Процесс противника
В этой статье отсутствует информация о деталях восприятия цвета. ( июнь 2022 г. ) |
Процесс противника — это теория цвета человека , которая утверждает, что зрительная система интерпретирует информацию о цвете , обрабатывая сигналы от фоторецепторных клеток антагонистическим образом. Теория противодействующего процесса предполагает, что существует три противостоящих канала , каждый из которых включает противоположную цветовую пару: красный против зеленого , синий против желтого и черный против белого ( яркость ). [ 1 ] Теория была впервые предложена в 1892 году немецким физиологом Эвальдом Герингом .
Теория цвета
[ редактировать ]Дополнительные цвета
[ редактировать ]Если некоторое время смотреть на яркий цвет (например, на красный), а затем отвести взгляд на белое поле, остаточное изображение воспринимается , такое, что исходный цвет будет вызывать его дополнительный цвет (зеленый, в случае красного ввода). Когда дополнительные цвета комбинируются или смешиваются, они «нейтрализуют друг друга» и становятся нейтральными (белыми или серыми). То есть дополнительные цвета никогда не воспринимаются как смесь; не существует «зеленовато-красного» или «желтовато-синего», несмотря на утверждения об обратном . Самый сильный цветовой контраст, который может иметь цвет, — это его дополнительный цвет. Дополнительные цвета также можно назвать «противоположными цветами», и по понятным причинам они составляют основу цветов, используемых в теории противодействующего процесса.
Уникальные оттенки
[ редактировать ]
Цвета, определяющие крайние значения для каждого канала противника, называются уникальными оттенками , в отличие от составных (смешанных) оттенков. Эвальд Геринг первым определил уникальные оттенки как красный, зеленый, синий и желтый и основал их на концепции, согласно которой эти цвета не могут восприниматься одновременно. Например, цвет не может быть одновременно красным и зеленым. [ 2 ] Эти определения были экспериментально уточнены и сегодня представлены средними углами оттенка 353 ° (карминно-красный), 128 ° (кобальтово-зеленый), 228 ° (кобальтово-синий), 58 ° (желтый). [ 3 ]
Уникальные оттенки могут различаться у разных людей и часто используются в психофизических исследованиях для измерения изменений в восприятии цвета из-за недостатков цветового зрения или цветовой адаптации. [ 4 ] Хотя при экспериментальном определении уникальных оттенков существуют значительные межсубъектные различия, [ 3 ] уникальные оттенки человека очень постоянны, с точностью до нескольких нанометров. [ 5 ]
Физиологическая основа
[ редактировать ]Связь с цветовым пространством LMS
[ редактировать ]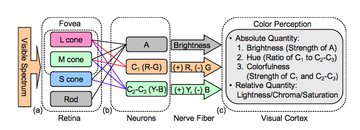
Хотя первоначально считалось, что теории трихроматического и противодействующего процессов противоречат друг другу, позже было показано, что механизмы, ответственные за противостоящий процесс, получают сигналы от трех типов колбочек, предсказанных трихроматической теорией , и обрабатывают их на более сложном уровне. [ 6 ]
имеются три разные колбочки У большинства людей в сетчатке , которые обеспечивают трехцветное цветовое зрение . Цвета определяются пропорциональным возбуждением этих трех типов колбочек, т.е. их квантовым захватом . Уровни возбуждения каждого типа колбочек являются параметрами, определяющими цветовое пространство LMS . процесса противника Чтобы вычислить значения трехстимулного из цветового пространства LMS, необходимо сравнить возбуждения колбочек: [ нужна ссылка ]
- Световой канал противника равен сумме всех трех колбочек (плюс стержневых ячеек ). в некоторых условиях
- Красно-зеленый канал противника равен разнице L- и M-конусов.
- Сине-желтый канал противника равен разности S-конуса и суммы L- и М-конусов.
Неврологическая основа
[ редактировать ]
Считается, что неврологическое преобразование цвета из цветового пространства LMS в противостоящий отросток в основном происходит в латеральном коленчатом ядре (LGN) таламуса , хотя может также происходить в биполярных клетках сетчатки . Ганглиозные клетки сетчатки передают информацию от сетчатки к LGN, который содержит три основных класса слоев: [ 7 ]
- Магноцеллюлярные слои (крупноклеточные) - в основном отвечают за канал яркости.
- Парвоцеллюлярные слои (мелкоклеточные) - в основном ответственны за красно-зеленое противостояние.
- Кониоцеллюлярные слои - в основном ответственны за сине-желтый контраст.
Преимущество
[ редактировать ]Передача информации в цветовом пространстве канала противника имеет преимущество перед передачей ее в цветовом пространстве LMS («необработанные» сигналы от каждого типа конусов). Существует некоторое перекрытие длин волн света , на которые реагируют три типа колбочек (L для длинноволнового света, M для средневолнового света и S для коротковолнового света), поэтому он более эффективен для зрительной системы (от перспектива динамического диапазона ) для записи различий между реакциями колбочек, а не индивидуальной реакции каждого типа колбочек. [ нужна ссылка ] [ сомнительно – обсудить ]
Цветовая слепота
[ редактировать ]Цветовую слепоту можно классифицировать по пораженной колбочке (протан, дейтан, тритан) или по пораженному каналу-противнику ( красно-зеленый или сине-желтый ). В любом случае канал может быть либо неактивным (в случае дихромазии ), либо иметь меньший динамический диапазон (в случае аномальной трихромазии ). Например, люди с дейтеранопией красного и зеленого не видят большой разницы между уникальными оттенками .
История
[ редактировать ]Иоганн Вольфганг фон Гете впервые изучил физиологический эффект противоположных цветов в своей «Теории цветов» в 1810 году. [ 8 ] Гете расположил свой цветовой круг симметрично, «поскольку цвета, диаметрально противоположные друг другу на этой диаграмме, - это те, которые взаимно вызывают друг друга в глазах. Так, желтый требует фиолетового; оранжевый, синий; красный, зеленый; и наоборот: Так снова все промежуточные градации взаимно вызывают друг друга». [ 9 ] [ 10 ]
Эвальд Геринг предложил теорию цвета оппонента в 1892 году. [ 2 ] Он считал, что красный, желтый, зеленый и синий цвета особенные, поскольку любой другой цвет можно описать как их смесь и что они существуют в противоположных парах. То есть воспринимается либо красный, либо зеленый, а не зеленовато-красный: хотя в теории цвета RGB желтый является смесью красного и зеленого, глаз не воспринимает его как таковой.
Новая теория Геринга противоречила преобладающей теории Юнга-Гельмгольца ( трихроматическая теория ), впервые предложенная Томасом Юнгом в 1802 году и развитая Германом фон Гельмгольцем в 1850 году. Эти две теории казались несовместимыми до 1925 года, когда Эрвин Шредингер смог примирить две теории. и показать, что они могут дополнять друг друга. [ 11 ]
Валидация
[ редактировать ]В 1957 году Лео Гурвич и Доротея Джеймсон предоставили психофизическое подтверждение теории Геринга. Их метод назывался «отмена оттенка» . Эксперименты по подавлению оттенка начинаются с цвета (например, желтого) и пытаются определить, сколько цвета противника (например, синего) к одному из компонентов исходного цвета необходимо добавить, чтобы достичь нейтральной точки. [ 12 ] [ 13 ]
В 1959 году Гуннар Светичин и МакНикол [ 14 ] записано из сетчатки рыб и сообщается о трех различных типах клеток:
- Одна клетка реагировала гиперполяризацией на все световые стимулы независимо от длины волны и была названа клеткой светимости.
- Другая клетка ответила гиперполяризацией на коротких волнах и деполяризацией на средних и длинных волнах. Это было названо ячейкой цветности.
- Третья клетка - также клетка цветности - ответила гиперполяризацией на довольно коротких длинах волн с максимумом около 490 нм и деполяризацией на длинах волн более 610 нм.
Светичин и МакНикол назвали клетки цветности желто-синими и красно-зелеными клетками-противниками цвета.
Подобные хроматически или спектрально противоположные клетки, часто включающие пространственную оппозицию (например, красный «включен» в центре и зеленый «выключен» вокруг), были обнаружены в сетчатке позвоночных и латеральном коленчатом ядре (LGN) в 1950-х и 1960-х годах Де Валуа и др. ., [ 15 ] Визель и Хубель, [ 16 ] и другие. [ 17 ] [ 18 ] [ 19 ] [ 20 ]
Следуя примеру Гуннара Светичина , клетки стали называть клетками цвета противника: красно-зелеными и желто-синими. В течение следующих трех десятилетий в сетчатке приматов и LGN продолжали обнаруживаться спектрально противоположные клетки. [ 21 ] [ 22 ] [ 23 ] [ 24 ] В литературе для описания этих клеток используются различные термины, включая хроматически противоположные или -оппоненты, спектрально противоположные или -оппоненты, цвет-противник, цвет-противник, ответ противника и просто клетка-противник.
В других областях
[ редактировать ]Другие применили идею противодействия стимуляции за пределами зрительных систем, описанную в статье о теории противодействующих процессов . В 1967 году Род Григг расширил эту концепцию, чтобы отразить широкий спектр противостоящих процессов в биологических системах. [ 25 ] В 1970 году Соломон и Корбит расширили общую модель неврологического оппонентного процесса Хурвича и Джеймсона, чтобы объяснить эмоции, наркозависимость и мотивацию к работе. [ 26 ] [ 27 ]
Приложения
[ редактировать ]Теорию цвета противника можно применить к компьютерному зрению и реализовать как гауссову цветовую модель. [ 28 ] и модель обработки естественного зрения . [ 29 ] [ 30 ] [ 31 ]
Критика и дополнительные цветовые клетки
[ редактировать ]Существует много споров по поводу того, является ли теория обработки сигналов оппонента лучшим способом объяснения цветового зрения. Было проведено несколько экспериментов со стабилизацией изображения (когда наблюдается потеря границ), которые дали результаты, предполагающие, что участники видели «невозможные» цвета или цветовые комбинации, которые люди не должны видеть в соответствии с теорией обработки противника. Однако многие критикуют, что этот результат может быть всего лишь иллюзорным опытом. Вместо этого критики и исследователи начали пытаться объяснить цветовое зрение, ссылаясь на механизмы сетчатки, а не на обработку оппонента, которая происходит в зрительной коре головного мозга.
По мере накопления записей отдельных клеток многим физиологам и психофизикам стало ясно, что цвета противника не могут удовлетворительным образом объяснить спектрально противоположные реакции отдельных клеток. Например, Джеймсон и Д'Андраде. [ 32 ] проанализировали теорию цветов противника и обнаружили, что уникальные оттенки не соответствуют спектрально противоположным реакциям. сам де Валуа [ 33 ] резюмировал это: «Хотя нас, как и других, больше всего впечатлило обнаружение клеток-противников, в соответствии с предположениями Геринга, когда дух времени в то время был категорически против этой идеи, самые ранние записи выявили несоответствие между клетками Геринга-Гурвича и Каналы восприятия противника Джеймсона и характеристики ответа клеток-противников в латеральном коленчатом ядре макаки». Вальберг [ 34 ] вспоминает, что « среди нейрофизиологов стало обычным использовать цветовые термины, когда речь идет о клетках-противниках, например, в обозначениях «красные клетки» и «зеленые клетки»… В ходе дискуссии… некоторые психофизики были рады увидеть то, что они считали оппозицией, подтвержденной на объективном, физиологическом уровне. Следовательно, не было проявлено никаких колебаний в отношении прямой связи уникальных и полярных цветовых пар с оппозицией колбочек, несмотря на доказательства обратного .... в учебниках до сих пор есть доказательства. , повторил заблуждение Связывание восприятия уникального цвета непосредственно с процессами противника на периферийном колбочке. Аналогия с гипотезой Геринга была развита еще дальше, предполагая, что каждый цвет в паре уникальных цветов противника может быть отождествлен либо с возбуждением, либо с торможением одного и того же типа. клетки-противника » Вебстер и др. [ 35 ] и Вюргер и др. [ 36 ] убедительно подтвердили, что спектрально противоположные реакции отдельных клеток не совпадают с цветами противника уникального оттенка.
Более поздние эксперименты показывают, что взаимосвязь между реакциями отдельных клеток-«цветовых противников» и воспринимаемым цветовым противником даже более сложна, чем предполагалось. Эксперименты Зеки и др., [ 37 ] используя Land Color Mondrian, показали, что когда обычные наблюдатели видят, например, зеленую поверхность, которая является частью разноцветной сцены и отражает больше зеленого, чем красного света, она выглядит зеленой, а ее остаточное изображение — пурпурным. Но когда одна и та же зеленая поверхность отражает больше красного, чем зеленого света, она все равно выглядит зеленой (из-за действия механизмов постоянства цвета), а ее послеизображение по-прежнему воспринимается как пурпурное. Это справедливо и для других цветов, и можно резюмировать это следующим образом: точно так же, как поверхности сохраняют свои цветовые категории, несмотря на широкомасштабные колебания длины волны и энергии отраженного от них света, цвет создаваемого остаточного изображения при просмотре поверхностей также сохраняет свою цветовую категорию и, следовательно, также не зависит от состава длины волны и энергии света, отраженного от просматриваемого участка. Другими словами, существует постоянство цветов последующих изображений. Это еще раз подчеркивает необходимость более глубокого исследования взаимосвязи между реакциями отдельных клеток-противников и перцептивной цветовой оппозицией, с одной стороны, и необходимость лучшего понимания того, генерируют ли физиологические процессы оппонента перцептивные цвета оппонента или же последние являются генерируется после генерации цветов.
В 2013 году Придмор [ 38 ] утверждал, что большинство красно-зеленых клеток, описанных в литературе, на самом деле кодируют красно-голубые цвета. Таким образом, клетки кодируют дополнительные цвета, а не цвета соперника. Придмор также сообщил о зелено-пурпурных клетках сетчатки и V1. Таким образом, он утверждал, что красно-зеленые и сине-желтые клетки следует называть комплементарными клетками «зелено-пурпурный», «красно-голубой» и «сине-желтый». Пример дополнительного процесса можно увидеть, глядя на красный (или зеленый) квадрат в течение сорока секунд, а затем сразу же глядя на белый лист бумаги. Затем наблюдатель видит голубой (или пурпурный) квадрат на чистом листе. Это дополнительное цветовое остаточное изображение легче объяснить с помощью трихроматической теории цвета ( теории Янга-Гельмгольца ), чем традиционной теории цвета RYB; В теории противодействующего процесса усталость путей, продвигающих красный цвет, создает иллюзию голубого квадрата. [ 39 ]
Авторское эссе Конвея , Малика-Мораледы и Гибсона за 2023 год. [ 40 ] заявил, что «проанализировал психологические и физиологические доказательства теории цветов оппонента», и прямо заявил, что «теория ошибочна». [ 40 ]
См. также
[ редактировать ]- Невозможный цвет
- Цветовое пространство CIELAB
- Система натуральных цветов
- Дополнительные цвета
- Остаточное изображение
Ссылки
[ редактировать ]- ^ Майкл Фостер (1891). Учебник физиологии . Леа Бразерс и Ко с. 921 .
- ^ Jump up to: а б Геринг Э., 1964. Очерки теории светочувствия . Кембридж, Массачусетс: Издательство Гарвардского университета.
- ^ Jump up to: а б Мияхара, Э. (2003). «Фокальные цвета и уникальные оттенки» . Перцептивные и моторные навыки . 97 (3_дополнение): 1038–1042. дои : 10.2466/pms.2003.97.3f.1038 . ПМК 1404500 . ПМИД 15002843 .
- ^ Трегиллус, Кэтрин (2019). «Долговременная адаптация к цвету» . Современное мнение в области поведенческих наук . 30 : 116–121. дои : 10.1016/j.cobeha.2019.07.005 . S2CID 201042565 .
- ^ Моллон, JD (1997). «О природе неповторимых красок» . Наследие цветового зрения Джона Далтона : 381–392.
- ^ Кандел Э.Р., Шварц Дж.Х. и Джесселл Т.М., 2000. Принципы нейронауки , 4-е изд., МакГроу-Хилл, Нью-Йорк. стр. 577–580.
- ^ М. Годрати, С.-М. Халиг-Разави, С.Р. Леки, На пути к построению более сложного представления о латеральном коленчатом ядре: последние достижения в понимании его роли, Prog. Нейробиол. 156:214–255, 2017.
- ^ «Теория цвета Гете» . Видение науки и возникновение современного искусства . Архивировано из оригинала 16 сентября 2008 г.
- ^ Гете Дж (1810). Теория цвета, параграф №50 .
- ^ «Гете о цветах» . Арт-Союз . 2 (18): 107. 15 июля 1840 г. Архивировано из оригинала 21 декабря 2017 г.
- ^ Найл, Кейт К. (1988). «О трихроматических теориях и теориях оппонентного процесса: статья Э. Шредингера». Пространственное видение . 3 (2): 79–95. дои : 10.1163/156856888x00050 . ПМИД 3153667 .
- ^ Гурвич Л.М., Джеймсон Д. (ноябрь 1957 г.). «Теория оппозиционного процесса цветового зрения». Психологический обзор . 64, часть 1 (6): 384–404. дои : 10.1037/h0041403 . ПМИД 13505974 . S2CID 27613265 .
- ^ Вулф Дж. М., Клюендер К. Р., Леви Д. М. (2009). Ощущение и восприятие (третье изд.). Sinauer Associates, Inc. Нью-Йорк: ISBN 978-1-60535-875-8 .
- ^ Светечин Г., Макнихол Э.Ф. (ноябрь 1959 г.). «Ретинальные механизмы хроматического и ахроматического зрения». Анналы Нью-Йоркской академии наук . 74 (2): 385–404. Бибкод : 1959NYASA..74..385S . дои : 10.1111/j.1749-6632.1958.tb39560.x . ПМИД 13627867 . S2CID 27130943 .
- ^ Де Валуа Р.Л., Смит С.Дж., Китай С.Т., Кароли А.Дж. (январь 1958 г.). «Реакция одиночных клеток латерального коленчатого ядра обезьяны на монохроматический свет». Наука . 127 (3292): 238–9. Бибкод : 1958Sci...127..238D . дои : 10.1126/science.127.3292.238 . ПМИД 13495504 .
- ^ Визель Т.Н., Хьюбель Д.Х. (ноябрь 1966 г.). «Пространственные и хроматические взаимодействия в латеральном коленчатом теле макаки-резуса». Журнал нейрофизиологии . 29 (6): 1115–56. дои : 10.1152/jn.1966.29.6.1115 . ПМИД 4961644 .
- ^ Вагнер Х.Г., Макнихол Э.Ф., Вольбаршт М.Л. (апрель 1960 г.). «Цветовые реакции противника в ганглиозных клетках сетчатки». Наука . 131 (3409): 1314. Бибкод : 1960Sci...131.1314W . дои : 10.1126/science.131.3409.1314 . ПМИД 17784397 . S2CID 46122073 .
- ^ Нака К.И., Раштон, Вашингтон (август 1966 г.). «S-потенциалы цветовых единиц сетчатки рыб (Cyprinidae)» . Журнал физиологии . 185 (3): 536–55. дои : 10.1113/jphysicalol.1966.sp008001 . ПМЦ 1395833 . ПМИД 5918058 .
- ^ Доу СЗ (ноябрь 1967 г.). «Сетчатка золотой рыбки: организация одновременного цветового контраста». Наука . 158 (3803): 942–4. Бибкод : 1967Sci...158..942D . дои : 10.1126/science.158.3803.942 . ПМИД 6054169 . S2CID 1108881 .
- ^ Бызов А.Л., Трифонов Ю.А. (июль 1968 г.). «Ответ на электрическую стимуляцию горизонтальных клеток сетчатки карпа». Исследование зрения . 8 (7): 817–22. дои : 10.1016/0042-6989(68)90132-6 . ПМИД 5664016 .
- ^ Гурас П., Зреннер Э. (январь 1981 г.). «Цветовое кодирование сетчатки приматов». Исследование зрения . 21 (11): 1591–8. дои : 10.1016/0042-6989(81)90039-0 . ПМИД 7336591 . S2CID 46225236 .
- ^ Деррингтон А.М., Краускопф Дж., Ленни П. (декабрь 1984 г.). «Хроматические механизмы в латеральном коленчатом ядре макаки» . Журнал физиологии . 357 (1): 241–65. дои : 10.1113/jphysicalol.1984.sp015499 . ПМЦ 1193257 . ПМИД 6512691 .
- ^ Рид Р.К., Шепли Р.М. (апрель 1992 г.). «Пространственная структура входов колбочек в рецептивные поля в латеральном коленчатом ядре приматов». Природа . 356 (6371): 716–8. Бибкод : 1992Natur.356..716R . дои : 10.1038/356716a0 . ПМИД 1570016 . S2CID 22357719 .
- ^ Ланкхит М.Дж., Ленни П., Краускопф Дж. (январь 1998 г.). «Отличительные характеристики подклассов красно-зеленых P-клеток в ЛГН макак». Визуальная нейронаука . 15 (1): 37–46. CiteSeerX 10.1.1.553.5684 . дои : 10.1017/s0952523898151027 . ПМИД 9456503 . S2CID 1558413 .
- ^ Григг Э.Р. (1967). Биологическая относительность . Чикаго: Книги Амаранта.
- ^ Соломон Р.Л., Корбит Дж.Д. (апрель 1973 г.). «Теория мотивации противного процесса. II. Сигаретная зависимость». Журнал аномальной психологии . 81 (2): 158–71. дои : 10.1037/h0034534 . ПМИД 4697797 .
- ^ Соломон Р.Л., Корбит Дж.Д. (март 1974 г.). «Теория оппозиционно-процессной мотивации. I. Временная динамика аффекта». Психологический обзор . 81 (2): 119–45. CiteSeerX 10.1.1.468.2548 . дои : 10.1037/h0036128 . ПМИД 4817611 .
- ^ Гезебрук Дж. М., ван ден Бумгаард Р., Смолдерс А. В., Гертс Х. (декабрь 2001 г.). «Цветовая инвариантность». Транзакции IEEE по анализу шаблонов и машинному интеллекту . 23 (12): 1338–1350. дои : 10.1109/34.977559 .
- ^ Баргут Л. (2014). «Визуальный таксометрический подход к сегментации изображений с использованием нечетко-пространственного разреза таксонов позволяет получить контекстуально релевантные регионы». Обработка информации и управление неопределенностью в системах, основанных на знаниях . Международное издательство Спрингер.
- ^ США 2004059754 , Баргут Л., Ли Л., «Система обработки перцептивной информации», опубликовано 25 марта 2004 г.
- ^ Баргут Л. (21 февраля 2014 г.). Зрение: глобальные изменения контекста восприятия. Обработка локального контраста. Обновлено с включением методов компьютерного зрения . Ученая пресса.
- ^ Джеймсон К., Д'Андраде Р.Г. (1997), «На самом деле это не красный, зеленый, желтый, синий: исследование перцепционного цветового пространства», Цветовые категории в мышлении и языке , Cambridge University Press, стр. 295–319, doi : 10.1017/cbo9780511519819.014 , ISBN 9780511519819
- ^ Де Валуа Р.Л., Де Валуа К.К. (май 1993 г.). «Многоступенчатая цветовая модель». Исследование зрения . 33 (8): 1053–65. дои : 10.1016/0042-6989(93)90240-w . ПМИД 8506645 . S2CID 53187961 .
- ^ Вальберг А. (сентябрь 2001 г.). «Исправление к «Уникальные оттенки: старая проблема нового поколения» » . Исследование зрения . 41 (21): 2811. doi : 10.1016/s0042-6989(01)00243-7 . ISSN 0042-6989 . S2CID 1541112 .
- ^ Вебстер М.А., Мияхара Э., Малкок Г., Рейкер В.Е. (сентябрь 2000 г.). «Вариации нормального цветового зрения. II. Уникальные оттенки». Журнал Оптического общества Америки А. 17 (9): 1545–55. Бибкод : 2000JOSAA..17.1545W . дои : 10.1364/josaa.17.001545 . ПМИД 10975364 .
- ^ Вюргер С.М., Аткинсон П., Кроппер С. (ноябрь 2005 г.). «Конус влияет на механизмы уникального оттенка» . Исследование зрения . 45 (25–26): 3210–23. дои : 10.1016/j.visres.2005.06.016 . ПМИД 16087209 . S2CID 5778387 .
- ^ Зеки С., Чидл С., Пеппер Дж., Милонас Д. (2017). «Постоянство цветных послеобразов» . Границы человеческой неврологии . 11 : 229. дои : 10.3389/fnhum.2017.00229 . ПМЦ 5423953 . ПМИД 28539878 .
- ^ Придмор RW (16 октября 2012 г.). «Спектрально противоположные реакции одной клетки: цвета противника или дополнительные цвета?». Журнал оптики . 42 (1): 8–18. дои : 10.1007/s12596-012-0090-0 . ISSN 0972-8821 . S2CID 122835809 .
- ^ Григгс Р.А. (2009). «Ощущение и восприятие» . Психология: краткое введение (2-е изд.). Стоит издательства . п. 92 . ISBN 978-1-4292-0082-0 . OCLC 213815202 .
Цветовая информация обрабатывается на уровне пострецепторных клеток (биполярными, ганглиозными, таламическими и корковыми клетками) в соответствии с теорией оппонентного процесса.
- ^ Jump up to: а б Конвей, Бевил Р .; Малик-Мораледа, Сайма; Гибсон, Эдвард (30 июня 2023 г.). «Появление цвета и конец теории цветов-оппонентов Геринга» . Клетка . 27 (9): 791–804. дои : 10.1016/j.tics.2023.06.003 . PMC 10527909. PMID 37394292 . Проверено 31 августа 2023 г.
Дальнейшее чтение
[ редактировать ]- Баккус С.А. (2007). «Время и вычисления во внутренней схеме сетчатки». Ежегодный обзор физиологии . 69 : 271–90. дои : 10.1146/annurev.physol.69.120205.124451 . ПМИД 17059359 .
- Масланд Р.Х. (август 2001 г.). «Разнообразие нейронов сетчатки». Современное мнение в нейробиологии . 11 (4): 431–6. дои : 10.1016/S0959-4388(00)00230-0 . ПМИД 11502388 . S2CID 42917038 .
- Масланд Р.Х. (сентябрь 2001 г.). «Основной план сетчатки». Природная неврология . 4 (9): 877–86. дои : 10.1038/nn0901-877 . ПМИД 11528418 . S2CID 205429773 .
- Соуден П.Т., Шинс П.Г. (декабрь 2006 г.). «Серфинг каналов в зрительном мозге» (PDF) . Тенденции в когнитивных науках . 10 (12): 538–45. дои : 10.1016/j.tics.2006.10.007 . ПМИД 17071128 . S2CID 6941223 . Архивировано (PDF) из оригинала 19 июля 2018 г. Проверено 24 сентября 2019 г.
- Вассле Х (октябрь 2004 г.). «Параллельная обработка данных в сетчатке млекопитающих». Обзоры природы. Нейронаука . 5 (10): 747–57. дои : 10.1038/nrn1497 . ПМИД 15378035 . S2CID 10518721 .
- Манзотти, Р. (2017). «Модель дополнительных остаточных изображений, основанная на восприятии» . СЕЙДЖ Открыть . 7 (1). дои : 10.1177/2158244016682478 .
- Юртоглу Н (2018). «Исторические исследования: Международный исторический журнал» (PDF) . 10 (7): 241–264. дои : 10.9737/истор.2018.658 . Архивировано (PDF) из оригинала 06 ноября 2021 г. Проверено 03 мая 2021 г.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - Брогаард Б., Гация Д.Э. (2016). «Корковый цвет и когнитивные науки» . Темы когнитивной науки . 9 (1): 135–150. дои : 10.1111/tops.12241 . ПМИД 28000986 . Архивировано из оригинала 07 октября 2022 г. Проверено 9 августа 2022 г.