2001). «Микроскопическая теория скорости сворачивания белков. I. тонкая структура профиля свободной энергии и пути сворачивания с помощью вариационного подхода». Дж. Хим. Физ . 114 (11): 5069. arXiv : cond-mat/0008454 . Бибкод : 2001JChPh.114.5069P . дои : 10.1063/1.1334662 . S2CID 14699178 .
- ^ Хамахер, К. (2008). «Связь эволюции последовательности протеазы ВИЧ1 с ее основной молекулярной механикой». Джин . 422 (1–2): 30–36. дои : 10.1016/j.gene.2008.06.007 . ПМИД 18590806 .
- ^ Хамахер, К.; Маккаммон, Дж. А. (2006). «Вычисление аминокислотной специфичности колебаний в биомолекулярных системах». Дж. Хим. Теория вычислений . 2 (3): 873–8. дои : 10.1021/ct050247s . ПМИД 26626694 .
- ^ Тирион, ММ (1996). «Упругие движения большой амплитуды в белках на основе однопараметрического атомного анализа». Физ. Преподобный Летт . 77 (9): 1905–1908. Бибкод : 1996PhRvL..77.1905T . doi : 10.1103/physrevlett.77.1905 . ПМИД 10063201 .
- ^ Ли, Х., Чанг, Ю.Ю., Ян, Л.В., и Бахар, И. (2016). iGNM 2.0: база данных моделей гауссовой сети для биомолекулярной структурной динамики. Исследования нуклеиновых кислот, 44(D1), D415-D422.
- ^ Атилган, Арканзас; Даррелл, СР; Джерниган, РЛ; Демирель, MC; Кескин, О.; Бахар, И. (2001). «Анизотропия флуктуационной динамики белков с эластичной сетевой моделью» . Биофиз. Дж . 80 (1): 505–515. Бибкод : 2001BpJ....80..505A . дои : 10.1016/s0006-3495(01)76033-x . ПМК 1301252 . ПМИД 11159421 .
- ^ Бакан, А.; Мейрелеш, LM; Бахар, И. (2011). «ProDy: динамика белков, выведенная из теории и экспериментов» . Биоинформатика . 27 (11): 1575–1577. doi : 10.1093/биоинформатика/btr168 . ПМК 3102222 . ПМИД 21471012 .
- ^ Бакан, А.; Дутта, А.; Мао, В.; Лю, Ю.; Ченнубхотла, К.; Лезон, ТР; Бахар, И. (2014). «Evol и ProDy для объединения эволюции белковых последовательностей и структурной динамики» . Биоинформатика . 30 (18): 2681–2683. doi : 10.1093/биоинформатика/btu336 . ПМЦ 4155247 . ПМИД 24849577 .