Синаптонемный комплекс
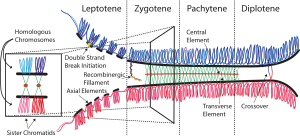

Синаптонемный комплекс ( СК ) представляет собой белковую структуру, которая образуется между гомологичными хромосомами (две пары сестринских хроматид ) во время мейоза и, как полагают, опосредует синапсис и рекомбинацию во время профазы I во время мейоза у эукариот . В настоящее время считается, что SC функционирует преимущественно как каркас, позволяющий взаимодействующим хроматидам завершить свою кроссинговерную деятельность. [ 1 ]
Состав
[ редактировать ]Синаптонемный комплекс представляет собой трехчастную структуру, состоящую из двух параллельных латеральных участков и центрального элемента. Эта «трехчастная структура» наблюдается на стадии пахитены первой профазы мейоза как у самцов, так и у самок во время гаметогенеза . До стадии пахитены, во время лептонемы, начинают формироваться латеральные элементы, которые инициируют и завершают свое спаривание на стадии зиготены. После окончания пахинемы СК обычно разбирается и его уже невозможно идентифицировать. [ 2 ]
У человека охарактеризованы три специфических компонента синаптонемного комплекса: белок SC-1 (SYCP1), белок SC-2 (SYCP2) и белок SC-3 ( SYCP3 ). SYCP1 Ген находится на хромосоме 1p13; ген SYCP2 находится на хромосоме 20q13.33; а ген SYCP3 находится на хромосоме 12q. [ 3 ]
Синаптонемный комплекс был описан Монтроузом Дж. Мозесом в 1956 г. в первичных сперматоцитах раков и Д. Фосеттом в сперматоцитах голубя, кошки и человека. [ 4 ] Как видно с помощью электронного микроскопа, синаптонемный комплекс образован двумя «латеральными элементами», главным образом образованными SYCP3 и, во вторую очередь, SYCP2, «центральным элементом», который содержит по крайней мере два дополнительных белка и аминоконцевую область SYCP1, и «центральная область» расположена между двумя боковыми элементами и содержит «поперечные нити», состоящие в основном из белка SYCP1. [ 3 ]
СК можно увидеть с помощью светового микроскопа с использованием окрашивания серебром или с помощью методов иммунофлуоресценции, которые метят белки SYCP3 или SYCP2.
Сборка и разборка
[ редактировать ]Формирование СК обычно отражает спаривание или « синапсис » гомологичных хромосом и может быть использовано для проверки наличия аномалий спаривания у лиц, несущих хромосомные аномалии как по количеству, так и по структуре хромосом. [ 5 ] Половые хромосомы самцов млекопитающих имеют только «частичный синапсис», поскольку обычно образуют лишь короткий SC в паре XY. SC демонстрирует очень небольшую структурную изменчивость среди эукариотических организмов, несмотря на некоторые существенные различия в белках. У многих организмов СК несет один или несколько «рекомбинационных узелков», связанных с его центральным пространством. Считается, что эти узелки соответствуют зрелым событиям генетической рекомбинации или «кроссоверам». У мышей-самцов гамма-облучение увеличивает мейотические кроссинговеры в СК. Это указывает на то, что экзогенно вызванные повреждения ДНК , вероятно, репарируются путем кроссоверной рекомбинации в СК. [ 3 ] Обнаружение взаимодействия между структурным компонентом СК [белком синаптонемного центрального элемента 2 (SYCE2)] и рекомбинационной репарации белком RAD51 также предполагает роль СК в репарации ДНК.
В развитии клеток синаптонемный комплекс исчезает в поздней профазе мейоза I. Он образуется во время зиготены.
Необходимость у эукариот
[ редактировать ]Сейчас очевидно, что синаптонемный комплекс не требуется для генетической рекомбинации у некоторых организмов. Например, у простейших инфузорий, таких как Tetrahymena thermophila и Paramecium Tetraurelia, генетический кроссинговер , по-видимому, не требует образования синаптонемного комплекса. [ 6 ] [ 7 ] Исследования показали, что не только СК образуются в результате генетической рекомбинации, но и мутантные дрожжевые клетки, неспособные собрать синаптонемный комплекс, все еще могут участвовать в обмене генетической информацией. Однако у других организмов, таких как нематода C. elegans , образование хиазм требует формирования синаптонемного комплекса.
Ссылки
[ редактировать ]- ^ Пейдж С.Л., Хоули Р.С. (8 октября 2004 г.). «Генетика и молекулярная биология синаптонемного комплекса». Ежегодный обзор клеточной биологии и биологии развития . 20 (1): 525–58. doi : 10.1146/annurev.cellbio.19.111301.155141 . ПМИД 15473851 .
- ^ Ян Ф., Ван П.Дж. (2009). «Синаптонемный комплекс млекопитающих: каркас и за его пределами». Геномная динамика . 5 : 69–80. дои : 10.1159/000166620 . ISBN 978-3-8055-8967-3 . ПМИД 18948708 .
- ^ Jump up to: а б с Болкун-Филас Э., Холл Е., Спид Р., Таггарт М., Грей С., де Масси Б. и др. (февраль 2009 г.). «Мутация мышиного гена Syce1 нарушает синапсис и предполагает связь между структурными компонентами синаптонемного комплекса и репарацией ДНК» . ПЛОС Генетика . 5 (2): e1000393. дои : 10.1371/journal.pgen.1000393 . ПМК 2640461 . ПМИД 19247432 .
- ^ Моисей, Монтроуз Дж. (1 декабря 1968 г.). «Синаптинемный комплекс». Ежегодный обзор генетики . 2 (1): 363–412. дои : 10.1146/annurev.ge.02.120168.002051 . ISSN 0066-4197 .
- ^ Циклер Д., Клекнер Н. (1 декабря 1999 г.). «Мейотические хромосомы: интегрирующая структура и функция». Ежегодный обзор генетики . 33 (1): 603–754. дои : 10.1146/annurev.genet.33.1.603 . ПМИД 10690419 .
- ^ Лукашевич А., Ховард-Тилль Р.А., Лойдл Дж. (ноябрь 2013 г.). «Нуклеаза Mus81 и геликаза Sgs1 необходимы для мейотической рекомбинации у простейших, лишенных синаптонемного комплекса» . Исследования нуклеиновых кислот . 41 (20): 9296–309. дои : 10.1093/нар/gkt703 . ПМЦ 3814389 . ПМИД 23935123 .
- ^ Чи Дж., Маэ Ф., Лойдл Дж., Логсдон Дж., Данторн М. (март 2014 г.). «Инвентаризация генов мейоза четырех инфузорий показывает преобладание независимого от синаптонемных комплексов перекрестного пути» . Молекулярная биология и эволюция . 31 (3): 660–72. дои : 10.1093/molbev/mst258 . ПМИД 24336924 .
Внешние ссылки
[ редактировать ]- [1] - Синаптонемный комплекс, полученный с помощью 3D-структурированного освещения, фотография доктора Чунг-Джу Рэйчел Ванг, Калифорнийский университет в Беркли, факультет молекулярной и клеточной биологии, Беркли, Калифорния, США, второе место в конкурсе цифровых изображений Olympus Bioscapes 2009 года. Соревнование.
- [2]
- Кунецова А. и др., Мейоз у мышей без синаптонемного комплекса PLOS ONE (2011)