Ячейка сетки

Ячейка сетки — это тип нейрона в энторинальной коре , который срабатывает через регулярные промежутки времени, когда животное перемещается по открытой местности, что позволяет ему понять свое положение в пространстве, сохраняя и интегрируя информацию о местоположении, расстоянии и направлении. [1] Решетчатые клетки были обнаружены у многих животных, в том числе у крыс . [1] мыши , [2] летучие мыши , [3] обезьяны , [4] и люди. [5] [6]
Клетки сетки были открыты в 2005 году Эдвардом Мозером , Мэй-Бритт Мозер и их студентами Торкелем Хафтингом, Марианной Фин и Стурлой Молден в Центре биологии памяти (CBM) в Норвегии. Они были удостоены Нобелевской премии по физиологии и медицине 2014 года вместе с Джоном О'Кифом за открытие клеток, составляющих систему позиционирования в мозге. Расположение пространственных полей возбуждения, расположенных на равных расстояниях от соседей, привело к гипотезе, что эти клетки кодируют нейронное представление евклидова пространства . [1] Открытие также предложило механизм динамического расчета собственного положения на основе постоянно обновляемой информации о положении и направлении.

Чтобы обнаружить активность клеток сетки в типичном эксперименте на крысах, электрод имплантируют в дорсомедиальную энторинальную кору , который может регистрировать активность отдельных нейронов , и он собирает записи, когда крыса свободно передвигается на открытой арене. Полученные данные можно визуализировать, отмечая положение крысы на карте арены каждый раз, когда нейрон запускает потенциал действия . Эти отметки со временем накапливаются, образуя набор небольших кластеров, которые, в свою очередь, образуют вершины сетки равносторонних треугольников. Правильный треугольник отличает ячейки сетки от других типов ячеек, демонстрирующих пространственную активацию. Напротив, если таким же образом исследовать клетку места из гиппокампа крысы, то в данной среде метки часто образуют только один кластер (одно «поле места»), и даже когда наблюдаются несколько кластеров, нет никакой достоверной информации. заметна закономерность в их расположении.
Предыстория открытия
[ редактировать ]В 1971 году Джон О'Киф и Джонатон Достровский сообщили об открытии клеток места крысы в гиппокампе — клеток, которые запускают потенциалы действия, когда животное проходит через определенную небольшую область пространства, которая называется полем места клетки. [7] Это открытие, поначалу вызывавшее споры, привело к серии исследований, кульминацией которых стала публикация в 1978 году книги О'Кифа и его коллеги Линн Надель под названием «Гиппокамп как когнитивная карта» (фраза, которая также фигурировала в названии книги). статья 1971 года) [8] — в книге утверждается, что нейронная сеть гиппокампа создает экземпляры когнитивных карт, как предположил психолог Эдвард К. Толман . Эта теория вызвала большой интерес и послужила мотивом для сотен экспериментальных исследований, направленных на выяснение роли гиппокампа в пространственной памяти и пространственной навигации.
Поскольку энторинальная кора обеспечивает наибольший входной сигнал в гиппокамп, было очевидно важно понять свойства пространственной активации энторинальных нейронов. Самые ранние исследования, такие как Quirk et al. (1992) описали нейроны энторинальной коры как имеющие относительно большие и нечеткие поля мест. [9] Но Мозеры полагали, что можно было бы получить иной результат, если бы записи были сделаны из другой части энторинальной коры. Энторинальная кора представляет собой полоску ткани, идущую вдоль заднего края мозга крысы от вентральной к дорсальной стороне. Анатомические исследования показали, что разные секторы энторинальной коры проецируются на разные уровни гиппокампа: дорсальный конец ЭК выступает в дорсальный гиппокамп, вентральный конец - в вентральный гиппокамп. [10] Это было актуально, поскольку несколько исследований показали, что клетки места в дорсальном гиппокампе имеют значительно более четкие поля мест, чем клетки более вентральных уровней. [11] Но в каждом исследовании энторинальной пространственной активности до 2004 г. использовались электроды, имплантированные вблизи вентрального конца ЭК. Соответственно, вместе с Марианной Фин, Стурлой Молден и Менно Виттером Мозеры приступили к изучению пространственной активации на различных дорсально-вентральных уровнях энторинальной коры. Они обнаружили, что в дорсальной части медиальной энторинальной коры (MEC) клетки имели четко очерченные поля мест, как в гиппокампе, но клетки активировались в нескольких местах. [12] В расположении обстреливающих полей были намеки на регулярность, но размер окружающей среды был слишком мал, чтобы в этом исследовании можно было увидеть пространственную периодичность.
В следующей серии экспериментов, о которых сообщалось в 2005 году, использовалась более крупная среда, что привело к признанию того, что клетки на самом деле активировались в виде шестиугольной сетки. [1] Исследование показало, что клетки на одинаковых дорсально-вентральных уровнях MEC имели одинаковый интервал сетки и ориентацию сетки, но что фаза сетки (смещение вершин сетки относительно осей x и y), по-видимому, была случайным образом распределена между клетки. Паттерн периодической стрельбы выражался независимо от конфигурации ориентиров, в темноте, а также при наличии видимых ориентиров и независимо от изменений скорости и направления животного, что позволило авторам предположить, что ячейки сетки выражают зависимость, зависящую от интеграции пути. динамический расчет местоположения животного.
За открытие ячеек сетки Мэй-Бритт Мозер и Эдвард Мозер были удостоены Нобелевской премии по физиологии и медицине в 2014 году вместе с Джоном О'Кифом.
Характеристики
[ редактировать ]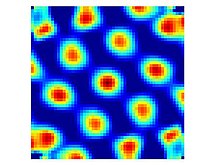
Ячейки сетки — это нейроны, которые активируются, когда свободно движущееся животное пересекает набор небольших областей (полей стрельбы), которые примерно равны по размеру и расположены в виде периодического треугольного массива, охватывающего всю доступную среду. [1] Клетки с этим паттерном активации были обнаружены во всех слоях дорсокаудальной медиальной энторинальной коры (dMEC), но клетки в разных слоях имеют тенденцию различаться в других отношениях. Уровень II содержит наибольшую плотность чистых ячеек сетки в том смысле, что они срабатывают одинаково независимо от направления, в котором животное пересекает ячейку сетки. Ячейки сетки из более глубоких слоев перемешаны с конъюнктивными клетками и клетками направления головы (т.е. в слоях III, V и VI есть клетки с решетчатым рисунком, которые срабатывают только тогда, когда животное смотрит в определенном направлении). [13]
Ячейки сетки, лежащие рядом друг с другом (т. е. ячейки, записанные с одного и того же электрода), обычно имеют одинаковый интервал сетки и ориентацию, но их вершины сетки смещены друг от друга по-видимому случайным смещением. Но ячейки, записанные с отдельных электродов, находящихся на расстоянии друг от друга, обычно имеют разные расстояния между сетками. Клетки, расположенные вентрально (дальше от дорсальной границы MEC), обычно имеют более крупные поля стрельбы в каждой вершине сетки и, соответственно, большее расстояние между вершинами сетки. [1] Общий диапазон шагов сетки точно не установлен: в первоначальном отчете описывался примерно двукратный диапазон шагов сетки (от 39 см до 73 см) в самой дорсальной части (верхние 25%) МЭК, [1] но есть признаки значительно большего масштаба сетки в более вентральных зонах. Брун и др. (2008) зарегистрировали ячейки сетки с нескольких уровней у крыс, бегущих по 18-метровой дорожке, и обнаружили, что интервал сетки увеличился примерно с 25 см в их самых дорсальных участках до примерно 3 м в самых вентральных участках. [14] Эти записи простирались только на 3/4 длины до вентрального кончика, поэтому возможно, что существуют еще более крупные сетки. Было показано, что такие многомасштабные представления теоретически желательны. [15]
Деятельность ячеек сетки не требует визуального ввода, поскольку узоры сетки остаются неизменными, когда все источники света в окружающей среде выключены. [1] Но когда присутствуют визуальные подсказки, они оказывают сильный контроль над выравниванием сеток: вращение карты-подсказки на стенке цилиндра приводит к повороту сеточных узоров на ту же величину. [1] Сетчатые узоры появляются при первом входе животного в новую среду и затем обычно остаются стабильными. [1] Когда животное перемещается в совершенно другую среду, ячейки сетки сохраняют свой интервал сетки, а сетки соседних ячеек сохраняют свои относительные смещения. [1]
Взаимодействие с клетками места гиппокампа
[ редактировать ]Когда крысу перемещают в другую среду, паттерны пространственной активности клеток места гиппокампа обычно демонстрируют «полное переназначение» — то есть паттерн полей места реорганизуется таким образом, что не имеет заметного сходства с паттерном в исходной среде. [16] Однако если характеристики окружающей среды изменяются менее радикально, структура поля места может демонстрировать меньшую степень изменения, называемую «переотображением скорости» , при которой многие клетки меняют свою скорость срабатывания, но большинство ячеек сохраняют поля места в те же места, что и раньше. Это было исследовано с использованием одновременных записей гиппокампа и энторинальных клеток и обнаружено, что в ситуациях, когда гиппокамп демонстрирует переназначение скорости, ячейки сетки демонстрируют неизмененные паттерны активации, тогда как когда гиппокамп демонстрирует полное переназначение, паттерны активации ячеек сетки показывают непредсказуемые сдвиги и вращения. [17]
Тета-ритмичность
[ редактировать ]Нейронная активность почти в каждой части гиппокампальной системы модулируется тета-ритмом гиппокампа , диапазон частот которого у крыс составляет около 6–9 Гц. Энторинальная кора не является исключением: как и гиппокамп, она получает холинергические и ГАМКергические сигналы из медиальной перегородочной области , центрального контроллера теты. Клетки сетки, как и клетки места гиппокампа, демонстрируют сильную тета-модуляцию. [1] Клетки сетки из слоя II MEC также напоминают клетки места гиппокампа тем, что они демонстрируют фазовую прецессию , то есть их пиковая активность продвигается от поздней к ранней фазе тета-цикла, когда животное проходит через вершину сетки. Недавняя модель активности ячеек сетки объяснила эту фазовую прецессию, предположив наличие одномерной сети аттракторов, состоящей из звездчатых ячеек. [18] Большинство ячеек сетки из слоя III не прецессируют, но их пиковая активность в основном ограничивается половиной тета-цикла. Прецессия фазы решетчатых клеток не происходит из гиппокампа, поскольку она продолжает появляться у животных, гиппокамп которых был инактивирован агонистом ГАМК. [19]
Возможные функции
[ редактировать ]Многие виды млекопитающих могут отслеживать пространственное местоположение даже при отсутствии визуальных, слуховых, обонятельных или тактильных сигналов, интегрируя свои движения — способность делать это называется в литературе интеграцией путей . В ряде теоретических моделей исследуются механизмы, с помощью которых нейронные сети могут выполнять интеграцию путей. В большинстве моделей, таких как модели Самсоновича и Макнотона (1997), [20] или Бурак и Фите (2009), [21] Основными ингредиентами являются (1) внутреннее представление положения, (2) внутренние представления скорости и направления движения и (3) механизм смещения закодированного положения на нужную величину при движении животного. Поскольку ячейки в MEC кодируют информацию о положении (ячейки сетки [1] ) и движение (клетки направления головы и конъюнктивные ячейки положения за направлением [13] ), эта область в настоящее время рассматривается как наиболее многообещающий кандидат на место в мозге, где происходит интеграция путей. Однако вопрос остается нерешенным, поскольку у людей энторинальная кора, по-видимому, не требуется для интеграции путей. [22] Бурак и Фите (2009) показали, что компьютерное моделирование системы ячеек сетки способно выполнить интеграцию путей с высоким уровнем точности. [21] Однако более поздние теоретические работы показали, что ячейки сетки могут выполнять более общий процесс шумоподавления, не обязательно связанный с пространственной обработкой. [23]
Хафтинг и др. (2005) [1] предположил, что код места вычисляется в энторинальной коре головного мозга и передается в гиппокамп , который может создавать ассоциации между местом и событиями, необходимые для формирования воспоминаний.

гиппокампа В отличие от ячейки места , ячейка сетки имеет несколько полей активации с регулярными интервалами, которые мозаично формируют окружающую среду в виде шестиугольного узора. Уникальные свойства ячеек сетки заключаются в следующем:
- Ячейки сетки имеют поля стрельбы, рассредоточенные по всей среде (в отличие от полей места, которые ограничены определенными областями среды).
- Поля стрельбы организованы в виде шестиугольной решетки.
- Поля обстрела обычно расположены на одинаковом расстоянии друг от друга, так что расстояние от одного поля обстрела до всех шести соседних полей обстрела примерно одинаковое (хотя при изменении размера среды расстояние между полями может сжиматься или расширяться по-разному в разных направлениях; Барри и др. 2007)
- Поля обстрела расположены одинаково, так что шесть соседних полей расположены с шагом примерно 60 градусов.
Ячейки сетки привязаны к внешним ориентирам, но сохраняются в темноте, что позволяет предположить, что ячейки сетки могут быть частью карты пространственной среды, основанной на самодвижении.
См. также
[ редактировать ]- Пограничная ячейка , открытая в 2008 году.
- Список различных типов клеток в организме взрослого человека
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н Хафтинг, Т.; Фин, М.; Молден, С.; Мозер, МБ; Мозер, Э.И. (2005). «Микроструктура пространственной карты энторинальной коры». Природа . 436 (7052): 801–806. Бибкод : 2005Natur.436..801H . дои : 10.1038/nature03721 . ПМИД 15965463 . S2CID 4405184 .
- ^ Фин, М.; Хафтинг, Т.; Виттер, член парламента; Мозер, Э.И.; Мозер, МБ (2008). «Сетчатые клетки у мышей» . Гиппокамп . 18 (12): 1230–1238. дои : 10.1002/hipo.20472 . ПМИД 18683845 . S2CID 25196865 .
- ^ Ярцев М.М.; Виттер, член парламента; Улановский, Н. (2011). «Ячейки сетки без тета-колебаний в энторинальной коре летучих мышей». Природа . 479 (7371): 103–107. Бибкод : 2011Natur.479..103Y . дои : 10.1038/nature10583 . ПМИД 22051680 . S2CID 1978069 .
- ^ Киллиан, Нью-Джерси; Ютрас, MJ; Буффало, Э.А. (2012). «Карта зрительного пространства в энторинальной коре приматов» . Природа . 491 (7426): 761–4. Бибкод : 2012Natur.491..761K . дои : 10.1038/nature11587 . ПМЦ 3565234 . ПМИД 23103863 .
- ^ Джейкобс, Дж.; Вайдеманн, Коннектикут; Миллер, Дж. Ф.; Солуэй, А.; Берк, Дж. Ф.; Вэй, XX; Сутана, Н.; Сперлинг, MR; Шаран, AD; Фрид, И.; Кахана, MJ (2013). «Прямые записи решетчатой активности нейронов в пространственной навигации человека» . Природная неврология . 16 (9): 1188–90. дои : 10.1038/nn.3466 . ПМЦ 3767317 . ПМИД 23912946 .
- ^ Доллер, CF; Барри, К.; Берджесс, Н. (2010). «Доказательства наличия ячеек сетки в сети человеческой памяти» . Природа . 463 (7281): 657–661. Бибкод : 2010Natur.463..657D . дои : 10.1038/nature08704 . ПМЦ 3173857 . ПМИД 20090680 .
- ^ О'Киф, диджей (1971). «Гиппокамп как пространственная карта. Предварительные данные об активности единиц у свободно передвигающейся крысы». Исследования мозга . 34 (1): 171–175. дои : 10.1016/0006-8993(71)90358-1 . ПМИД 5124915 .
- ^ О'Киф, Дж.; Надель, Л. (1978). Гиппокамп как когнитивная карта . Издательство Оксфордского университета. Архивировано из оригинала 24 марта 2011 г. Проверено 5 ноября 2009 г.
- ^ Квирк, MR (1992). «Позиционные стреляющие свойства медиальных энторинальных нейронов: описание и сравнение с клетками места гиппокампа» . Журнал неврологии . 12 (5): 1945–1963. doi : 10.1523/JNEUROSCI.12-05-01945.1992 . ПМК 6575876 . ПМИД 1578279 .
- ^ Мозер М.Б., Мозер Э.И. (1998). «Функциональная дифференциация в гиппокампе». Гиппокамп . 8 (6): 608–19. doi : 10.1002/(SICI)1098-1063(1998)8:6<608::AID-HIPO3>3.0.CO;2-7 . ПМИД 9882018 . S2CID 32384692 .
- ^ Маурер, АП; Ванродс, СР; Сазерленд, Греция; Липа, П.; Макнотон, Б.Л. (2005). «Самодвижение и происхождение дифференциального пространственного масштабирования вдоль септо-височной оси гиппокампа». Гиппокамп . 15 (7): 841–852. дои : 10.1002/hipo.20114 . ПМИД 16145692 . S2CID 7237113 .
- ^ Фин, М.; Молден, С.; Виттер, член парламента; Мозер, Э.И.; Мозер, М.-Б. (2004). «Пространственное представление в энторинальной коре». Наука . 305 (5688): 1258–1264. Бибкод : 2004Sci...305.1258F . дои : 10.1126/science.1099901 . ПМИД 15333832 .
- ^ Jump up to: а б Сарголини, Ф.; Фин, М.; Хафтинг, Т.; Макнотон, БЛ; Виттер, член парламента; Мозер, М.-Б.; Мозер, Э.И. (2006). «Конъюнктивное представление положения, направления и скорости в энторинальной коре». Наука . 312 (5774): 758–762. Бибкод : 2006Sci...312..758S . дои : 10.1126/science.1125572 . ПМИД 16675704 .
- ^ Брун, В.Х.; Солстад, Т.; Кьельструп, КБ; Фин, М.; Виттер, член парламента; Мозер, Э.И.; Мозер, МБ (2008). «Прогрессивное увеличение масштаба сетки от дорсальной до вентральной медиальной энторинальной коры» . Гиппокамп . 18 (12): 12:00–12:12. дои : 10.1002/hipo.20504 . ПМИД 19021257 .
- ^ Матис, А.; Герц, АВМ; Стеммлер, МБ (2012). «Оптимальные коды населения для пространства: ячейки сетки превосходят ячейки места» . Нейронные вычисления . 24 (9): 2280–2317. дои : 10.1162/NECO_a_00319 . ПМИД 22594833 . S2CID 15755674 .
- ^ Мюллер, РУ; Куби, Дж.Л. (1987). «Влияние изменений окружающей среды на пространственную активацию спайковых клеток гиппокампа» . Дж. Нейроски . 7 (7): 1951–68. doi : 10.1523/JNEUROSCI.07-07-01951.1987 . ПМК 6568940 . ПМИД 3612226 .
- ^ Фин, М.; Хафтинг, Т.; Тревес, А.; Мозер, МБ; Мозер, Э.И. (2007). «Переназначение гиппокампа и перестройка сетки в энторинальной коре». Природа . 446 (7132): 190–194. Бибкод : 2007Natur.446..190F . дои : 10.1038/nature05601 . ПМИД 17322902 . S2CID 4431307 .
- ^ Ковач К.А. (сентябрь 2020 г.). «Эпизодические воспоминания: как гиппокамп и энторинальные кольцевые аттракторы взаимодействуют, создавая их?» . Границы системной нейронауки . 14:68 . дои : 10.3389/fnsys.2020.559186 . ПМЦ 7511719 . ПМИД 33013334 .
- ^ Хафтинг, Т.; Фин, М.; Бонневи, Т.; Мозер, МБ; Мозер, Э.И. (2008). «Независимая от гиппокампа фазовая прецессия в клетках энторинальной решетки». Природа . 453 (7199): 1248–1252. Бибкод : 2008Natur.453.1248H . дои : 10.1038/nature06957 . ПМИД 18480753 . S2CID 597353 .
- ^ Самсонович А, МАБ (1997). «Интеграция путей и когнитивное картографирование в модели нейронной сети с непрерывным аттрактором» . Журнал неврологии . 17 (15): 5900–5920. doi : 10.1523/JNEUROSCI.17-15-05900.1997 . ПМЦ 6573219 . ПМИД 9221787 .
- ^ Jump up to: а б Бурак Ю.; Фите, ИК; Спорнс, О. (2009). Спорнс, Олаф (ред.). «Точная интеграция путей в моделях непрерывных сетевых аттракторов ячеек сетки» . PLOS Вычислительная биология . 5 (2): e1000291. arXiv : 0811.1826 . Бибкод : 2009PLSCB...5E0291B . дои : 10.1371/journal.pcbi.1000291 . ПМЦ 2632741 . ПМИД 19229307 .
- ^ Шрагер, Ю.; Кирван, CB; Сквайр, ЛР (2008). «Нейронная основа когнитивной карты: интеграция пути не требует гиппокампа или энторинальной коры» . Труды Национальной академии наук . 105 (33): 12034–8. Бибкод : 2008PNAS..10512034S . дои : 10.1073/pnas.0805414105 . ПМЦ 2575247 . ПМИД 18687893 .
- ^ Шринивасан, С; Фите, я (2011). «Ячейки сетки генерируют аналоговый код исправления ошибок для исключительно точных нейронных вычислений». Природная неврология . 14 (10): 1330–7. дои : 10.1038/nn.2901 . ПМИД 21909090 . S2CID 17158393 .
Внешние ссылки
[ редактировать ]- Mosers Group. Архивировано 15 апреля 2019 г. в Wayback Machine.
- Видео ячейки сетки