Млекопитающее
| Млекопитающие Временный диапазон:
| |
|---|---|
 | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Амниота |
| Клада : | Synapsida |
| Клада : | Mammaliaformes |
| Сорт: | Млекопитающие Linnaeus , 1758 |
| Живые подгруппы | |
Млекопитающее латинской (от ) мамы «грудь» [ 1 ] это позвоночного животное класса млекопитающих ( / m ə ˈ m eɪ l i . ə / ). Млекопитающие характеризуются присутствием молочных молочных желез для кормления своих молодых, широкой области неокортексного мозга, меха или волос и трех костей среднего уха . Эти характеристики отличают их от рептилий и птиц , от которых их предки расходились в каменноугольный период более 300 миллионов лет назад. Около 6400 существующих видов млекопитающих были описаны и разделены на 29 заказов .
Самыми крупными порядками млекопитающих по количеству видов являются грызуны , летучих мышей и эйлипотифла (включая ежи , родинки и загар ). Следующие три-это приматы (включая людей , обезьян и лемуров ), копыт с ровным носом (включая свиней , верблюдов и китов ) и плотоядные (включая кошек , собак и тюленей ).
Млекопитающие являются единственными живыми членами Synapsida ; Эта клада вместе с Sauropsida (рептилиями и птицами) представляет собой более крупную кладу амниоты . Ранние синапсиды называются « пеликозаврами ». Более продвинутые терапсиды стали доминирующими во время среднего перми . Млекопитающие происходили из Cynodonts , продвинутой группы терапсидов, во время позднего триаса до ранней юры . Млекопитающие достигли своего современного разнообразия в палеогена и неогена периоды кайнозойской эпохи после вымирания неавийских динозавров , и стали доминирующей группой наземных животных с 66 миллионов лет назад до настоящего времени.
Основной тип телосложения млекопитающих в четыре раза , и большинство млекопитающих используют свои четыре конечности для наземного локомоции ; Но в некоторых конечностях адаптированы для жизни в море , в воздухе , на деревьях , под землей или на двух ногах . Китообразные и сирены потеряли свои задние конечности. Млекопитающие варьируются в размере от 30–40 мм (1,2–1,6 дюйма)) Bate Bate до 30 м (98 футов) голубого кита - возможно, самого большого животного, которое когда -либо жили. Максимальная продолжительность жизни варьируется от двух лет для стропения до 211 лет для болотного кита . Все современные млекопитающие рожают живой молодых, за исключением пяти видов монотримов , которые являются млекопитающими. Наиболее богатая вида группу млекопитающих, инфракла, называемое плацентами , имеет плаценту , которая позволяет кормить плода во время беременности .
Большинство млекопитающих интеллектуальны , некоторые обладают большим мозгом, самосознанием и использованием инструментов . Млекопитающие могут общаться и озвучить несколькими способами, включая производство ультразвука , маркировки ароматов , сигналов тревоги , пения , эхолокации ; и, в случае с людьми, сложный язык . Млекопитающие могут организовать себя в общества деления , гаремы и иерархии , но также могут быть одинокими и территориальными . Большинство млекопитающих полигин , но некоторые могут быть моногамными или полиандрузными .
Одомашнивание многих типов млекопитающих людей сыграло важную роль в неолитической революции и привело к тому, что сельское хозяйство заменило охоту и собрание в качестве основного источника пищи для людей. Это привело к серьезной реструктуризации человеческих обществ от кочевых до сидячего, с большим сотрудничеством между более крупными и более крупными группами и, в конечном итоге, развитии первых цивилизаций . Одомашненные млекопитающие предоставляются и продолжают предоставлять питание для транспорта и сельского хозяйства, а также продукты питания ( мясные и молочные продукты ), мех и кожа . На млекопитающих также охотятся и учатся в спорте, хранятся в качестве домашних животных и рабочих животных различных типов, и используются в качестве модельных организмов в науке. Млекопитающие были изображены в искусстве со времен палеолита и появляются в литературе, кино, мифологии и религии. Снижение количества и вымирание многих млекопитающих в первую очередь обусловлено человеческим браконьерством и разрушением среды обитания , в первую очередь обезлесение .
Классификация
Более 70% видов млекопитающих находятся в приказе Роденции , Чироптера и Eulipotyphla .
Классификация млекопитающих прошла через несколько пересмотров с тех пор, как Карл Линнеус первоначально определил класс и в настоящее время [ Когда? ] , Система классификации не принимается. McKenna & Bell (1997) и Wilson & Reeder (2005) предоставляют полезные недавние сборники. [ 2 ] Симпсон (1945) [ 3 ] Обеспечивает систематику происхождения млекопитающих и отношений, которые преподавались повсеместно до конца 20 -го века. Тем не менее, с 1945 года было постепенно обнаружено большое количество новой и более подробной информации: палеонтологическая запись была перекалибрована, а в промежуточные годы наблюдаются много споров и прогресса в отношении теоретических оснований самой систематизации, отчасти благодаря новой концепции кладистика . Хотя полевые работы и лабораторная работа постепенно устарела классификация Симпсона, она остается самой близкой к официальной классификации млекопитающих, несмотря на его известные проблемы. [ 4 ]
Большинство млекопитающих, в том числе шесть наиболее богатых видами заказов , принадлежат плацентарной группе. Три крупнейших ордена в количестве видов - родантия : мыши , крысы , дикобраны , бобры , капибары и другие грызущие млекопитающие; Chiroptera : летучие мыши; и Soricomorpha : летители , родинки и солинодоны . Следующими тремя крупнейшими порядками, в зависимости от биологической классификации используемой схемы , являются приматы : обезьяны , обезьяны и лемуры ; Cetartiodactyla ; : киты и неприятные копылы и плотояд , в которую входят кошки , собаки , ласки , медведи , печати и союзники. [ 5 ] Согласно видам млекопитающих мира , в 2006 году было идентифицировано 5416 видов. Они были сгруппированы в 1229 родов , 153 семейства и 29 заказов. [ 5 ] В 2008 году Международный союз сохранения природы (МСОП) завершил пятилетнюю глобальную оценку млекопитающих для своего красного списка МСОП , который подсчитывал 5488 видов. [ 6 ] Согласно исследованию, опубликованному в журнале «Мамминогии» в 2018 году, число признанных видов млекопитающих составляет 6 495, в том числе 96 недавно вымерших. [ 7 ]
Определения
Слово « млекопитающее » является современным, от научной имени млекопитающих, придуманных Карлом Линнеусом в 1758 году, полученной от латинской мамы («Соска, пап»). В влиятельной статье 1988 года Тимоти Роу определил млекопитающие филогенетически как группу короны млекопитающих, клада , состоящая из самого последнего общего предка живых монотримов ( эхидны и утконоемы ) и терианских млекопитающих ( изрядовые и плаценты ) и все потомки этого предка. [ 8 ] Поскольку этот предок жил в юрский период, определение Роу исключает всех животных из предыдущего триаса , несмотря на то, что триасовые окаменелости в Арамийде были переданы млекопитающим с середины 19-го века. [ 9 ] Если млекопитающие считаются группой короны, ее происхождение может быть примерно как первое известное появление животных, более тесно связанных с некоторыми существующими млекопитающими, чем с другими. Амбондро более тесно связан с монотримами, чем с мирианскими млекопитающими, в то время как амфилеса и амфитерум более тесно связаны с терицами; Поскольку ископаемые из всех трех родов датировались около 167 миллионов лет назад в средней юре , это разумная оценка для появления Короне группы. [ 10 ]
TS KEMP предоставил более традиционное определение: « Синапсиды , которые обладают челюсти зрения между верхними и нижними молярами с поперечным компонентом к движению» или, эквивалентно, с точки зубной артикуляцией и окклюзии Кемпа, клада, происходящая с последним общим предком Синоконодон и живые млекопитающие. [ 11 ] Самым ранним известным синапсид, удовлетворяющим определения Кемпа, это тиктерейум , от 225 млн. Лет , поэтому появление млекопитающих в этом более широком смысле может быть назначена в эту позднюю триасовую дату. [ 12 ] [ 13 ] Тем не менее, это животное, возможно, фактически развивалось во время неогена. [ 14 ]
Молекулярная классификация плацентов

По состоянию на начало 21 -го века молекулярные исследования, основанные на анализе ДНК , предположили новые отношения между семьями млекопитающих. Большинство из этих результатов были независимо подтверждены ретротранспозона данными о присутствии/отсутствии . [ 16 ] Системы классификации, основанные на молекулярных исследованиях, выявляют три основные группы или линии плацентарных млекопитающих - афротерия , ксенартра и болооэутерия , которые расходились в меловом языке . Отношения между этими тремя линиями спорны, и были предложены все три возможные гипотезы, в отношении которой группа является базальной . Этими гипотезами являются Atlantogenata (базальная бороэврия), эпитрация (базальная ксенартра) и exafroplacentalia (базальная афротерия). [ 17 ] Бореоэутерия, в свою очередь, содержит две основные линии - Eaurchontoglires и Laurasiatheria .
Оценки времени дивергенции между этими тремя плацентарными группами варьируются от 105 до 120 миллионов лет назад, в зависимости от типа используемой ДНК (например, ядерной или митохондриальной ) [ 18 ] и различные интерпретации палеогеографических данных. [ 17 ]
| Tarver et al. 2016 [ 19 ] | Сандра Альварес-Карреро и соавт. 2022 [ 20 ] [ 21 ] | |||
|---|---|---|---|---|
|

Эволюция
Происхождение
Synapsida , клада, которая содержит млекопитающих и их вымершие родственники, возникла во время Пенсильванского подпериода (~ 323 миллиона до ~ 300 миллионов лет назад), когда они отделяются от линии рептилий. Млекопитающие Crown Group эволюционировали из более ранних млекопитающих во время ранней юры . Кладограмма принимает млекопитающую, чтобы быть группой короны. [ 22 ]
| Mammaliaformes |
| ||||||||||||||||||||||||||||||||||||||||||
Эволюция от старых амниотов

Первыми полностью наземными позвоночными были амниоты . Как и их амфибийные ранние предшественники в тетраподе , у них были легкие и конечности. Амниотические яйца, однако, имеют внутренние мембраны, которые позволяют развивающемуся эмбриону дышать, но сохраняют воду. Следовательно, амниоты могут откладывать яйца на сухую землю, в то время как амфибии обычно необходимо откладывать яйца в воду.
Первые амниоты, по -видимому, возникли в Пенсильванском подпериоде каменноугольной . Они произошли от более ранних рептилиоморфных амфибийных тетрапод, [ 23 ] который жил на земле, которая уже была заселена насекомыми и другими беспозвоночными, а также папоротниками , мхами и другими растениями. В течение нескольких миллионов лет две важные амниотные линии стали различными: синапсиды , которые впоследствии включают общего предка млекопитающих; и сауропсиды , которые теперь включают черепах , ящериц , змей , крокодилов и динозавров (включая птиц ). [ 24 ] Синапсиды имеют одно отверстие ( височный фенестра ) низкий на каждой стороне черепа. Примитивные синапсиды включали самых больших и жестоких животных раннего пермца , таких как Dimetrodon . [ 25 ] Не меммерские синапсиды были традиционно-и неправильно-называемыми «рептилиями, похожими на млекопитающие» или пеликозаврами ; Теперь мы знаем, что они не были ни рептилиями, ни частью линии рептилий. [ 26 ] [ 27 ]
Therapsids , группа синапсидов, развивалась в среднем пермском , около 265 миллионов лет назад и стала доминирующими земельными позвоночными. [ 26 ] Они отличаются от базальных эупеликозавров в нескольких чертах черепа и челюстей, в том числе: более крупные черепа и резцы , которые равны по размеру в терапидах, но не для эупеликозавров. [ 26 ] Терапсидная линия, приводящая к млекопитающим, прошла через серию этапов, начиная с животных, которые были очень похожи на их ранние предки синапсид и заканчивались вероятностными цинодонтами , некоторые из которых могут легко ошибаться за млекопитающих. Эти этапы характеризовались: [ 28 ]
- Постепенное развитие костного вторичного неба .
- Резкое приобретение эндотермии среди млекопитающих , таким образом, до происхождения млекопитающих на 30–50 миллионов лет [ 29 ] .
- Прогресс в направлении прямой осанки конечности, которая увеличит выносливость животных, избегая ограничения переноса . Но этот процесс был медленным и неустойчивым: например, все травоядные не маммерные терапиды сохраняли растягивающиеся конечности (некоторые поздние формы могли иметь семиректные задние конечности); Пермские плотоядные терапиды имели растянутые передние конечности, а некоторые поздние пермцы также имели полузащитные задние конечности. На самом деле, у современных монотримов все еще есть полузащитные конечности.
- Значения триасом постепенно стали основной костью нижней челюсти, которая, по рептилий).
Первые млекопитающие
Событие перми -триасасического вымирания около 252 миллионов лет назад, которое было длительным событием из -за накопления нескольких импульсов вымирания, закончило доминирование плотоядных терапсидов. [ 30 ] В раннем триасе большинство ниш плотоядных животных средних и крупных сухопутных животных были захвачены архозаврами [ 31 ] который в течение длительного периода (35 миллионов лет) включал крокодиломорфы , [ 32 ] птерозавры ; и динозавры [ 33 ] Тем не менее, крупные цинодонты, такие как Trucidocynodon и Traversodontids, все еще занимали крупные плотоядные и травоядные ниши соответственно. Специализом, динозавры стали доминировать в больших наземных травоядных нишах. [ 34 ]
Первые млекопитающие (в смысле Кемпа) появились в поздней триасовой эпохе (около 225 миллионов лет назад), через 40 миллионов лет после первых терапидов. Они расширились из своей ночной ниши насекомоядного с середины юридического и дающего; [ 35 ] Например, юрская касторокауда была близким родственником истинных млекопитающих, которые имели адаптацию для плавания, копания и ловли рыбы. [ 36 ] Считается, что большинство, если не все, оставались ночными ( ночная узкая часть ), что составляет большую часть типичных признаков млекопитающих. [ 37 ] Большинство видов млекопитающих, которые существовали в мезозойскую эпоху, были многотуркуляциями, эвриконодонты и спалакотерииды . [ 38 ] Самым ранним известным метатетрианом является Sinodelphys , найденная в 125-миллионном раннем мелах -сланцу Китая в северо- в северо-восточной провинции восточной провинции . Окаменевает почти завершено и включает в себя пучки меха и отпечатки мягких тканей. [ 39 ]

Самым старым известным ископаемым среди эвтерии («истинных зверей») является небольшая застрявшая юриста, похожая на Юрию Синенсис , или «мать юрского периода из Китая», датированная 160 миллионов лет назад в поздней юрской игре. [ 40 ] Более поздний эврианский родственник, Эомая , датируемый 125 миллионами лет назад в раннем мелах, обладал некоторыми общими признаками с сумчатными, но не с плацентами, свидетельствуют о том, что эти признаки присутствовали в последнем общем предке двух групп, но позже были позже Потерянный в линии плаценты. [ 41 ] В частности, эпипубические кости простираются вперед от таза. Они не встречаются ни в одной современной плацентарной, но они встречаются в сумчатых, монотримах, других нетерианских млекопитающих и Ukhaetherium , раннемеязном животном в эврианском орденом Asioryctitheria . Это также относится к многотуркуляциям. [ 42 ] Это, по -видимому, является наследственной особенностью, которая впоследствии исчезла в линии плаценты. Эти эпипубические кости, по -видимому, функционируют, усиливая мышцы во время локомоции, уменьшая количество представленного пространства, которое необходимо для содержания плода в течение периодов беременности. Узкая розетка таза указывает на то, что молодые были очень маленькими при рождении, и поэтому беременность была короткой, как и в современных сумчатых. Это говорит о том, что плацента была более поздней разработкой. [ 43 ]
Одним из самых ранних известных монотримов был Teinolophos , который жил около 120 миллионов лет назад в Австралии. [ 44 ] Монотримы имеют некоторые особенности, которые могут быть унаследованы от исходных амниотов, таких как одно и то же отверстие для мочеиспускания, испражнения и воспроизведения ( CLOACA ) - как ящерицы и птицы также делают - [ 45 ] И они откладывают яйца , которые кожистые и необладанные. [ 46 ]
Самые ранние появления функций
Hadrocodium , чьи окаменелости датируются примерно 195 миллионами лет назад, в ранней юре , предоставляет первое четкое свидетельство того, что челюстная сустав, сформированная исключительно сквамозальными и зубными костями; В челюсти нет места для суставной кости, вовлеченной в челюсти всех ранних синапсидов. [ 47 ]

Самое раннее четкое свидетельство о волосах или мехе-окаменелости Касторокауда и Мегакона , с 164 миллионов лет назад в середине юридического возраста. В 1950 -х годах было высказано предположение, что отверстия (отрывки) в верхних челюстях и преднасилах (кости в передней части челюсти) цинодонтов были каналами, которые содержали кровеносные сосуды и нервы ( усы ), как и доказательства волос или волосы шерсть; [ 48 ] [ 49 ] Однако вскоре было отмечено, что на форамин не обязательно показывает, что у животного было вибриссы, так как у современной ящерицы Тупинамбис есть отверстие, которое почти идентична тем, которые встречаются в неммальянском Cynodont thrinaxodon . [ 27 ] [ 50 ] Тем не менее, популярные источники продолжают приписывать усы с Тринаксодоном . [ 51 ] Исследования на пермских копролитах показывают, что не млекопитающие синапсиды эпохи уже имели мех, что устанавливает эволюцию волос, возможно, еще в отношении дицинодонтов . [ 52 ]
Когда Эндотермия впервые появилась в эволюции млекопитающих, неясна, хотя, как правило, она следовала, что впервые развивалась в терапидах, не являющихся млекопитающими . [ 52 ] [ 53 ] Современные монотримы имеют более низкую температуру тела и более изменяющие скорости метаболизма, чем сумчатые и плаценты, [ 54 ] Но есть доказательства того, что у некоторых из их предков, возможно, включающих предков терицев, могли быть температуры тела, как у современных терицев. [ 55 ] Аналогично, некоторые современные Therians, такие как Afrotheres и Xenarthrans, вторично развили температуру нижней части тела. [ 56 ]
Эволюция прямых конечностей у млекопитающих является неполной - живые и ископаемые монотримы имеют растянутые конечности. Парасагиттальная (беспробранная) позиция конечности появилась где -то в позднем юрском или раннемеязном; Это обнаружено в эврианской эомаии и метатринской Sinodelphys , которые датировались 125 миллионами лет назад. [ 57 ] Эпипубические кости, особенность, которая сильно повлияла на воспроизводство большинства клад млекопитающих, впервые обнаружена в TrityLodontidae , что позволяет предположить, что это синапоморфия между ними и MammaliaFormes . Они вездесущие в непласентационных млекопитающих, хотя мегазостродон и эритротерум, по-видимому, их не хватало. [ 58 ]
Было высказано предположение, что первоначальная функция лактации ( производство молока ) заключалась в том, чтобы сохранить яйца влажными. Большая часть аргумента основана на монотринах, млекопитающих, увлекающихся яйцами. [ 59 ] [ 60 ] У людей из людей молочные железы становятся полностью развитыми во время полового созревания, независимо от беременности. [ 61 ]
Подъем млекопитающих

Млекопитающие Thrian взяли на себя экологические ниши среднего и большого размера в кайнозои , после того, как событие из-за мела-палеогена примерно 66 миллионов лет назад опустошенное экологическое пространство, когда-то заполненное неавийскими динозаврами и другими группами рептилий, а также различными другими Группы млекопитающих, [ 63 ] и перенес экспоненциальное увеличение размера тела ( мегафауна ). [ 64 ] Затем млекопитающие очень быстро диверсифицировались; Как птицы, так и млекопитающие демонстрируют экспоненциальный рост разнообразия. [ 63 ] Например, самая ранняя известная летучая мышь датируется около 50 миллионов лет назад, всего через 16 миллионов лет после исчезновения неавийских динозавров. [ 65 ]
Первоначально молекулярные филогенетические исследования показали, что большинство заказов плаценты расходились от 100 до 85 миллионов лет назад и что современные семьи появились в период с позднего эоцена до миоцена . [ 66 ] Тем не менее, не было обнаружено никаких плацентарных окаменелостей до конца мелового. [ 67 ] Самые ранние бесспорные окаменелости плацентов происходят из раннего палеоцена после исчезновения неавийских динозавров. [ 67 ] (Ученые идентифицировали раннее палеоценовое животное по имени Potungulatum donnae в качестве одного из первых млекопитающих плацентаров, [ 68 ] но с тех пор он был реклассифицирован как непласентарный эврик.) [ 69 ] Перекалибровка уровня генетического и морфологического разнообразия предположила позднее меловое происхождение для плацент и палеоценовое происхождение для большинства современных клад. [ 70 ]
Самым ранним известным предком Приматов является Archicebus Achilles [ 71 ] примерно 55 миллионов лет назад. [ 71 ] Этот крошечный примат весил 20–30 граммов (0,7–1,1 унции) и мог помещаться в человеческую ладонь. [ 71 ]
Анатомия
Отличительные черты
Живые виды млекопитающих могут быть идентифицированы с помощью присутствия потовых желез , в том числе тех, которые специализируются на производстве молока, чтобы питать их молодых. [ 72 ] Однако при классификации окаменелостей необходимо использовать другие особенности, поскольку мягкие железы и многие другие особенности не видны у окаменелостей. [ 73 ]
Многие черты, разделяемые всеми живыми млекопитающими, появились среди самых ранних членов группы:
- Jaw Saint - зубная кость (нижняя челюстная кость, которая несет зубы) и сквомозал (небольшая черепная кость) встречаются, чтобы сформировать сустав. В большинстве гнатостомов , включая ранние терапиды , сустав состоит из суставной (маленькой кости в задней части нижней челюсти) и квадратной (небольшая кость в задней части верхней челюсти). [ 47 ]
- Среднее ухо -у млекопитающих корон-групп звук несет с барабанной перепонки цепью из трех костей, маллеуса , инков и стапов . Ознакомительно, Malleus и Incus получены из суставной и квадратной костей, которые составляли челюстный сустав ранних терапидов. [ 74 ]
- Замена зубов - зубы могут быть заменены один раз ( дифхёдонти ) или (как у зубчатых китов и грызунов -муридов ), совсем не ( монофиодонти ). [ 75 ] Слоны, ламантины и кенгуру постоянно вырабатывают новые зубы на протяжении всей своей жизни ( Polyphyodonty ). [ 76 ]
- Призматическая эмаль - эмалевое покрытие на поверхности зуба состоит из призмов, твердых, похожих на стержня, простирающихся от дентина до поверхности зуба. [ 77 ]
- Затылочные мыщечки - две ручки у основания черепа вписываются в самый верхний позвонок шеи ; У большинства других тетрапод , напротив, есть только одна такая ручка. [ 78 ]
По большей части эти характеристики не присутствовали у триасовых предков млекопитающих. [ 79 ] Почти все млекопитающие обладают эпипубической костью, исключением являются современные плаценты. [ 80 ]
Сексуальный диморфизм

В среднем, мужские млекопитающие больше, чем женщины, а мужчины на 10% больше, чем женщины, у более 45% исследованных видов. с мужчинами Большинство заказов млекопитающих также демонстрируют сексуальное диморфизм , хотя некоторые заказы не показывают никакого смещения или значительно смещены от женщин ( Lagomorpha ). Диморфизм сексуального размера увеличивается с размером тела у млекопитающих ( правило Ренша ), что позволяет предположить, что существует параллельное отборочное давление как на мужчин, так и на женском размере. Диморфизм с мужчинами связан с сексуальным отбором для мужчин через конкуренцию мужского пола для женщин, так как существует положительная корреляция между степенью сексуального отбора, о чем свидетельствуют системы спаривания и степень диморфизма, смещенного в мужчине. Степень сексуального отбора также положительно коррелирует с мужским и женским размером у млекопитающих. Кроме того, давление параллельного отбора на женскую массу идентифицируется в этом возрасте при от груди значительно выше у более полигиновых видов, даже при корректировке массы тела. Кроме того, репродуктивная скорость ниже для более крупных женщин, что указывает на то, что выбор плодовитости выбирает для небольших женщин у млекопитающих. Хотя эти паттерны удерживаются у млекопитающих в целом, существует значительное различие между порядками. [ 81 ]
Биологические системы
У большинства млекопитающих есть семь шейных позвонков (кости в шее). Исключениями являются ламантин и ленивцы с двумя точками , которые имеют шесть, и ленив с тремя толками , в котором девять. [ 82 ] Все мозги млекопитающих обладают неокортексом , областью мозга, уникальной для млекопитающих. [ 83 ] Плацентарный мозг имеет мозолистую корпус , в отличие от монотримов и сумчатых. [ 84 ]




Системы кровообращения
млекопитающего Сердце имеет четыре камеры, две верхние предсердности , приемные камеры и два нижних желудочка , разряжающиеся камеры. [ 85 ] Сердце имеет четыре клапана, которые разделяют его камеры и обеспечивают потоки крови в правильном направлении через сердце (предотвращение обратного потока). После газового обмена в легочных капиллярах (кровеносные сосуды в легких), богатая кислородом крови возвращается в левое предсердие через одну из четырех легочных вен . Кровь течет почти непрерывно обратно в атриум, который действует как принимающая камера, и отсюда через отверстие в левом желудочке. Большая часть крови пассивно впадает в сердце, в то время как как предсердие, так и желудочки расслаблены, но к концу периода релаксации желудочков левый предсердий сжимается, перекачивая кровь в желудочку. Сердце также требует питательных веществ и кислорода, найденных в крови, как и другие мышцы, и поставляется через коронарные артерии . [ 86 ]
Респираторные системы
Легкие млекопитающих губки и соты. Дыхание в основном достигается с помощью диафрагмы , которая делит грудную клетку от брюшной полости, образуя купол выпуклости к грудной клетке. Сокращение диафрагмы выравнивает купол, увеличивая объем полости легких. Воздух попадает через полости рта и носа и путешествует по гортани, трахеи и бронхам и расширяет альвеоли . Расслабление диафрагмы имеет противоположный эффект, уменьшая объем полости легких, вызывая выталкивание воздуха из легких. Во время упражнений брюшная стенка сокращается , увеличивая давление на диафрагму, что вынуждает воздух быстрее и сильнее. Клетка грудной клетки способна расширять и сжимать полость грудной клетки через действие других дыхательных мышц. Следовательно, воздух втягивается или исключает из легких, всегда движется по градиенту давления. [ 87 ] [ 88 ] Этот тип легкого известен как легкое сильфоу из -за его сходства с кузнецом . [ 88 ]
Интенсивные системы
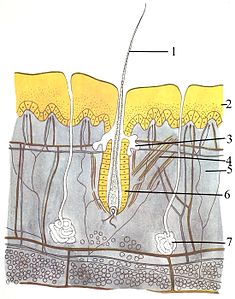
Интенсивная система (кожа) состоит из трех слоев: наиболее внешней эпидермис , дермы и гиподермы . Эпидермис обычно составляет от 10 до 30 клеток; Его основная функция - обеспечить водонепроницаемый слой. Его внешние клетки постоянно теряются; Его клетки Bottommost постоянно делятся и толкают вверх. Средний слой, дерма, в 15-40 раз толщится, чем эпидермис. Дерма состоит из многих компонентов, таких как костные структуры и кровеносные сосуды. Гуподма состоит из жировой ткани , которая хранит липиды и обеспечивает амортизацию и изоляцию. Толщина этого слоя широко варьируется от видов к видам; [ 89 ] : 97 Морские млекопитающие требуют толстой гиподермы ( выпадения ) для изоляции, а правые киты имеют самый толстый взрыв при 20 дюймах (51 см). [ 90 ] Хотя у других животных есть такие особенности, как усы, перья , щетинки или реснички , которые поверхностно напоминают их, ни у животных, кроме млекопитающих . Это окончательная характеристика класса, хотя у некоторых млекопитающих очень мало. [ 89 ] : 61
Пищеварительные системы


Травоядные животные разработали разнообразные физические структуры, чтобы облегчить потребление растительного материала . Чтобы разбить неповрежденные растительные ткани, у млекопитающих развились структуры зубов , которые отражают их предпочтения кормления. Например, у плодоводов (животных, которые питаются в основном фруктами) и травоядные животные, которые питаются мягкой листвой, имеют низкокровированные зубы, специализирующиеся на измельчающей листве и семенах . У пасающих животных, которые имеют тенденцию есть твердые, богатые кремнеземами травы, имеют высокие крови зубы, которые способны измельчать жесткие ткани растений и не изнашиваются так же быстро, как низкокрасированные зубы. [ 91 ] У большинства плотоядных млекопитающих есть плотные зубы (различная длина в зависимости от диеты), длинных клыков и аналогичных схем замены зубов. [ 92 ]
Желудок неглетных колясок (artiodactyla) делится на четыре секции: рубца , ретикулум , омасум и абомасам (только жвачки имеют рубен). После того, как растительный материал употребляется, он смешивается с слюной в рубце и ретикулуме и разделяется на твердый и жидкий материал. Твердые тела объединяются, чтобы сформировать болюс (или CUD ), и отрыгнута. Когда болюс попадает в рот, жидкость выжимается языком и снова глотает. Принимаемая пища переходит в рубцовый и ретикулум, где целлюлолитические микробы ( бактерии , простейшие и грибы ) продуцируют целлюлазу , которая необходима для разрушения целлюлозы в растениях. [ 93 ] Пескодактилы , в отличие от жвачных животных, хранят перевариваемую пищу, которая оставила желудок в увеличенной слепой кишке , где она ферментируется бактериями. [ 94 ] У хищников есть простой желудок, адаптированный к перевариванию в основном мясо, по сравнению с сложными пищеварительными системами травоядных животных, которые необходимы для разрушения прочных сложных растений. Caecum или намного шире , либо отсутствует, либо короткий и простой, а большая кишка не саккулирован чем тонкая кишка. [ 95 ]
Экскреторные и мочеполовые системы


млекопитающих Экскреторная система включает в себя множество компонентов. Как и большинство других наземных животных, млекопитающие являются уреотелитными и превращают аммиак в мочевину , которая выполняется печенью как часть цикла мочевины . [ 96 ] Билирубин , отходный продукт, полученный из клеток крови , пропускается через желчь и мочу с помощью ферментов, выделяемых печени. [ 97 ] Прохождение билирубина через желчь через кишечный тракт придает фекалиям млекопитающих отличительную коричневую окраску. [ 98 ] Отличительные особенности почки млекопитающего включают присутствие почечной таза и почечных пирамид , а также четко различимой коры и мозгового мозга , что связано с наличием удлиненных петель Henle . Только в почках млекопитающих есть форма бобов, хотя есть некоторые исключения, такие как многопользовательские прочитанные почки из пехотинцев, китообразных и медведей. [ 99 ] [ 100 ] У большинства взрослых плацентарных млекопитающих нет оставшихся следов клоаки . В эмбрионе эмбриональная клоака делится на заднюю область, которая становится частью заднего прохода, и передняя область, которая имеет разные судьбы в зависимости от пола человека: у женщин он развивается в вестибюль или урогенитальный пазух , который получает уретру и влагалище , в то время как у мужчин он образует всю уретру полового члена . [ 100 ] [ 101 ] Тем не менее, Tenrecs , Golden Poles и некоторые выстрелы сохраняют клоаку как взрослые. [ 102 ] В сумчатых, генитальные тракты отделены от заднего прохода, но след у исходной клоаки действительно остается внешне. [ 100 ] Монотрим, который переводится с греческого в «одиночную дыру», имеют настоящую клоаку. [ 103 ] Моча течет из мочеточников в клоаку в монотринах и в мочевой пузырь у плацентарных млекопитающих. [ 100 ]
Звуковое производство

Как и во всех других тетраподах, у млекопитающих есть гортани , которая может быстро открываться и близко к изготовлению звуков, и супраларингеальный вокальный тракт , который фильтрует этот звук. Легкие и окружающая мускулатура обеспечивают воздушный поток и давление, необходимое для профолаза . Гортань управляет высотой и объемом звука, но прочность, которую легкие оказывают в выдохе, также способствует громкости. Более примитивные млекопитающие, такие как эхидна, могут только шипять, поскольку звук достигается исключительно путем выдыхания через частично закрытую гортань. Другие млекопитающие протекают с использованием вокальных складок . Движение или напряженность вокальных складок могут привести к многим звукам, таким как мурлыки и крик . Млекопитающие могут изменить положение гортани, позволяя им дышать через нос, проглотив рту и образуя как оральные, так и носовые звуки; Носовые звуки, такие как собачья, как правило, являются мягкими звуками, а устные звуки, такие как кора для собак, как правило, громкие. [ 104 ]
У некоторых млекопитающих есть большая гортани и, следовательно, низкокачественный голос, а именно летучая мышь с головами с молотом ( Hypsignathus monstrosus ), где гортань может занять всю грудную полость , толкая легкие, сердце и трахею в живот . [ 105 ] Большие вокальные прокладки также могут опустить шаг, как в низких реве с большими кошками . [ 106 ] Производство инфразуна возможно у некоторых млекопитающих, таких как африканский слон ( Loxodonta spp.) И Baleen Whales . [ 107 ] [ 108 ] Маленькие млекопитающие с небольшими гортани могут производить ультразвук , что может быть обнаружено путем модификаций в среднем ухе и улитке . Ультразвук не является апелтуальным для птиц и рептилий, что могло быть важно во время мезозойского, когда птицы и рептилии были доминирующими хищниками. Этот частный канал используется некоторыми грызунами, например, в общении с матерью-ядром и летучими мышами при эхолокации. Зубные киты также используют эхолокацию, но, в отличие от вокальной мембраны, которая простирается вверх от вокальных складок, у них есть дыня для манипулирования звуками. Некоторые млекопитающие, а именно приматы, имеют воздушные мешки, прикрепленные к гортани, что может функционировать для снижения резонансов или увеличения объема звука. [ 104 ]
Система вокального производства контролируется ядрами черепного нерва в мозге и поставляется рецидивирующим нервами гортани и верхним гортатным нервом , ветвями блуждающего нерва . Глокальный тракт поставляется гипоглоссальным нервом и лицевыми нервами . Электрическая стимуляция периакедуктальной серой млекопитающих области (ПЭГ) среднего мозга вызывает вокализации. Способность изучать новые вокализации иллюстрируется только людям, тюленями, китообразными, слонами и, возможно, летучими мышами; У людей это является результатом прямой связи между моторной корой , которая контролирует движение, и моторными нейронами в спинном мозге. [ 104 ]
Шерсть

Основной функцией меха млекопитающих является терморегуляция . Другие включают защиту, сенсорные цели, гидроизоляцию и камуфляж. [ 109 ] Различные виды меха служат разным целям: [ 89 ] : 99
- Окончательный - который может быть сброшен после достижения определенной длины
- Vibrissae - сенсорные волосы, чаще всего усы
- Пелаж-охраняйте волосы, недостаточные волосы и волосы
- Шипы - жесткие охранные волосы, используемые для защиты (например, на дикобразах )
- Компания - длинные волосы, обычно используемые в визуальных сигналах. льва (например, грива )
- Велеи - часто называют «пуховым мехом», который изолирует новорожденных млекопитающих
- Шерсть - длинная, мягкая и часто кудрявая
Терморегуляция
Длина волос не является фактором в терморегуляции: например, некоторые тропические млекопитающие, такие как ленивцы, имеют одинаковую длину меха, что и некоторые арктические млекопитающие, но с меньшей изоляцией; И, наоборот, другие тропические млекопитающие с короткими волосами имеют такую же изоляционную ценность, что и арктические млекопитающие. Плотность меха может увеличить изоляционную ценность животного, а у арктических млекопитающих особенно есть плотный мех; Например, у мускусного быка есть охраняющие волосы, размеры 30 см (12 дюймов), а также плотную нижнюю часть, которая образует герметичный слой, что позволяет им выжить при температуре -40 ° C (-40 ° F). [ 89 ] : 162–163 Некоторые млекопитающие в пустыне, такие как верблюды, используют плотный меха, чтобы предотвратить достижение солнечного тепла, позволяя животному оставаться прохладным; Мех верблюда может достигать 70 ° C (158 ° F) летом, но кожа остается при 40 ° C (104 ° F). [ 89 ] : 188 Водные млекопитающие , наоборот, ловят воздух в мехе, чтобы сохранить тепло, сохраняя кожу сухим. [ 89 ] : 162–163

Окраска
Шаты из млекопитающих окрашены по разным причинам, основные селективные давления, включая камуфляж , половой отбор , связь и терморегуляцию. Окраска как в волосах, так и в коже млекопитающих в основном определяется типом и количеством меланина ; Eumelanins для коричневых и черных цветов и феомеланина для диапазона желтоватых до красновато -красновато -цветов, придавая млекопитающим в земле . [ 110 ] [ 111 ] У некоторых млекопитающих есть более яркие цвета; Некоторые обезьяны, такие как мандриллы и обезьяны -вервет , а также опоссумы, такие как мексиканские опоссумы мыши и шерстяные опоссумы Дерби , имеют синюю кожу из -за световой дифракции в коллагеновых волокнах. [ 112 ] Многие ленивцы кажутся зелеными, потому что их мех занимает зеленые водоросли ; Это может быть симбиотическое отношение, которое обеспечивает камуфляж для ленивцев. [ 113 ]
Камуфляж - это мощное влияние на большое количество млекопитающих, поскольку он помогает скрывать людей от хищников или добычи. [ 114 ] У арктических и субарктических млекопитающих, таких как «Арктическая лиса » ( Alopex Lagopus ), воротничный лемминг ( Dicrostonyx Groenlandicus ), Стоат ( Mustela Erminea ) и снегоступа ( Lepus Americanus ), сезонное изменение цвета между коричневыми и белыми зимой движется в режиме зимы Камуфляж. [ 115 ] Некоторые древесные млекопитающие, в частности, приматы и сумчатые, имеют оттенки фиолетовой, зеленой или синей кожи на частях их тел, что указывает на некоторое явное преимущество в их в значительной степени древесной средой обитания из -за сходящейся эволюции . [ 112 ]
Апосематизм , предупреждающий возможных хищников, является наиболее вероятным объяснением черно-белого пелажа многих млекопитающих, которые способны защищать себя, например, в плавно пахнущем скунсе и мощном и агрессивном медовом барсуке . [ 116 ] Цвет пальто иногда является сексуально диморфным , как у многих видов приматов . [ 117 ] Различия в цвете женского и мужского пальто могут указывать на питание и уровень гормонов, что важно при отборе партнеров. [ 118 ] Цвет пальто может влиять на способность сохранять тепло, в зависимости от того, сколько света отражается. Млекопитающие с более темным цветным пальто могут поглощать больше тепла от солнечного излучения и оставаться теплее, а у некоторых меньших млекопитающих, таких как полевые , более темный мех зимой. Белый, без пигментный мех арктических млекопитающих, таких как белый медведь, может отражать больше солнечного излучения непосредственно на кожу. [ 89 ] : 166–167 [ 109 ] Ослепительная черно-белая полоса зебр , похоже, обеспечивает некоторую защиту от кусающих мух. [ 119 ]
Репродуктивная система

Млекопитающие размножаются путем внутреннего оплодотворения [ 120 ] и являются исключительно гонохоричными (животное рождается с мужскими или женскими гениталиями, в отличие от гермафродитов , где такого раскола нет). [ 121 ] Мужские млекопитающие ослабевают женщин во время совокупления и эякулята спермы в сам по себе через половой член , который может быть содержится в приготовлении , когда не будет прямо. Мужские плаценты также мочатся через пенис, а некоторые плаценты также имеют кость полового члена ( Baculum ). [ 122 ] [ 123 ] [ 124 ] Уборщики обычно раздвоились пенисы, [ 125 ] в то время как пенис Echidna обычно имеет четыре головы только с двумя функционирующими. [ 126 ] В зависимости от вида, эрекция может быть вызвана кровотоком в сосудистую, губчатую ткань или мышечное действие. [ 122 ] Яички , большинства млекопитающих спускаются в мошонку которая обычно является позади полового члена, но часто является передней в сумчатых. У женщин -млекопитающих обычно есть вульва ( клитор и губы ) снаружи, в то время как внутренняя система содержит парные яйцеклетки , 1–2 матки , 1–2 шейения и влагалище . [ 127 ] [ 128 ] У сумчатых есть два боковых влагалища и медиальное влагалище. «Влагалище» монотримов лучше понимается как «урогенитальный синус». Системы матки плацентарных млекопитающих могут варьироваться между дуплексом, где есть два матки и шейки, которые открываются во влагалище, двухпартий, где два рога матки имеют единую шейку матки, которая соединяется с влагалищем, двумя Рога, которые подключены дистально, но отдельные медиально создают Y-образную форму, и Simplex, который имеет одну матку. [ 129 ] [ 130 ] [ 89 ] : 220–221, 247

Состояние наследственного репродукции млекопитающих-это рождение относительно неразвитого молодого, либо через прямую вивипарию , либо в короткие сроки в качестве яиц с мягкими оболочниками. Вероятно, это связано с тем, что туловище не может расширяться из -за наличия эпипубических костей . Самая старая демонстрация этого репродуктивного стиля - с Kayentatherium , который производил неразвитые перинаты , но при гораздо более высоких размерах мусора, чем любое современное млекопитающее, 38 образцов. [ 131 ] Большинство современных млекопитающих вивипарусны , рожая жизнь молодым. Тем не менее, пять видов монотримы, утконос и четыре вида эхидны, откладывают яйца. Монотримы имеют систему определения пола, отличная от системы большинства других млекопитающих. [ 132 ] В частности, сексуальные хромосомы утконоса больше похожи на хромосовые, чем у млекопитающего Териана. [ 133 ]
Вивипарусные млекопитающие находятся в подклассной терии; Те, кто живут сегодня, находятся в сумчатых и плацентарных инфракласах. Компания имеет короткий период беременности , обычно короче, чем его эстральный цикл , и, как правило, рожает ряд неразвитых новорожденных, которые затем подвергаются дальнейшему развитию; Во многих видах это происходит в мешочке, похожую на мешок, марсупий матери , расположенный в передней части живота . Это плезиоморфное состояние среди вивипарусных млекопитающих; Наличие эпипубических костей у всех неплочных млекопитающих предотвращает расширение торса, необходимого для полной беременности. [ 80 ] Даже непласентальные эвтериан, вероятно, воспроизводили этот путь. [ 42 ] Плаценты рожают относительно полные и развитые молодые, обычно после длительных периодов беременности. [ 134 ] Они получают свое название от плаценты , которая соединяет развивающийся плод с стенкой матки, чтобы обеспечить поглощение питательных веществ. [ 135 ] У плацентарных млекопитающих эпипубик либо полностью утерян, либо преобразуется в бакулум; Позволяя туловище иметь возможность расширяться и, следовательно, при рождении развилось потомство. [ 131 ]
Молочные железы млекопитающих специализируются на производстве молока, основного источника питания для новорожденных. Монотрейцы рано разветвлялись от других млекопитающих и не имеют сосков у большинства млекопитающих, но у них есть молочные железы. Молодой лизнут молоко из молочного участка на животе матери. [ 136 ] По сравнению с плацентарными млекопитающими молоко сумчатых сильно изменяется как в скорости производства, так и в составе питательных веществ из -за слаборазвитой молодых. Кроме того, молочные железы обладают большей автономией, позволяющей им поставлять отдельные молодые молодые на разных этапах разработки. [ 137 ] Лактоза является основным сахаром в плацентарном молоке млекопитающих, в то время как в монотриме и сумчатном молоке преобладают олигосахариды . [ 138 ] Отлучение - это процесс, в котором млекопитающее становится менее зависимым от молока их матери и больше от твердой пищи. [ 139 ]
Эндотермия
Почти все млекопитающие эндотермические («теплые»). У большинства млекопитающих также есть волосы, чтобы помочь им согреться. Как и птицы, млекопитающие могут добывать или охотиться в погоде и слишком холодный климат для эктотермических («хладнокровных») рептилий и насекомых. Эндотермия требует много пищевой энергии, поэтому млекопитающие едят больше пищи на единицу массы тела, чем большинство рептилий. [ 140 ] Небольшие насекомоядные млекопитающие едят потрясающие количества для своего размера. Редкое исключение, обнаженная молька производит небольшую метаболическую тепло, поэтому он считается эксплуатационной пуикилотермой . [ 141 ] Птицы также эндотермические, поэтому эндотермия не уникальна для млекопитающих. [ 142 ]
Виды продолжительность жизни
Среди млекопитающих максимальная продолжительность жизни видов значительно варьируется (например, срок службы в течение двух лет, в то время как самый старый кит -лубовый кот . [ 143 ] Хотя основная основа для этих различий в продолжительности жизни все еще является неопределенной, многочисленные исследования показывают, что способность восстанавливать повреждение ДНК является важным фактором, определяющим срок службы млекопитающих. В исследовании Hart and Setlow 1974 года. [ 144 ] Было обнаружено, что способность восстановления удаления ДНК систематически увеличивается с продолжительностью срока службы видов среди семи видов млекопитающих. Наблюдалось, что срок службы видов надежно коррелировала с способностью распознавать двойные разрывы ДНК, а также уровень белка, репаратного ДНК, Ku80 . [ 143 ] В исследовании клеток из шестнадцати видов млекопитающих, было обнаружено, что гены, используемые в репарации ДНК, повышаются у долгоживущих видов. [ 145 ] Было обнаружено , что клеточный уровень ферментного фермента, репаратированного ДНК , коррелирует с сроком службы видов в исследовании 13 видов млекопитающих. [ 146 ] В трех дополнительных исследованиях различных видов млекопитающих также сообщалось о корреляции между жизнью видов и способностью восстановления ДНК. [ 147 ] [ 148 ] [ 149 ]
Локомоция
Наземный

Большинство позвоночных - амфибии, рептилии и некоторые млекопитающие, такие как люди и медведя - идут по всей нижней стороне ноги. Многие млекопитающие, такие как кошки и собаки, являются цифровыми , ходящими на пальцах ног, тем больше длины шага позволяет больше скорости. Некоторые животные, такие как лошади , являются неглиградом , ходят по кончикам ног. Это еще больше увеличивает их длину шага и, следовательно, их скорость. [ 150 ] Несколько млекопитающих, а именно великие обезьяны, также известно, что они ходят по костяшкам , по крайней мере, за передние ноги. Гигантские муравьеды [ 151 ] и утконоси [ 152 ] также кулаки. Некоторые млекопитающие - двуногие , использующие только две конечности для передвижения, которые можно увидеть, например, у людей и великих обезьян. Двухвосткие виды имеют большую область зрения, чем в четвероногих, сохраняют больше энергии и имеют возможность манипулировать объектами своими руками, что помогает пищу. Вместо того, чтобы ходить, некоторые двуноги прыгают, такие как кенгуру и крысы кенгуру . [ 153 ] [ 154 ]
Животные будут использовать разные походки для разных скоростей, местности и ситуаций. Например, лошади показывают четыре натуральных походки, самая медленная походка для лошадей - это прогулка , а затем есть три более быстрые походки, которые, от самой медленной до самой быстрой, - это рысь , галоп и галоп . У животных также могут быть необычные походки, которые используются иногда, например, для движения вбок или назад. Например, основными человеческими походками являются двуногие ходьба и бегут , но иногда они используют много других походки, включая четвероногий ползание в узких пространствах. [ 155 ] Млекопитающие показывают огромный диапазон походки , порядок, который они размещают и поднимают свои придатки в локомоции. Походки могут быть сгруппированы в категории в соответствии с их моделями поддержки. Для четвероногих есть три основные категории: ходячие походки, бегущие походки и прыгающие походки . [ 156 ] Ходьба - самая распространенная походка, где на земле есть несколько ног в любой момент времени, и почти на всех ногах. Считается, что бег происходит, когда в некоторых моментах в ходу все ноги находятся за землей в момент подвески. [ 155 ]
Дервара

Дервареальные животные часто имеют удлиненные конечности, которые помогают им пересекать зазоры, достигать плодов или других ресурсов, проверять твердость поддержки впереди, а в некоторых случаях - крэхиат (развернуться между деревьями). [ 157 ] Многие древесные виды, такие как дикобразы для деревьев, шелковистые муравьеды , пауковые обезьяны и опоссумы , используют претенсивные хвосты для захвата ветвей. У пауковой обезьяны кончик хвоста имеет либо обнаженное пятно, либо клейкую площадку, которая обеспечивает повышенное трение. Когти могут использоваться для взаимодействия с грубыми субстратами и переориентации направления сил, которое применяет животное. Это то, что позволяет белкам подниматься на стволы деревьев, которые настолько велики, чтобы быть по существу плоскими с точки зрения такого маленького животного. Тем не менее, когти могут мешать способности животного понимать очень маленькие ветви, так как они могут обернуться слишком далеко и уколоть собственную лапу животного. Воплощение трения используется приматами, полагаясь на неровные кончики пальцев. Сжатие ветви между кончиками пальцев генерирует силу трения, которая удерживает руку животного к ветке. Однако этот тип сцепления зависит от угла силы трения, таким образом, от диаметра ветви, с большими ветвями, что приводит к снижению способности захвата. Чтобы контролировать спуск, особенно вниз по ветвям больших диаметров, некоторые древесные животные, такие как белки, развили очень мобильные суставы лодыжки, которые позволяют вращать ногу в «обратную» позу. Это позволяет когтям зацепиться в шероховатую поверхность коры, выступая против силы тяжести. Небольшой размер обеспечивает много преимуществ для древесных видов: такие как увеличение относительного размера ветвей для животного, нижний центр масс, повышенная стабильность, более низкая масса (позволяя двигаться на небольших ветвях) и способность перемещаться по более загроможденной среде обитания. [ 157 ] Размер, связанный с весом, влияет на скользящих животных, таких как сахарный планер . [ 158 ] Некоторые виды приматов, летучих мышей и всех видов ленивца достигают пассивной стабильности, висящей под ветвью. Как качка, так и чаевые становятся неактуальными, поскольку единственным методом отказа был бы потерять свою хватку. [ 157 ]
Антенна
Летучие мыши - единственные млекопитающие, которые могут по -настоящему летать. Они летят по воздуху на постоянной скорости, перемещая свои крылья вверх и вниз (обычно с некоторым передним движением). Поскольку животное находится в движении, есть некоторый поток воздуха по сравнению с его телом, который в сочетании со скоростью крыльев генерирует более быстрый поток воздуха, движущийся над крылом. Это генерирует вектор силы подъема, указывающий вперед и вверх, и вектор силы сопротивления, указывающий назад и вверх. Вверх компоненты этих противодействий тяжести, удерживая тело в воздухе, в то время как передний компонент обеспечивает тягу, чтобы противодействовать как сопротивлению от крыла, так и от тела в целом. [ 159 ]
Крылья летучих мышей намного тоньше и состоят из большего количества костей, чем у птиц, что позволяет летучим мышам более точно маневрировать и летать с большим количеством подъема и меньше сопротивления. [ 160 ] [ 161 ] Складывая крылья внутрь к их телу наверх, они используют на 35% меньше энергии во время полета, чем птицы. [ 162 ] Мембраны деликатны, легко разрываются; Тем не менее, ткань мембраны летучей мыши способна отрастать, так что маленькие слезы могут быстро залечить. [ 163 ] Поверхность их крыльев оснащена чувствительными рецепторами на небольших ударах, называемых клетками Меркель , также обнаруженными на кончиках пальцев человека. Эти чувствительные участки различны в летучих мышах, так как у каждого удара есть крошечные волосы в центре, что делает их еще более чувствительными и позволяя биту обнаружить и собирать информацию о воздухе, текущем по крыльям, и более эффективно летать путем изменения формы его крылья в ответ. [ 164 ]
Фоссол и подземный


Fossorial (от латинского Fossor , что означает «копатель») - это животное, адаптированное к копанию, которое живет в первую очередь, но не только под землей. Некоторые примеры- Барсуки и голые моль-скалы . Многие виды грызунов также считаются подачами, потому что они живут в норах для большинства, но не весь день. Виды, которые живут исключительно под землей, являются подземными, и с ограниченными адаптациями к подкопсу Fossorial Lifestyle. Некоторые организмы являются искоренными, чтобы помочь в регулировании температуры , в то время как другие используют подземную среду обитания для защиты от хищников или для хранения продуктов питания . [ 165 ]
Фоссористые млекопитающие имеют вездее тело, самого толстого у плеч и сужающиеся у хвоста и носа. Неспособность видеть в темных норах, большинство имеют вырожденные глаза, но дегенерация варьируется между видами; Например, карманные суслики являются лишь полупрофессиональными и имеют очень маленькие, но функциональные глаза, в полностью истешенной баранической моле Глаза вырождаются, а бесполезные, кроты талпа имеют рудиментарные глаза, а у мыса золотой моль есть слой кожи, покрывающий глаза Полем Внешние уши клапаны также очень маленькие или отсутствуют. Поистине высыпанные млекопитающие имеют короткие, крепкие ножки, так как сила важнее, чем скорость для мучительного млекопитающего, но у полупроводных млекопитающих изгибные ножки. Передние лапы широкие и имеют сильные когти, чтобы помочь в ослаблении грязи при раскопок, а задняя лапы имеют лямки, а также когти, что помогает бросить ослабленную грязь назад. У большинства есть большие резцы, чтобы не дать грязи летать во рту. [ 166 ]
Многие вынужденные млекопитающие, такие как застройки, ежи и родинки, были классифицированы под ныне устаревшим порядком насекомого . [ 167 ]
Водный
Полностью водные млекопитающие, китообразные и сирены , потеряли ноги и имеют хвостовой плавник, чтобы продвинуться через воду. Движение Flipper непрерывное. Киты плавают, перемещая свой хвостовой плавник и нижнюю часть тела вверх и вниз, выдвигая себя через вертикальное движение, в то время как их ласты в основном используются для рулевого управления. Их скелетная анатомия позволяет им быть быстрыми пловцами. У большинства видов есть спинный плавник , чтобы не допустить вверх ногами в воде. [ 168 ] [ 169 ] Желебные переводы поднимаются вверх и вниз в длинных ударах, чтобы перемещать животное вперед, и могут быть скручены, чтобы повернуться. Передатки-это ласточки, похожие на весло, которые помогают поворачиваться и замедляться. [ 170 ]
Полуаватические млекопитающие, такие как выносливые, имеют две пары ластаков спереди и сзади, передовые пластики и задние плитки. Локти и лодыжки заключены в тело. [ 171 ] [ 172 ] Уносимых имеют несколько адаптаций для уменьшения сопротивления . В дополнение к их обтекаемым телам, они имеют гладкие сети мышечных пучков в коже, которые могут увеличить ламинарный поток и облегчить им проскальзывать через воду. Им также не хватает пили , поэтому их мех может быть упорядочен во время плавания. [ 173 ] Они полагаются на свои передовые ток-липперы за локомоцию в похожей на крыло, похожее на пингвины и морские черепахи . [ 174 ] Переднее движение непрерывнее, а животное скользит между каждым ударом. [ 172 ] По сравнению с наземными плотовальниками, передние конечности уменьшаются по длине, что дает локомоторные мышцы на плече и локтевых суставах большее механическое преимущество; [ 171 ] Задние плитки служат стабилизаторами. [ 173 ] Другие полугодовые млекопитающие включают бобры, бегемоты , выдры и уткономе. [ 175 ] Гиппоты-очень большие полуаком-млекопитающие, а их бочкообразные тела имеют гравипортала , скелетные структуры [ 176 ] Адаптируется к ношению их огромного веса, и их удельная гравитация позволяет им погрузиться и двигаться вдоль дна реки. [ 177 ]
Поведение
Общение и вокализация

Многие млекопитающие общаются, вокалируя. Вокальное общение служит многим целям, в том числе в ритуалах спаривания, как предупреждающие звонки , [ 179 ] указывать источники пищи и для социальных целей. Мужчины часто вызывают во время ритуалов спаривания, чтобы отразить других мужчин и привлекать женщин, как в рев Львов оленей и красных . [ 180 ] Песни ; горбатого кита могут быть сигналами для женщин [ 181 ] У них разные диалекты в разных регионах океана. [ 182 ] Социальные вокализации включают территориальные призывы Гиббонов и использование частоты в больших копьях летучих мышах для различения групп. [ 183 ] Обезьяна Vervet дает отчетливый сигнал тревоги для каждого из четырех разных хищников, и реакции других обезьян варьируются в зависимости от вызова. Например, если сигнал тревоги сигнализирует о питоне, обезьяны забираются в деревья, тогда как будильник орла заставляет обезьян искать укрытие на земле. [ 178 ] У Prairie Dogs также есть сложные вызовы, которые сигнализируют о типе, размере и скорости приближающегося хищника. [ 184 ] Слоны социально общаются с различными звуками, в том числе фырканье, кричание, трубичую, ревущие и грохочущие. Некоторые из грохочущих вызовов инфразсонки , ниже линейки слуха людей, и могут быть услышаны другими слонами до 6 миль (9,7 км) в неподвижном временах, возле рассвета и заката. [ 185 ]
Млекопитающие сигнализируют различными средствами. Многие дают визуальные сигналы анти-преседатора , как, когда, когда оленя и газель их состояние подгонки , честно указывая на и их способность убежать, [ 186 ] [ 187 ] или когда белохвостые оленя и другие добычные млекопитающие флаг с заметной хвостовой маркировкой, когда встревожены, сообщив хищнику, что он был обнаружен. [ 188 ] Многие млекопитающие используют маркировку ароматов , иногда, возможно, чтобы помочь защитить территорию, но, вероятно, с целым рядом функций как внутри, так и между видами. [ 189 ] [ 190 ] [ 191 ] Микробаты и зубчатые киты, в том числе океанические дельфины, озвучивают как социально, так и в эхолокации . [ 192 ] [ 193 ] [ 194 ]
Кормление
Для поддержания высокой постоянной температуры тела является энергетической дорогой - поэтому млекопитающие нуждаются в питательной и обильной диете. В то время как самые ранние млекопитающие были, вероятно, хищники, с тех пор разные виды адаптировались для удовлетворения своих диетических требований различными способами. Некоторые едят других животных - это хищная диета (и включает насекомоядную диету). Другие млекопитающие, называемые травоядными , едят растения, которые содержат сложные углеводы, такие как целлюлоза. Травоядница включает в себя подтипы, такие как граниватор (еда семян), фоливори (питание листьев), фругиварив (питание с фруктами), нектаривное (нектарное питание), Gumvivory (еда для жвачки) и микофагию (еда грибков). Пищеварительный тракт травоядного животного - это хозяин бактерий, которые ферментируют эти сложные вещества, и делают их доступными для пищеварения, которые либо размещены в многоасинентном желудке , либо в большой слепой кишке. [ 93 ] Некоторые млекопитающие копра , потребляющие фекалии для поглощения питательных веществ, не перевариваемых, когда пища впервые проглатывалась. [ 89 ] : 131–137 Всеядное питание ест как добычу и растения. Утояемые млекопитающие имеют простой пищеварительный тракт , потому что белки , липиды и минералы, обнаруженные в мясе Исключения из этого включают Baleen Whales , которые также размещают кишечную флору в многоканированном желудке, как наземные травоядные животные. [ 195 ]
Размер животного также является фактором определения типа диеты ( правило Аллена ). Поскольку небольшие млекопитающие имеют высокое соотношение термоясовой площади поверхности к тепловой объеме, они имеют тенденцию иметь высокие потребности в энергии и высокую скорость метаболизма . Млекопитающие, которые весят менее 18 унций (510 г; 1,1 фунта), в основном насекомые, потому что они не могут переносить медленный, сложный пищеварительный процесс травоядного. Более крупные животные, с другой стороны, генерируют больше тепла и меньше этого тепла теряются. Следовательно, они могут переносить либо более медленный процесс сбора (плотоядные, которые питаются более крупными позвоночными), либо более медленным пищеварительным процессом (травоядные). [ 196 ] Кроме того, млекопитающие, которые весят более 18 унций (510 г; 1,1 фунта), обычно не могут собирать достаточно насекомых в свои бодрствующие часы, чтобы поддерживать себя. Единственные крупные насекомоядные млекопитающие - это те, которые питаются огромными колониями насекомых ( муравьев или термитов ). [ 197 ]


Некоторые млекопитающие являются всеядными и демонстрируют различную степень плотоядного и травоядного, как правило, наклоняются в пользу одного больше, чем другое. Поскольку растения и мясо расщепляются по -разному, предпочитают одно над другим, как у медведей, где некоторые виды могут быть в основном плотоядными, а другие в основном травоядные. [ 199 ] Они сгруппированы в три категории: мезогарнословия (50–70% мяса), гиперкарнословия (70% и больше мяса) и гипокарнословия (50% или менее мяса). Улечение гипокарноводов состоит из тусклых, треугольных карнассиальных зубов, предназначенных для измельчения пищи. Гиперкарноворы, однако, имеют конические зубы и острые карнассии, предназначенные для сокращения, а в некоторых случаях сильные челюсти для костей, как в случае гиен , позволяя им потреблять кости; В некоторых вымерших группах, в частности, у Machairodontinae в форме сабли , были клыки . [ 198 ]
Некоторые физиологические хищники потребляют растительные вещества, а некоторые физиологические травоядные животные потребляют мясо. С поведенческого аспекта это сделало бы их всеядными, но с физиологической точки зрения это может быть связано с зоофармакогнозией . Физиологически животные должны быть в состоянии получить как энергию, так и питательные вещества из материалов растений и животных, которые считаются всеядными. Таким образом, такие животные по -прежнему могут классифицироваться как плотоядные и травоядные, когда они просто получают питательные вещества из материалов, происходящих из источников, которые, по -видимому, не дополняют их классификацию. [ 200 ] Например, хорошо задокументировано, что некоторые заболевания, такие как жирафы, верблюды и крупный рогатый скот, будут грызть кости, чтобы потреблять определенные минералы и питательные вещества. [ 201 ] Кроме того, кошки, которые, как правило, рассматриваются как облигатные плотоядные, иногда едят траву, чтобы извергнуть неперевариваемое материал (например, волосы ), помощь с производством гемоглобина и в качестве слабительного. [ 202 ]
Многие млекопитающие, в отсутствие достаточных требований к продуктам питания в окружающей среде, подавляют их метаболизм и сохраняют энергию в процессе, известном как спячка . [ 203 ] В предыдущий период спячки более крупные млекопитающие, такие как медведи, становятся полифагическими для увеличения запасов жира, тогда как меньшие млекопитающие предпочитают собирать и хранить пищу. [ 204 ] Замедление метаболизма сопровождается сниженной скоростью сердца и дыхания, а также падением внутренних температур, которые в некоторых случаях могут быть вокруг температуры окружающей среды. Например, внутренние температуры изгибающих арктических заземляющих белок могут упасть до -2,9 ° C (26,8 ° F); Однако голова и шея всегда остаются выше 0 ° C (32 ° F). [ 205 ] Несколько млекопитающих в горячей среде эспетивируют во времена засухи или сильной жары, например, жирного хвостого карликового лемура ( Cheirogaleus medius ). [ 206 ]
Питьевой

По необходимости наземные животные в неволе привыкли к питьевой воде, но большинство животных свободного воспитания остаются увлажненными через жидкости и влагу в свежих продуктах. [ 207 ] и научитесь активно искать продукты с высоким содержанием жидкости. [ 208 ] Когда условия побуждают их пить из тел воды, методы и движения сильно различаются среди видов. [ 209 ]
Кошки , клыки и жвачные животные все опускают шею и колен в воде своими мощными языками. [ 209 ] Кошки и клыки впитывают воду с языком в виде ложки. [ 210 ] Клыки на коленях вода, выкопав ее в рот языком, который принял форму ковша. Однако с кошками только кончик языка (который гладкий) касается воды, и затем кошка быстро тянет язык обратно в рот, который вскоре закрывается; Это приводит к тому, что колонна жидкости втягивается в рот кошки, который затем закрепляется на закрытие рта. [ 211 ] Жвачные животные и большинство других травоядных, частично погружают кончик рта, чтобы привлечь воду с помощью погружного действия с языком, держащим прямо. [ 212 ] Кошки пьют значительно более медленными темпами, чем жвачные животные, которые сталкиваются с большими естественными опасностями хищников. [ 209 ]
Многие пустынные животные не пьют, даже если вода становится доступной, но полагается на употребление сочных растений . [ 209 ] В холодной и замороженной среде некоторые животные, такие как зайцы , белки деревьев и Bighorn Resort к употреблению снега и сосульков. [ 213 ] В саваннах метод питья жирафов был источником спекуляций за его очевидное неповиновение гравитации; Самая последняя теория предусматривает, что длинная шея животного функционирует как насос плунжера . [ 214 ] Уникально слоны втягивают воду в свои стволы и впрыскивают ее в рот. [ 209 ]Интеллект
У интеллектуальных млекопитающих, таких как приматы , головность больше по сравнению с остальной частью мозга. Сам интеллект нелегко определить, но показания интеллекта включают в себя способность учиться, сопоставленные с поведенческой гибкостью. Например, крысы считаются очень умными, поскольку они могут учиться и выполнять новые задачи, способность, которая может быть важной, когда они впервые колонизируют свежую среду обитания . У некоторых млекопитающих сбор пищи, по -видимому, связан с интеллектом: у оленя, питаясь растениями, мозг меньше кошки, который должен подумать, чтобы перехитрить свою добычу. [ 197 ]

Использование инструментов животными может указывать на различные уровни обучения и познания . Морская выдр использует камни в качестве важных и обычных частей своего поведения в поисках пищи (разбивая ущельщику из камней или разбившиеся открытые раковины), причем некоторые популяции тратят 21% своего времени на создание инструментов. [ 215 ] Другое использование инструмента, такое как шимпанзе , использующие веточки для «рыбы» для термитов, может быть разработано, наблюдая за тем, как другие используют инструменты и могут быть даже настоящим примером обучения животных. [ 216 ] Инструменты могут даже использоваться при решении головоломки, в которых животное, по -видимому, испытывает «момент эврики» . [ 217 ] Другие млекопитающие, которые не используют инструменты, такие как собаки, также могут испытать момент Eureka. [ 218 ]
Размер мозга ранее считался основным показателем интеллекта животного. Поскольку большая часть мозга используется для поддержания телесных функций, большие отношения мозга к массе тела могут увеличить количество массы мозга, доступную для более сложных когнитивных задач. Аллометрический анализ указывает на то, что размер мозга млекопитающих масштабируется примерно на 2 ~ 3 или 3 ~ 4 показания массы тела. Сравнение размер мозга конкретного животного с ожидаемым размером мозга на основе такого аллометрического анализа обеспечивает коэффициент энцефализации , который можно использовать в качестве еще одного показания интеллекта животных. [ 219 ] Сперматозоиды имеют самую большую массу мозга среди всех животных на земле, в среднем 8000 кубических сантиметров (490 куб. [ 220 ]
Самосознание, по-видимому, является признаком абстрактного мышления. Считается, что самосознание, хотя и не определенное, является предшественником более продвинутых процессов, таких как метакогнитивные рассуждения . Традиционный метод измерения этого-это зеркальный тест , который определяет, обладает ли животным способностью самопознания. [ 221 ] Млекопитающие, прошедшие тест на зеркал, включают азиатских слонов (некоторые проходят, некоторые нет); [ 222 ] шимпанзе; [ 223 ] бонобо ; [ 224 ] орангутаны ; [ 225 ] люди, с 18 месяцев ( зеркальная стадия ); [ 226 ] общие узкие дельфины ; [ А ] [ 227 ] Орки ; [ 228 ] и ложные убийцы китов . [ 228 ]
Социальная структура

Euscociality является самым высоким уровнем социальной организации. Эти общества имеют совпадение взрослых поколений, разделения репродуктивного труда и кооперативной заботы о молодежи. Обычно насекомые, такие как пчелы , муравьи и термиты, имеют эусоциальное поведение, но это продемонстрировано у двух видов грызунов: голый моль-рат [ 229 ] и Дамараленд Моль-Рэт . [ 230 ]
Предварительность - это когда животные демонстрируют больше, чем просто сексуальные взаимодействия с членами одного и того же вида, но не соответствуют квалификации как Eusocial. То есть преподобные животные могут демонстрировать общинную жизнь, кооперативную уход за молодым или примитивное разделение репродуктивного труда, но они не демонстрируют все три важные признаки эусоциальных животных. Люди и некоторые виды Callitrichidae ( Marmosets и Tamarins ) являются уникальными среди приматов в их степени кооперативного ухода за молодыми. [ 231 ] Гарри Харлоу организовал эксперимент с резус -обезьянами , преоциональными приматами, в 1958 году; Результаты этого исследования показали, что социальные встречи необходимы для того, чтобы молодые обезьяны развивались как умственно, так и сексуально. [ 232 ]
Общество деления -фьюзия - это общество, которое часто меняется в своем размере и композиции, составляющая постоянную социальную группу под названием «родительская группа». Постоянные социальные сети состоят из всех отдельных членов сообщества и часто варьируются для отслеживания изменений в своей среде. В обществе деления -фьюзина основная родительская группа может перелома (деление) в более мелкие стабильные подгруппы или отдельных лиц для адаптации к окружающей среде или социальным обстоятельствам. Например, многие мужчины могут оторваться от основной группы, чтобы охотиться или на корм для еды в течение дня, но ночью они могут вернуться, чтобы присоединиться (Fusion) основной группе, чтобы делиться едой и участвовать в других мероприятиях. Многие млекопитающие демонстрируют это, такие как приматы (например, орангутаны и обезьяны -пауки ), [ 233 ] слоны, [ 234 ] пятнистые гиены , [ 235 ] львы, [ 236 ] и дельфины. [ 237 ]
Одиночные животные защищают территорию и избегают социальных взаимодействий с членами его видов, за исключением сезона размножения. Это значит избегать конкуренции за ресурсами, так как два особенности одного и того же вида будут занимать одну и ту же нишу и предотвратить истощение пищи. [ 238 ] Одиночное животное, во время кормления, также может быть менее заметным для хищников или добычи. [ 239 ]

В иерархии люди либо доминируют, либо покорны. Деспотическая иерархия - это то, где один человек является доминирующим, в то время как остальные покорны, как у волков и лемуров, [ 240 ] И порядок клевания - это линейный рейтинг людей, где есть лидерный человек и нижний человек. Заказы о клевете также могут быть ранжированы по сексу, где самый низкий индивид в сексе имеет более высокий рейтинг, чем лучший человек из другого пола, как у гиен. [ 241 ] Доминирующие люди, или альфа, имеют высокую вероятность репродуктивного успеха, особенно в гаремах , где один или несколько мужчин (жителей) имеют исключительные права размножения для женщин в группе. [ 242 ] Мужчины нерезидента также могут быть приняты в гаремах, но некоторые виды, такие как общая летучая мышь вампира ( Desmodus rotundus ), могут быть более строгими. [ 243 ]
Некоторые млекопитающие являются совершенно моногамными , что означает, что они спариваются на всю жизнь и не берут других партнеров (даже после смерти первоначального партнера), как и в случае с волками, евразийскими бобрами и выдрами. [ 244 ] [ 245 ] Существует три типа многоженства: один или множественный доминантный мужчины имеют права на размножение ( полигиния ), множество мужчин, с которыми спаривают женщины (полиандрие), или несколько мужчин имеют эксклюзивные отношения с несколькими женщинами ( полигинандри ). Полигинозное спаривание гораздо чаще встречается, что, за исключением лек , по оценкам, встречается до 90% млекопитающих. [ 246 ] Спечное спаривание происходит, когда мужчины собираются вокруг женщин и пытаются привлечь их различными показателями ухаживания и вокализация, как в гавани. [ 247 ]
Все более высокие млекопитающие (исключая монотримы) имеют две основные адаптации для ухода за молодым: живорождение и лактация. Они подразумевают в группе выбор степени родительской помощи . Они могут строить гнезда и копать норы, чтобы поднять своих молодых или кормить и охранять их часто в течение длительного периода времени. Многие млекопитающие выбираются в K и вкладывают больше времени и энергии в своих молодых выбранных R. животных, Когда двое животных спариваются, они оба имеют интерес к успеху потомства, хотя часто к разным крайностям. Самки млекопитающих демонстрируют некоторую степень материнской агрессии, еще один пример родительской помощи, который может быть нацелен на других женщин вида или молодых женщин; Тем не менее, некоторые млекопитающие могут «тетя» младенцев других женщин и заботиться о них. Мужчины млекопитающих могут играть роль в воспитании детей, как и в случае с TenRecs, однако это варьируется с видами, даже в рамках одного рода. Например, мужчины южной свиньи макаки ( Macaca Nemestrina ) не участвуйте в уходе за ребенком, тогда как мужчины японской макаки ( M. fuscata ) делают. [ 248 ]
Люди и другие млекопитающие
В человеческой культуре

Нечеловеческие млекопитающие играют множество ролей в человеческой культуре. Они являются самыми популярными из домашних животных , с десятками миллионов собак, кошек и других животных, включая кроликов и мышей, хранящихся в семьях по всему миру. [ 249 ] [ 250 ] [ 251 ] Млекопитающие, такие как мамонт , лошади и оленя, являются одними из самых ранних предметов искусства, находящихся на картинах верхней палеолитической пещеры, таких как в Lascaux . [ 252 ] Крупные артисты, такие как Альбрехт Дюрер , Джордж Стаббс и Эдвин Ландсир, известны своими портретами млекопитающих. [ 253 ] Многие виды млекопитающих охотились за спортом и за пищу; Олень и дикий кабан особенно популярны как игровые животные . [ 254 ] [ 255 ] [ 256 ] Млекопитающие, такие как лошади и собаки, широко учатся в спорте, часто в сочетании с ставками на результат . [ 257 ] [ 258 ] Существует напряженность между ролью животных как компаньонов для людей, и их существование как личности с собственными правами . [ 259 ] Млекопитающие далее играют широкий спектр ролей в литературе, [ 260 ] [ 261 ] [ 262 ] фильм, [ 263 ] Мифология и религия. [ 264 ] [ 265 ] [ 266 ]
Использование и важность

Одомашнивание млекопитающих сыграло важную роль в неолитическом развитии сельского хозяйства и цивилизации , заставляя фермеров заменить охотников-собирателей по всему миру. [ B ] [ 268 ] Этот переход от охоты и сбора к пасти и выращиванию культур был серьезным шагом в истории человечества. Новая сельскохозяйственная экономика, основанная на одомашненных млекопитающих, вызвала «радикальную реструктуризацию человеческих обществ, мировые изменения в биоразнообразии и значительные изменения в формах земель и ее атмосферу ... знамительные результаты». [ 269 ]
Внутренние млекопитающие составляют большую часть скота, поднятого для мяса по всему миру. Они включают (2009) около 1,4 миллиарда крупного рогатого скота , 1 миллиард овец , 1 миллиард домашних свиней , [ 270 ] [ 271 ] и (1985) более 700 миллионов кроликов. [ 272 ] Рабочие домашние животные, в том числе крупный рогатый скот и лошадей, использовались для работы и транспортировки из происхождения сельского хозяйства, их число сокращалось с появлением механизированного транспорта и сельскохозяйственного оборудования . В 2004 году они по -прежнему обеспечивали около 80% власти для в основном небольших ферм в третьем мире, а также около 20% мирового транспорта, опять же, в основном в сельских районах. В горных регионах, не подходящих для колесных транспортных средств, упаковывают животные, продолжают транспортировать товары. [ 273 ] Шкины млекопитающих обеспечивают кожу для обуви , одежды и обивки . Шерсть от млекопитающих, включая овец, коз и альпак, веками использовалась для одежды. [ 274 ] [ 275 ]
Млекопитающие играют важную роль в науке в качестве экспериментальных животных , как в фундаментальных биологических исследованиях, таких как генетика, [ 277 ] и в разработке новых лекарств, которые должны быть протестированы, чтобы продемонстрировать их безопасность . [ 278 ] Миллионы млекопитающих, особенно мышей и крыс, используются в экспериментах каждый год. [ 279 ] Нокаутная мышь представляет собой генетически модифицированную мышь с инактивированным геном , замененной или нарушенной искусственным произведением ДНК. Они позволяют изучать секвенированные гены, функции которых неизвестны. [ 280 ] Небольшой процент млекопитающих-это нечеловеческие приматы, используемые в исследованиях для их сходства с людьми. [ 281 ] [ 282 ] [ 283 ]
Несмотря на преимущества одомашненных млекопитающих для развития человека, люди оказывают все более вредное влияние на диких млекопитающих по всему миру. Было подсчитано, что масса всех диких млекопитающих снизилась до 4% всех млекопитающих, причем 96% млекопитающих являются людьми и их скотом (см. Рисунок). На самом деле, наземные дикие млекопитающие составляют только 2% всех млекопитающих. [ 284 ] [ 285 ]
Гибриды


Гибриды - это потомство в результате размножения двух генетически различных индивидуумов, что обычно приведет к высокой степени гетерозиготности, хотя гибридные и гетерозиготные не являются синонимом. Преднамеренная или случайная гибридизация двух или более видов близкородственных животных посредством размножения в неволе - это человеческая деятельность, которая существует на протяжении тысячелетий и выросла для экономических целей. [ 286 ] Гибриды между различными подвидами внутри вида (например, между бенгальским тигра и сибирского тигра ) известны как внутриспецифические гибриды. Гибриды между различными видами в пределах одного рода (например, между львами и тиграми) известны как межвидовые гибриды или скрещивания. Гибриды между различными родами (например, между овец и коз) известны как межпогничные гибриды. [ 287 ] Природные гибриды будут происходить в гибридных зонах , где две популяции видов в одних и тех же родах или видах, живущих в одних и тех же или прилегающих районах, будут перемещаться друг с другом. Некоторые гибриды были признаны видами, такими как красный волк (хотя это спорно). [ 288 ]
Искусственный отбор , преднамеренное избирательное размножение домашних животных, используется для разведения недавно вымерших животных в попытке достичь породы животных с фенотипом , который напоминает этот вымерший предок дикого типа . Гибрид размножения (внутривидового) гибрида может быть очень похож на вымерший вид, экологическую нишу и в некоторой степени генетику, но начальный генный пул этого дикого типа навсегда теряется с его вымиранием . В результате породы Bred Back в лучшем случае являются смутными взглядами вымерших диких вещей, поскольку, черт возьми, крупный рогатый скот из Aurochs . [ 289 ]
Чистокровные дикие виды, развитые до определенной экологии, могут быть угрожают исчезновением [ 290 ] Благодаря процессу генетического загрязнения неконтролируемая гибридизация, интрогрессия генетическая болота, которая приводит к гомогенизации или концентрации от гетерозических гибридных видов. [ 291 ] Когда новые популяции импортируются или выборочно разводятся людьми или когда модификация среды обитания приводит к контакту ранее изолированных видов, возможно, вымирание у некоторых видов, особенно редких сортов, возможно. [ 292 ] Перестрографирование может затопить более редкий генный пул и создавать гибриды, истощая чистокровный генный пул. Например, исчезающему буйволам дикой воды наиболее под угрозой исчезновения генетическим загрязнением из домашнего водного буйвола . Такие вымирания не всегда очевидны с морфологической точки зрения. Некоторая степень потока генов является нормальным эволюционным процессом, тем не менее, гибридизация угрожает существованию редких видов. [ 293 ] [ 294 ]
Угрозы

Потеря видов из экологических сообществ, дефицита , в первую очередь обусловлена человеческой деятельностью. [ 295 ] Это привело к пустым лесам , экологические сообщества истощены от крупных позвоночных. [ 296 ] [ 297 ] В событии четвертичного вымирания массовое умирание разнообразия мегафауна совпала с появлением людей, что предполагает влияние человека. Одна гипотеза заключается в том, что люди охотились на крупных млекопитающих, таких как шерстяной мамонт , в вымирание. [ 298 ] [ 299 ] В отчете о глобальной оценке 2019 года о биоразнообразии и экосистемных услугах IPBE говорится , что общая биомасса диких млекопитающих снизилась на 82 процента с начала человеческой цивилизации. [ 300 ] [ 301 ] Дикие животные составляют всего 4% биомассы млекопитающих на земле, в то время как люди и их одомашненные животные составляют 96%. [ 285 ]
Предполагается, что различные виды вымерли в ближайшем будущем , [ 302 ] Среди них носорог , [ 303 ] жирафы , [ 304 ] и виды приматов [ 305 ] и панголины . [ 306 ] WWF о живой планете на 2020 год Согласно отчету , с 1970 года популяции диких животных позвоночных сократились на 68% в результате деятельности человека, в частности, чрезмерного потребления , роста населения и интенсивного сельского хозяйства , что является доказательством того, что люди вызвали шестое событие массового вымирания . [ 307 ] [ 308 ] Одна охота угрожает сотням видов млекопитающих по всему миру. [ 309 ] [ 310 ] Ученые утверждают, что растущий спрос на мясо способствует потере биоразнообразия, поскольку это является значительным движением обезлесения и разрушения среды обитания ; Богатые виды обитания, такие как значительные части тропического леса Амазонки , превращаются в сельскохозяйственные земли для производства мяса. [ 311 ] [ 312 ] [ 313 ] Другим влиянием является чрезмерное охота и браконьерство , которое может уменьшить общую популяцию животных животных, [ 314 ] особенно те, которые расположены рядом с деревнями, [ 315 ] как в случае пекари . [ 316 ] Эффекты браконьерства особенно можно увидеть в торговле слоновой костью с африканскими слонами. [ 317 ] Морские млекопитающие подвергаются риску запутывания от рыболовного снаряжения, в частности, китообразных , с отброшенными смертностью от 65 000 до 86 000 человек в год. [ 318 ]
Внимание уделяется исчезающим видам во всем мире, в частности, посредством соглашения о биологическом разнообразии , иначе известном как Аккорь Рио, которое включает 189 подписавших стран, которые сосредоточены на выявлении исчезающих видов и среды обитания. [ 319 ] Другая известная охраняемая организация-МСОП, в которой участвуют более 1200 правительственных и неправительственных организаций. [ 320 ]
Недавние вымирания могут быть напрямую отнести к человеческим влияниям. [ 321 ] [ 295 ] МСКН характеризует «недавнее» вымирание как те, которые произошли после точки отсечения 1500, [ 322 ] И около 80 видов млекопитающих вымерли с того времени и 2015 года. [ 323 ] Некоторые виды, такие как оленя Пера Давида [ 324 ] вымерли в дикой природе и выживают исключительно в пленных популяциях. Другие виды, такие как Флоридская Пантера , экологически вымерли , выживают в таких низких количествах, что они по существу не оказывают влияния на экосистему. [ 325 ] : 318 Другие популяции только локально вымерли (истекло), все еще существуют в другом месте, но уменьшаются в распределении, [ 325 ] : 75–77 Как и в случае вымирания серых китов в Атлантике . [ 326 ]
Смотрите также
- Список родов млекопитающих - живые млекопитающие
- Список млекопитающих
- Список монотримов и сумчатых
- Список плацентарных млекопитающих
- Список доисторических млекопитающих
- Список исчезающих млекопитающих
- Списки млекопитающих по размеру популяции
- Списки млекопитающих по региону
- Млекопитающие, описанные в 2000 -х годах
- Млекопитающие в культуре
- Маленькое млекопитающее
Примечания
- ^ Снижение задержки, чтобы приблизиться к зеркало, повторяющееся головокружение и тщательное просмотр заметных областей считалось признаками самосознания, поскольку у них нет оружия и не может прикоснуться к заметным участкам. [ 227 ]
- ^ Diamond обсудил этот вопрос дальше в своих книжных орудиях, микробах и стали 1997 года . [ 267 ]
Ссылки
- ^ Льюис, Чарлтон Т.; Короткий, Чарльз (1879). "Мама" . Латинский словарь . Цифровая библиотека Персея. Архивировано из оригинала 29 сентября 2022 года . Получено 29 сентября 2022 года .
- ^ Воган Т.А., Райан Дж. М., Чеплвски Нью -Джерси (2013). "Классификация млекопитающих" МАММАЛОГИ (6 -е изд.). Джонс и Барттт обучение. ISBN 978-1-284-03209-3 .
- ^ Симпсон Г.Г. (1945). «Принципы классификации и классификация млекопитающих». Американский музей естественной истории . 85
- ^ Szalay FS (1999). «Классификация млекопитающих выше уровня видов: обзор». Журнал палеонтологии позвоночных . 19 (1): 191–195. doi : 10.1080/02724634.1999.10011133 . ISSN 0272-4634 . JSTOR 4523980 .
- ^ Jump up to: а беременный Уилсон де , Ридер Д.М., ред. (2005). «Предисловие и вводной материал» . Виды млекопитающих мира: таксономическая и географическая ссылка (3 -е изд.). Johns Hopkins University Press . п. XXVI. ISBN 978-0-8018-8221-0 Полем OCLC 62265494 .
- ^ «Млекопитающие» . Красный список МСОП . Международный союз сохранения природы (МСОП). Апрель 2010. Архивировано с оригинала 3 сентября 2016 года . Получено 23 августа 2016 года .
- ^ Burgin CJ, Colella JP, Kahn PL, Upham NS (1 февраля 2018 года). "Сколько там видов млекопитающих?" Полем Журнал млекопитающих . 99 (1): 1–14. doi : 10.1093/jmammal/gyx147 .
- ^ Роу Т (1988). «Определение, диагноз и происхождение млекопитающих» (PDF) . Журнал палеонтологии позвоночных . 8 (3): 241–264. Bibcode : 1988jvpal ... 8..241r . doi : 10.1080/02724634.1988.10011708 . Архивировано (PDF) от оригинала 18 января 2024 года . Получено 25 января 2024 года .
- ^ Lyell C (1871). Студенческие элементы геологии . Лондон: Джон Мюррей. п. 347. ISBN 978-1-345-18248-4 .
- ^ Cifelli RL, Davis BM (декабрь 2003 г.). «Палеонтология. Коричневое происхождение». Наука . 302 (5652): 1899–1900. doi : 10.1126/science.1092272 . PMID 14671280 . S2CID 83973542 .
- ^ Кемп Т.С. (2005). Происхождение и эволюция млекопитающих (PDF) . Великобритания: издательство Оксфордского университета. п. 3. ISBN 978-0-19-850760-4 Полем OCLC 232311794 . Архивировано (PDF) из оригинала 26 сентября 2023 года . Получено 25 января 2024 года .
- ^ Datta PM (2005). «Самое раннее млекопитающее с поперечным расширенным верхним моляром из позднего триасового (карнского) формации тики, южная бассейн Рева Гондвана, Индия». Журнал палеонтологии позвоночных . 25 (1): 200–207. doi : 10.1671/0272-4634 (2005) 025 [0200: emwteu] 2.0.co; 2 . S2CID 131236175 .
- ^ Luo ZX, Martin T (2007). «Анализ молярной структуры и филогения родов docodont» (PDF) . Бюллетень музея естественной истории Карнеги . 39 : 27–47. doi : 10.2992/0145-9058 (2007) 39 [27: aomsap] 2.0.co; 2 . S2CID 29846648 . Архивировано из оригинала (PDF) 3 марта 2016 года . Получено 8 апреля 2013 года .
- ^ Averianov, Alexander O.; Войта, Леонид Л. (март 2024 г.). «Предполагаемый триасовый ствол Tikitherium copei - неоген -строительство» . Журнал эволюции млекопитающих . 31 (1). doi : 10.1007/s10914-024-09703-w . ISSN 1064-7554 .
- ^ Кригс Джо, Чураков Г., Кифманн М., Джордан У, Брозиус Дж., Шмитц Дж. (Апрель 2006 г.). «Ретрополированные элементы как архивы эволюционной истории плацентарных млекопитающих» . PLOS Биология . 4 (4): E91. doi : 10.1371/journal.pbio.0040091 . PMC 1395351 . PMID 16515367 .
- ^ Jump up to: а беременный Нишихара Х., Маруяма С., Окада Н (март 2009 г.). «Анализ ретропозона и недавние геологические данные свидетельствуют о почти сильной дивергенции трех опекунов млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (13): 5235–5240. Bibcode : 2009pnas..106.5235n . doi : 10.1073/pnas.0809297106 . PMC 2655268 . PMID 19286970 .
- ^ Springer MS, Murphy WJ, Eizirik E, O'Brien SJ (февраль 2003 г.). «Диверсификация плацентарных млекопитающих и граница мела -третья» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (3): 1056–1061. Bibcode : 2003pnas..100.1056s . doi : 10.1073/pnas.0334222100 . PMC 298725 . PMID 12552136 .
- ^ Tarver JE, Dos Reis M, Mirarab S, Moran RJ, Parker S, O'Reilly JE, et al. (Январь 2016). «Взаимосвязи плацентарных млекопитающих и пределы филогенетического вывода» . Биология и эволюция генома . 8 (2): 330–344. doi : 10.1093/gbe/evv261 . HDL : 1983/64D6E437-3320-480D-A16C-2E5B2E6B61D4 . PMC 4779606 . PMID 26733575 .
- ^ Alvarez-Carretero S, Tamuri Au, Battini M, Nascimento FF, Carlisle E, Asher RJ, Yang Z, Donoghue PC, et al. (2022). «График на уровне вида эволюции млекопитающих, интегрирующие филогеномные данные» . Природа . 602 (7896): 263–267. Bibcode : 2022natur.602..263a . doi : 10.1038/s41586-021-04341-1 . HDL : 1983/DE841853-D57B-40D9-876F-9BFCF7253F12 . PMID 34937052 . S2CID 245438816 .
- ^ Альварес-Трайд, Сандра; Тамури, Асиф; Баттини, Маттео; Рождение, Fabricia F.; Карлайл, Эмили; Ашер, Роберт; Ян, Зихенг; Donghue, Philip; Дос Рейс, Марио (2021). «Данные для временной шкалы на уровне видов эволюции млекопитающих, интегрирующих филогеномные данные» . Figshare . Doi : 10.6084/m9.figshare.14885691.v1 . Архивировано из оригинала 16 декабря 2023 года . Получено 11 ноября 2023 года .
- ^ Meng J, Wang Y, Li C (апрель 2011 г.). «Среднее ухо переходного млекопитающего от нового мелового Джехола Эутриконодонта». Природа . 472 (7342): 181–185. Bibcode : 2011natur.472..181m . doi : 10.1038/nature09921 . PMID 21490668 . S2CID 4428972 .
- ^ Альберг П.Е., Милнер А.Р. (апрель 1994 г.). «Происхождение и ранняя диверсификация тетрапод». Природа . 368 (6471): 507–514. Bibcode : 1994natur.368..507a . doi : 10.1038/368507a0 . S2CID 4369342 .
- ^ «Амниота - палеос» . Архивировано из оригинала 20 декабря 2010 года.
- ^ «Обзор Synapsida - Palaeos» . Архивировано из оригинала 20 декабря 2010 года.
- ^ Jump up to: а беременный в Кемп Т.С. (июль 2006 г.). «Происхождение и раннее излучение рептилий, подобных млекопитающим,: палеобиологическая гипотеза» (PDF) . Журнал эволюционной биологии . 19 (4): 1231–1247. doi : 10.1111/j.1420-9101.2005.01076.x . PMID 16780524 . S2CID 3184629 . Архивировано из оригинала (PDF) 8 марта 2021 года . Получено 14 января 2012 года .
- ^ Jump up to: а беременный Беннетт А.Ф., Рубен Дж.А. (1986). «Метаболический и терморегуляционный статус терапидов». В Hotton III N, MacLean JJ, Roth J, Roth EC (Eds.). Экология и биология рептилий, подобных млекопитающим . Вашингтон, округ Колумбия: Смитсоновская институциональная пресса. С. 207–218. ISBN 978-0-87474-524-5 .
- ^ Кермак Д.М., Кермак К.А. (1984). Эволюция персонажей млекопитающих . Вашингтон, округ Колумбия: Крюм Хелм. ISBN 978-0-7099-1534-8 Полем OCLC 10710687 .
- ^ Арауджо; и др. (28 июля 2022 г.). «Биомеханика внутреннего уха раскрывает позднее триасовое происхождение для эндотермии млекопитающих». Природа . 607 (7920): 726–731. Bibcode : 2022natur.607..726a . doi : 10.1038/s41586-022-04963-z . PMID 35859179 . S2CID 236245230 .
- ^ Таннер Л.Х., Лукас С.Г., Чепмен М.Г. (2004). «Оценка записи и причин позднего триасового вымирания» (PDF) . Земля-наука обзоров . 65 (1–2): 103–139. Bibcode : 2004esrv ... 65..103t . doi : 10.1016/s0012-8252 (03) 00082-5 . Архивировано из оригинала (PDF) 25 октября 2007 года.
- ^ Brusatte SL, Benton MJ, Ruta M, Lloyd GT (сентябрь 2008 г.). «Превосходство, конкуренция и оппортунизм в эволюционном радиации динозавров» (PDF) . Наука . 321 (5895): 1485–1488. Bibcode : 2008Sci ... 321.1485b . doi : 10.1126/science.1161833 . HDL : 20.500.11820/00556BAF-6575-44D9-AF39-BDD0B072AD2B . PMID 18787166 . S2CID 13393888 . Архивировано (PDF) от оригинала 19 июля 2018 года . Получено 12 октября 2019 года .
- ^ Gauthier JA (1986). «Сауришская монофилия и происхождение птиц». В Падие К (ред.). Происхождение птиц и эволюция полета. Мемуары Калифорнийской академии наук . Тол. 8. Сан -Франциско: Калифорнийская академия наук. С. 1–55.
- ^ Sereno PC (1991). «Базальные архозавры: филогенетические отношения и функциональные последствия». Мемуары Общества палеонтологии позвоночных . 2 : 1–53. doi : 10.2307/3889336 . JSTOR 3889336 .
- ^ Macleod N, Rawson PF, Forey PL, Banner FT, Boudagher-Fadel MK, Bouth PR, et al. (1997). «Биологический переход мелового нерва». Журнал геологического общества . 154 (2): 265–292. Bibcode : 1997jgsoc.154..265m . doi : 10.1144/gsjgs.154.2.0265 . S2CID 129654916 .
- ^ Hunt DM, Hankins MW, Collin SP, Marshall NJ (2014). Эволюция визуальных и не визуальных пигментов . Лондон: Спрингер. п. 73. ISBN 978-1-4614-4354-4 Полем OCLC 892735337 .
- ^ Бакалар Н. (2006). «Юрский» Бивер »найден; переписывает историю млекопитающих» . National Geographic News . Архивировано из оригинала 3 марта 2006 года . Получено 28 мая 2016 года .
- ^ Холл М.И., Камилар Дж. М., Кирк Е.К. (декабрь 2012 г.). «Форма глаз и ночное узкое место млекопитающих» . Труды Королевского общества B: Биологические науки . 279 (1749): 4962–4968. doi : 10.1098/rspb.2012.2258 . PMC 3497252 . PMID 23097513 .
- ^ LUO ZX (декабрь 2007 г.). «Трансформация и диверсификация в ранней эволюции млекопитающих». Природа . 450 (7172): 1011–1019. Bibcode : 2007natur.450.1011L . doi : 10.1038/nature06277 . PMID 18075580 . S2CID 4317817 .
- ^ Пикрелл Дж. (2003). «Самая старая сумчатая ископаемость, найденная в Китае» . National Geographic News. Архивировано из оригинала 17 декабря 2003 года . Получено 28 мая 2016 года .
- ^ Jump up to: а беременный Luo ZX, Yuan CX, Meng QJ, Ji Q (август 2011 г.). «Юрская эврианская млекопитающая и дивергенция сумчатых и плацент». Природа . 476 (7361): 442–5. Bibcode : 2011natur.476..442L . doi : 10.1038/nature10291 . PMID 21866158 . S2CID 205225806 .
- ^ Ji Q, Luo ZX, Yuan CX, Wible JR, Zhang JP, Georgi JA (апрель 2002 г.). «Самое раннее известное эврикское млекопитающее». Природа . 416 (6883): 816–822. Bibcode : 2002natur.416..816j . doi : 10.1038/416816a . PMID 11976675 . S2CID 4330626 .
- ^ Jump up to: а беременный Novacek MJ, Rougier GW, Wible JR, McKenna MC, Dashzeveg D, Horovitz I (октябрь 1997 г.). «Epipubic кости у эврианских млекопитающих от покойного мела Монголии». Природа . 389 (6650): 483–486. Bibcode : 1997natur.389..483n . doi : 10.1038/39020 . PMID 9333234 . S2CID 205026882 .
- ^ Power ML, Schulkin J (2012). «Эволюция живорождения у млекопитающих» . Эволюция человеческой плаценты . Балтимор: издательство Джона Хопкинса. п. 68. ISBN 978-1-4214-0643-5 .
- ^ Rowe T, Rich TH, Vickers-Rich P, Springer M, Woodburne Mo (январь 2008 г.). «Самый древний утконос и его подшипник на сроках дивергенции кладов утконос и эхидны» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (4): 1238–1242. BIBCODE : 2008PNAS..105.1238R . doi : 10.1073/pnas.0706385105 . PMC 2234122 . PMID 18216270 .
- ^ Грант Т (1995). «Воспроизведение» . Птлаток: уникальное млекопитающее . Сидней: Университет Нового Южного Уэльса. п. 55. ISBN 978-0-86840-143-0 Полем OCLC 33842474 .
- ^ Голдман как (июнь 2012 г.). «Эволюция иммунных функций молочной железы и защита младенца». Медицина для грудного вскармливания . 7 (3): 132–142. doi : 10.1089/bfm.2012.0025 . PMID 22577734 .
- ^ Jump up to: а беременный Роуз К.Д. (2006). Начало эпохи млекопитающих . Балтимор: издательство Джона Хопкинса. С. 82–83. ISBN 978-0-8018-8472-6 Полем OCLC 646769601 .
- ^ Бринк как (1955). «Исследование скелета Diademodon ». Palaeontologia Africana . 3 : 3–39.
- ^ Кемп Т.С. (1982). Млекопитающие рептилии и происхождение млекопитающих . Лондон: академическая пресса. п. 363. ISBN 978-0-12-404120-2 Полем OCLC 8613180 .
- ^ Estes R (1961). «Анатомия черепа рептилий Cynodont thrinaxodon liorhinus ». Бюллетень Музея сравнительной зоологии (1253): 165–180.
- ^ « Тринаксодон: новое млекопитающее» . National Geographic Daily News. 11 февраля 2009 года. Архивировано из оригинала 14 февраля 2009 года . Получено 26 августа 2012 года .
- ^ Jump up to: а беременный Bajdek P, Qvarnström M, Owocki K, Sulej T, Sennikov AG, Golubev VK, Niedźwiedzki G (2015). «Микробиота и остатки пищи, включая возможные доказательства волос до млекопитающих в верхних пермских копролитах из России». Летая . 49 (4): 455–477. doi : 10.1111/let.12156 .
- ^ Бота-Бринк Дж., Ангилчик К.Д. (2010). «Должны ли чрезвычайно высокие темпы роста в пермо-триассических дицинододонтах (Therapsida, Anomodontia), объясняют свой успех до и после окончания конечного пермического вымирания?» Полем Зоологический журнал Линневого общества . 160 (2): 341–365. doi : 10.1111/j.1096-3642.2009.00601.x .
- ^ Пол Г.С. (1988). Хищные динозавры мира . Нью -Йорк: Саймон и Шустер. п. 464 . ISBN 978-0-671-61946-6 Полем OCLC 18350868 .
- ^ Уотсон Дж. М., Грейвс Дж.А. (1988). «Монотримные клеточные циклы и эволюция гомеотермии». Австралийский журнал зоологии . 36 (5): 573–584. doi : 10.1071/ZO9880573 .
- ^ McNab BK (1980). «Энергетика и пределы умеренного распределения в армадилло». Журнал млекопитающих . 61 (4): 606–627. doi : 10.2307/1380307 . JSTOR 1380307 .
- ^ Kiilan-Jowwska Z, Huum JH (2006). "PDF): растяжение или PDF) . Acta Poolonological Польша 51 (3): 10237–10239. Archilded (PDF) с 25 января 25 2024января
- ^ Lillegraven JA, Kielan-Jaworowska Z, Clemens WA (1979). Мезозойские млекопитающие: первые две трети истории млекопитающих . Калифорнийский университет. п. 321. ISBN 978-0-520-03951-3 Полем OCLC 5910695 .
- ^ Oftedal OT (июль 2002 г.). «Молочная железа и ее происхождение во время эволюции синапсид». Журнал биологии молочной железы и неоплазии . 7 (3): 225–252. doi : 10.1023/a: 1022896515287 . PMID 12751889 . S2CID 25806501 .
- ^ Oftedal OT (июль 2002 г.). «Происхождение лактации как источник воды для яиц с пергаментом». Журнал биологии молочной железы и неоплазии . 7 (3): 253–266. doi : 10.1023/a: 1022848632125 . PMID 12751890 . S2CID 8319185 .
- ^ «Развитие груди» . Техасская детская больница . Архивировано из оригинала 13 января 2021 года . Получено 13 января 2021 года .
- ^ Пфафф, Катрин; Нагель, Дорис; Ганнелл, Грегг; Вебер, Герхард В.; Кривет, Юрген; Морло, Майкл; Бастл, Катхарина (2017). «Палеблогология Hyaenondon Exiiguus (Yyaenondonta, Mammalia) на основе морфометрического анализа костного лабиринта » Журнал анатомии 230 (2): 282–2 Doi : 10.1111/ joa.1 5244453PMC 27666133PMID
- ^ Jump up to: а беременный Sahney S, Benton MJ, Ferry PA (август 2010 г.). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и расширением позвоночных на земле» . Биологические письма . 6 (4): 544–547. doi : 10.1098/rsbl.2009.1024 . PMC 2936204 . PMID 20106856 .
- ^ Смит Ф.А., Бойер А.Г., Браун Дж.Х., Коста Д.П., Даян Т., Эрнест С.К. и др. (Ноябрь 2010). «Эволюция максимального размера тела земных млекопитающих». Наука . 330 (6008): 1216–1219. BIBCODE : 2010SCI ... 330.1216S . Citeseerx 10.1.1.383.8581 . doi : 10.1126/science.1194830 . PMID 21109666 . S2CID 17272200 .
- ^ Симмонс Н.Б., Сеймур К.Л., Хаберсетцер Дж., Гуннелл Г.Ф. (февраль 2008 г.). «Примитивная ранняя эоценовая летучая мышь из Вайоминга и эволюция полета и эхолокации» . Природа . 451 (7180): 818–821. Bibcode : 2008natur.451..818s . doi : 10.1038/nature06549 . HDL : 2027.42/62816 . PMID 18270539 . S2CID 4356708 . Архивировано из оригинала 25 января 2024 года . Получено 25 января 2024 года .
- ^ Bininda-Emonds OR, Cardillo M, Jones KE, Macpee RD, Beck RM, Grenyer R, et al. (Март 2007 г.). «Задержка роста современных млекопитающих» (PDF) . Природа . 446 (7135): 507–512. Bibcode : 2007natur.446..507b . doi : 10.1038/nature05634 . PMID 17392779 . S2CID 4314965 . Архивировано (PDF) из оригинала 25 января 2024 года . Получено 25 января 2024 года .
- ^ Jump up to: а беременный Wible JR, Rougier GW, Novacek MJ, Asher RJ (июнь 2007 г.). «Цикеты -эвхеристы и лоазинское происхождение для плацентарных млекопитающих возле границы K/T». Природа . 447 (7147): 1003–1006. Bibcode : 2007natur.447.1003w . doi : 10.1038/nature05854 . PMID 17581585 . S2CID 4334424 .
- ^ О'Лири М.А., Блох Джи, Флинн Дж.Дж., Гаудин Т.Дж., Гиалломбардо А., Джаннини Н.П. и др. (Февраль 2013 г.). «Плацентарный предок млекопитающих и радиация плацентов после K-PG» . Наука . 339 (6120): 662–667. Bibcode : 2013sci ... 339..662O . doi : 10.1126/science.1229237 . HDL : 11336/7302 . PMID 23393258 . S2CID 206544776 . Архивировано с оригинала 10 ноября 2021 года . Получено 30 июня 2022 года .
- ^ Hallide TJ, Upchurch P, Goswami A (февраль 2017 г.). «Устранение отношений палеоценовых плацентарных млекопитающих» . Биологические обзоры Кембриджского философского общества . 92 (1): 521–550. doi : 10.1111/brv.12242 . PMC 6849585 . PMID 28075073 .
- ^ Hallide TJ, Upchurch P, Goswami A (июнь 2016 г.). «Евтериане испытали повышенные эволюционные показатели сразу же после вымирания великого палеогена» . Разбирательство. Биологические науки . 283 (1833): 20153026. DOI : 10.1098/rspb.2015.3026 . PMC 4936024 . PMID 27358361 .
- ^ Jump up to: а беременный в Ni X, Gebo DL, Dagosto M, Meng J, Tafforeau P, Flynn JJ, Beard KC (июнь 2013 г.). «Самый старый известный скелет приматов и ранняя эволюция гаплорина». Природа . 498 (7452): 60–64. Bibcode : 2013natur.498 ... 60n . doi : 10.1038/nature12200 . PMID 23739424 . S2CID 4321956 .
- ^ Romer SA, Parsons TS (1977). Тело позвоночных . Филадельфия: Holt-Saunders International. С. 129–145. ISBN 978-0-03-910284-5 Полем OCLC 60007175 .
- ^ Purves WK, Sadava DE, Orian GH, Helle HC (2001). Жизнь: наука о биологии (6 -е изд.). Нью -Йорк: Sinauer Associates, Inc. p. 593. ISBN 978-0-7167-3873-2 Полем OCLC 874883911 .
- ^ Anthwal N, Joshi L, Tucker As (январь 2013 г.). «Эволюция среднего уха и челюсти млекопитающих: адаптация и новые структуры» . Журнал анатомии . 222 (1): 147–160. doi : 10.1111/j.1469-7580.2012.01526.x . PMC 3552421 . PMID 22686855 .
- ^ Ван Нивельт А.Ф., Смит К.К. (2005). «Чтобы заменить или не заменить: значение уменьшенной функциональной замены зубов у сумчатых и плацентарных млекопитающих». Палеобиология . 31 (2): 324–346. doi : 10.1666/0094-8373 (2005) 031 [0324: TRONTR] 2.0.CO; 2 . S2CID 37750062 .
- ^ Libertini G, Ferrara N (апрель 2016 г.). «Старение многолетних клеток и части органов в соответствии с запрограммированной парадигмой старения» . Возраст . 38 (2): 35. DOI : 10.1007/S11357-016-9895-0 . PMC 5005898 . PMID 26957493 .
- ^ Mao F, Wang Y, Meng J (2015). «Систематическое исследование микроструктуры зубной эмали Lambdopsalis Bulla (многотуркуляция, млекопитающие) - последствия для многотуркуляции биологии и филогения» . Plos один . 10 (5): E0128243. BIBCODE : 2015PLOSO..1028243M . doi : 10.1371/journal.pone.0128243 . PMC 4447277 . PMID 26020958 .
- ^ Osborn HF (1900). «Происхождение млекопитающих, III. Затылочные мыщелки рептильного трипробранного типа» . Американский натуралист . 34 (408): 943–947. doi : 10.1086/277821 . JSTOR 2453526 .
- ^ Crompton AW, Jenkins JR FA (1973). «Млекопитающие от рептилий: обзор происхождения млекопитающих». Ежегодный обзор земли и планетарных наук . 1 : 131–155. Bibcode : 1973areps ... 1..131c . doi : 10.1146/annurev.ea.01.050173.001023 .
- ^ Jump up to: а беременный Power ML, Schulkin J (2013). Эволюция человеческой плаценты . Балтимор: издательство Джона Хопкинса. С. 1890–1891. ISBN 978-1-4214-0643-5 Полем OCLC 940749490 .
- ^ Lindenfors P, Gittleman JL, Jones Ke (2007). «Диморфизм сексуального размера у млекопитающих». Пол, размер и гендерные роли: эволюционные исследования диморфизма сексуального размера . Оксфорд: издательство Оксфордского университета. С. 16–26. ISBN 978-0-19-920878-4 Полем Архивировано из оригинала 25 января 2024 года . Получено 25 января 2024 года .
- ^ Дирауф Л.А., Гулланд Ф.М. (2001). Справочник CRC по морской медицине млекопитающих: здоровье, болезнь и реабилитацию (2 -е изд.). Boca Raton: CRC Press. п. 154. ISBN 978-1-4200-4163-7 Полем OCLC 166505919 .
- ^ Луи Дж. Х., Хансен Д.В., Кригштейн А.Р. (июль 2011 г.). «Развитие и эволюция человеческого неокортекса» . Клетка . 146 (1): 18–36. doi : 10.1016/j.cell.2011.06.030 . PMC 3610574 . PMID 21729779 .
- ^ Килер CE (июнь 1933 г.). «Отсутствие мозолистого корпуса как менедлизационного персонажа в домашней мышью» . Труды Национальной академии наук Соединенных Штатов Америки . 19 (6): 609–611. Bibcode : 1933pnas ... 19..609k . doi : 10.1073/pnas.19.6.609 . JSTOR 86284 . PMC 1086100 . PMID 16587795 .
- ^ Standring S, Borley NR (2008). Анатомия Грея: анатомическая основа клинической практики (40 -е изд.). Лондон: Черчилль Ливингстон. С. 960–962. ISBN 978-0-8089-2371-8 Полем OCLC 213447727 .
- ^ Betts JF, Desaix P, Johnson E, Johnson JE, Korol O, Kruse D, et al. (2013). Анатомия и физиология . Хьюстон: издательство Райс Университет. С. 787–846. ISBN 978-1-938168-13-0 Полем OCLC 898069394 . Архивировано из оригинала 23 февраля 2022 года . Получено 25 января 2024 года .
- ^ Левицки М.Г. (2013). «Механика дыхания». Физиология легких (8 -е изд.). Нью-Йорк: McGraw-Hill Medical. ISBN 978-0-07-179313-1 Полем OCLC 940633137 .
- ^ Jump up to: а беременный Умеш К.Б. (2011). «Легочная анатомия и физиология» . Справочник по механической вентиляции . Нью -Дели: Медицинская публикация Jaypee Brothers. п. 12. ISBN 978-93-80704-74-6 Полем OCLC 945076700 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Feldhamer GA, Drickamer LC, Vessey SH, Merritt JF, Krajewski C (2007). МАММАЛОГИ: Адаптация, разнообразие, экология (3 -е изд.). Балтимор: издательство Джона Хопкинса. ISBN 978-0-8018-8695-9 Полем OCLC 124031907 .
- ^ Tinker SW (1988). Киты мира . Брилл Архив. п. 51. ISBN 978-0-935848-47-2 .
- ^ Ромер как (1959). История позвоночных (4 -е изд.). Чикаго: Университет Чикагской Прессы. ISBN 978-0-226-72490-4 .
- ^ De Muizon C, Lange-Badré B (1997). «Плотоядные зубные адаптация у трибосфенических млекопитающих и филогенетической реконструкции». Летая . 30 (4): 353–366. Bibcode : 1997Letha..30..353d . doi : 10.1111/j.1502-3931.1997.tb00481.x .
- ^ Jump up to: а беременный Лангер P (июль 1984 г.). «Сравнительная анатомия желудка у травоядных млекопитающих». Ежеквартальный журнал экспериментальной физиологии . 69 (3): 615–625. doi : 10.1113/expphysiol.1984.sp002848 . PMID 6473699 . S2CID 30816018 .
- ^ Воган Т.А., Райан Дж. М., Чеплвски Нью -Джерси (2011). "Perissodactyla " МАММАЛОГИ (5 -е изд.). Джонс и Бартлетт. П. 322. ISBN 978-0-7637-6299-5 Полем OCLC 437300511 .
- ^ Flower WH, Lydekker R (1946). Введение в изучение млекопитающих, живущих и вымерших . Лондон: Адам и Чарльз Блэк. п. 496. ISBN 978-1-110-76857-8 .
- ^ Sreekumar S (2010). Основная физиология . PHI Learning Pvt. ООО с. 180–181. ISBN 978-81-203-4107-4 .
- ^ Cheifetz As (2010). Оксфордский американский справочник гастроэнтерологии и гепатологии . Оксфорд: издательство Оксфордского университета, США. п. 165. ISBN 978-0-19-983012-1 .
- ^ Кунц Е. (2008). Гепатология: учебник и атлас . Германия: Спрингер. п. 38. ISBN 978-3-540-76838-8 .
- ^ Ортис Р.М. (июнь 2001 г.). «Осморгуляция у морских млекопитающих» . Журнал экспериментальной биологии . 204 (Pt 11): 1831–1844. doi : 10.1242/jeb.204.11.1831 . PMID 11441026 . Архивировано из оригинала 25 января 2024 года . Получено 25 января 2024 года .
- ^ Jump up to: а беременный в дюймовый Роман А.С., Парсонс Т.С. (1977). Тело позвоночных . Филадельфия: Holt-Saunders International. С. 396–399. ISBN 978-0-03-910284-5 .
- ^ Линзи, Дональд В. (2020). Биология позвоночных: систематика, таксономия, естественная история и сохранение . Johns Hopkins University Press. п. 306. ISBN 978-1-42143-733-0 Полем Архивировано из оригинала 22 января 2024 года . Получено 22 января 2024 года .
- ^ Symonds, Matthew Re (февраль 2005 г.). «Биологические обзоры - журналы Cambridge» . Биологические обзоры . 80 (1): 93–128. doi : 10.1017/s1464793104006566 . PMID 15727040 . Архивировано с оригинала 22 ноября 2015 года . Получено 21 января 2017 года .
- ^ Докинз Р., Вонг Ю. (2016). Сказка предка: паломничество на рассвет эволюции (2 -е изд.). Бостон: моряк книги. п. 281. ISBN 978-0-544-85993-7 .
- ^ Jump up to: а беременный в Fitch WT (2006). «Производство вокализаций у млекопитающих» (PDF) . В коричневом k (ред.). Энциклопедия языка и лингвистики . Оксфорд: Elsevier. С. 115–121. Архивировано из оригинала 1 июня 2024 года . Получено 25 января 2024 года .
- ^ Langevin P, Barclay RM (1990). « Hypsignathus monstrosus » . Виды млекопитающих (357): 1–4. doi : 10.2307/3504110 . JSTOR 3504110 .
- ^ Weissengruber GE, Forstenpointner G, Peters G, Kübber-Heiss A, Fitch WT (сентябрь 2002 г.). «Хиоидный аппарат и глотка в левке (пантера Лео), Ягуар (пантера Онка), Тигр (Пантера Тигрис), Гепарда (ацинонихджубатус) и домашний кот (Фелис Сильвестрис ф. Catus)» . Журнал анатомии . 201 (3): 195–209. doi : 10.1046/j.1469-7580.2002.00088.x . PMC 1570911 . PMID 12363272 .
- ^ Stoeger AS, Heilmann G, Zeppelzauer M, Ganswindt A, Hensman S, Charlton BD (2012). «Визуализация звуковой эмиссии вокализаций слонов: доказательство двух типов производства Rumble» . Plos один . 7 (11): E48907. BIBCODE : 2012PLOSO ... 748907S . doi : 10.1371/journal.pone.0048907 . PMC 3498347 . PMID 23155427 .
- ^ Кларк CW (2004). «BALEEN WHALE Infrasonic Sounds: естественная изменчивость и функция». Журнал Акустического общества Америки . 115 (5): 2554. Bibcode : 2004saj..115.2554c . doi : 10.1121/1,4783845 .
- ^ Jump up to: а беременный Доусон Т.Дж., Вебстер К.Н., Малони С.К. (февраль 2014 г.). «Мех млекопитающих в открытых средах; обязательно ли Crypsis и тепловые потребности конфликтуют? По сравнению с белым медведем и сумчатым коалой». Журнал сравнительной физиологии б . 184 (2): 273–284. doi : 10.1007/s00360-013-0794-8 . PMID 24366474 . S2CID 9481486 .
- ^ Сломенский А., Тобин Д.Дж., Шибахара С., Вортсман Дж (октябрь 2004 г.). «Пигментация меланина у кожи млекопитающих и ее гормональная регуляция». Физиологические обзоры . 84 (4): 1155–1228. doi : 10.1152/physrev.00044.2003 . PMID 15383650 . S2CID 21168932 .
- ^ Хилтон -младший Б. (1996). «Дикая природа Южной Каролины» . Цвета животных . 43 (4). Хилтон -пруд Центр: 10–15. Архивировано из оригинала 25 января 2024 года . Получено 26 ноября 2011 года .
- ^ Jump up to: а беременный Prum Ro, Torres RH (май 2004 г.). «Структурная окраска кожи млекопитающих: сходящаяся эволюция когерентно рассеяния массивов кожных коллагенов» (PDF) . Журнал экспериментальной биологии . 207 (Pt 12): 2157–2172. doi : 10.1242/jeb.00989 . HDL : 1808/1599 . PMID 15143148 . S2CID 8268610 . Архивировано (PDF) из оригинала 5 июня 2024 года . Получено 25 января 2024 года .
- ^ Suutari M, Majaneva M, Mese Dp, Voirin B, Aiello A, Friedl T, et al. (Март 2010 г.). «Молекулярные доказательства разнообразного сообщества зеленых водорослей, растущей в волосах ленивцев, и конкретной ассоциации с Trichophilus welckeri (Chlorophyta, Ulvophyceae)» . BMC Эволюционная биология . 10 (86): 86. Bibcode : 2010bmcee..10 ... 86 с . doi : 10.1186/1471-2148-10-86 . PMC 2858742 . PMID 20353556 .
- ^ Каро Т (2005). «Адаптивное значение окраски у млекопитающих» . Биоссака . 55 (2): 125–136. doi : 10.1641/0006-3568 (2005) 055 [0125: tasoci] 2.0.co; 2 .
- ^ Mills LS, Zimova M, Oyler J, Runge S, Abatzoglou JT, Lukacs PM (апрель 2013 г.). «Камуфляжное несоответствие в сезонном цвете пальто из -за снижения продолжительности снега» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (18): 7360–7365. Bibcode : 2013pnas..110.7360M . doi : 10.1073/pnas.1222724110 . PMC 3645584 . PMID 23589881 .
- ^ Каро Т (февраль 2009 г.). «Контрастная окраска у наземных млекопитающих» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 364 (1516): 537–548. doi : 10.1098/rstb.2008.0221 . PMC 2674080 . PMID 18990666 .
- ^ Plavcan JM (2001). «Сексуальный диморфизм в эволюции приматов» . Американский журнал физической антропологии . Suppl 33 (33): 25–53. doi : 10.1002/ajpa.10011 . PMID 11786990 . S2CID 31722173 .
- ^ Брэдли Б.Дж., Джеральд М.С., Виддиг А., Манди Н.И. (2012). «Изменение цветового цвета и экспрессия гена пигментации в макаках -резусах ( Macaca Mulatta )» (PDF) . Журнал эволюции млекопитающих . 20 (3): 263–270. doi : 10.1007/s10914-012-9212-3 . S2CID 13916535 . Архивировано из оригинала (PDF) 24 сентября 2015 года.
- ^ Caro T , Izzo A, Riner RC, Walker H, Stankowich T (апрель 2014 г.). «Функция полос зебры» . Природная связь . 5 : 3535. Bibcode : 2014natco ... 5.3535c . doi : 10.1038/ncomms4535 . PMID 24691390 . S2CID 9849814 .
- ^ Нагиб, Марк (19 апреля 2020 года). Достижения в изучении поведения . Академическая пресса. ISBN 978-0-12-820726-0 .
- ^ Kobayashi K, Kitano T, Iwao Y, Kondo M (2018). Репродуктивные и стратегии развития: преемственность жизни . Спрингер. п. 290. ISBN 978-4-431-56609-0 .
- ^ Jump up to: а беременный Ломбарди Дж. (1998). Сравнительное воспроизведение позвоночных . Springer Science & Business Media. ISBN 978-0-7923-8336-9 .
- ^ Либби Генриетта Хайман (15 сентября 1992 г.). Сравнительная анатомия Хаймана . Университет Чикагской Прессы. С. 583–. ISBN 978-0-226-87013-7 .
- ^ Нагиб, Марк (19 апреля 2020 года). Достижения в изучении поведения . Академическая пресса. ISBN 978-0-12-820726-0 .
- ^ Tyndale-Biscoe H, Renfree M (1987). Репродуктивная физиология сумчатых . Издательство Кембриджского университета. ISBN 978-0-521-33792-2 .
- ^ Johnston SD, Smith B, Pyne M, Stenzel D, Holt WV (2007). «Односторонняя эякуляция пучков спермы эхидны» (PDF) . Американский натуралист . 170 (6): E162 - E164. doi : 10.1086/522847 . PMID 18171162 . S2CID 40632746 .
- ^ Бача -младший, Уильям Дж.; Бача, Линда М. (2012). Цветный атлас ветеринарной гистологии . Уайли. п. 308. ISBN 978-1-11824-364-0 Полем Получено 28 ноября 2023 года .
- ^ Кук, Фред; Брюс, Дженни (2004). Энциклопедия животных: полное визуальное руководство . Калифорнийский университет. п. 79. ISBN 978-0-52024-406-1 Полем Получено 28 ноября 2023 года .
- ^ Максвелл К.Е. (2013). Секс -императив: эволюционная история о сексуальном выживании . Спрингер. С. 112–113. ISBN 978-1-4899-5988-1 .
- ^ Воган Т.А., Ян Дж.П., Чеплвски Нью -Джерси (2011). Млекопитающая Jones & Barttt Publishers. П. 387. ISBN 978-0-03-025034-7 .
- ^ Jump up to: а беременный Hoffman EA, Rowe TB (сентябрь 2018 г.). «Перинаты юрских стволовых млекопитающих и происхождение размножения и роста млекопитающих». Природа . 561 (7721): 104–108. Bibcode : 2018natur.561..104H . doi : 10.1038/s41586-018-0441-3 . PMID 30158701 . S2CID 205570021 .
- ^ Wallis MC, Waters PD, Delbridge ML, Kirby PJ, Pask AJ, Grützner F, et al. (2007). «Определение пола у утконос и эхидны: аутосомное местоположение SOX3 подтверждает отсутствие SRY в монотримах». Исследование хромосом . 15 (8): 949–959. doi : 10.1007/s10577-007-1185-3 . PMID 18185981 . S2CID 812974 .
- ^ Маршалл Грейвс JA (2008). «Странные геномы животных и эволюция сексуальных и половых хромосом позвоночных» (PDF) . Ежегодный обзор генетики . 42 : 565–586. doi : 10.1146/annurev.genet.42.110807.091714 . PMID 18983263 . Архивировано из оригинала (PDF) 4 сентября 2012 года . Получено 25 января 2024 года .
- ^ Салли М (2005). «Поведение млекопитающих и образ жизни» . Млекопитающие . Чикаго: Рейнтри. п. 6. ISBN 978-1-4109-1050-9 Полем OCLC 53476660 .
- ^ Верма П.С., Пандей Б.П. (2013). ISC Biology Book I для класса XI . Нью -Дели: С. Чанд и Компания. п. 288. ISBN 978-81-219-2557-0 .
- ^ Oftedal OT (июль 2002 г.). «Молочная железа и ее происхождение во время эволюции синапсид». Журнал биологии молочной железы и неоплазии . 7 (3): 225–252. doi : 10.1023/a: 1022896515287 . PMID 12751889 . S2CID 25806501 .
- ^ Krockenberger A (2006). «Лактация». В Dickman CR, Armati PJ, Hume Id (ред.). Сумчатые . Издательство Кембриджского университета. п. 109. ISBN 978-1-139-45742-2 .
- ^ Schulkin J, Power ML (2016). Молоко: биология лактации . Johns Hopkins University Press. п. 66. ISBN 978-1-4214-2042-4 .
- ^ Томпсон К.В., Бейкер А.Дж., Бейкер А.М. (2010). «Отцовская забота и поведенческое развитие у пленных млекопитающих». В Kleiman DG, Thompson KV, Baer CK (Eds.). Дикие млекопитающие в принципах и методах управления зоопарком (2 -е изд.). Университет Чикагской Прессы. п. 374. ISBN 978-0-226-44011-8 .
- ^ NA, восстановимый NA Recele of JB (2002). Биология (6 -е изд.). Бенджамин Каммингс. п. 845 ISBN 978-0-8053-6624-2 Полем OCLC 47521441 .
- ^ Баффенштейн Р., Яхав С. (1991). «Является ли голая глабер с голым человеком-ратом Hererocephalus эндотермическим, но Poikilothermic млекопитающим?». Журнал термической биологии . 16 (4): 227–232. Bibcode : 1991jtbio..16..227b . doi : 10.1016/0306-4565 (91) 90030-6 .
- ^ Шмидт-Нильсен К., Дьюк Дж.Б. (1997). «Температурные эффекты» . Физиология животных: адаптация и окружающая среда (5 -е изд.). Кембридж: издательство Кембриджского университета. п. 218. ISBN 978-0-521-57098-5 Полем OCLC 35744403 .
- ^ Jump up to: а беременный Lorenzini A, Johnson FB, Oliver A, Tresini M, Smith JS, Hdeib M, et al. (2009). «Значительная корреляция долговечности видов с распознаванием ДНК двойной цепи, но не с длиной теломер» . Механизмы старения и развития . 130 (11–12): 784–792. doi : 10.1016/j.mad.2009.10.004 . PMC 2799038 . PMID 19896964 .
- ^ Hart RW, Setlow RB (июнь 1974 г.). «Корреляция между дезоксирибонуклеиновой кислотой эксцизионной репрессией и жизненно важным промежутком у ряда видов млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (6): 2169–2173. Bibcode : 1974pnas ... 71.2169h . doi : 10.1073/pnas.71.6.2169 . PMC 388412 . PMID 4526202 .
- ^ Ma S, Upneja A, Galecki A, Tsai YM, Burant CF, Raskind S, et al. (Ноябрь 2016). «Профилирование на основе клеточной культуры между млекопитающими выявляет восстановление ДНК и метаболизм в качестве детерминантов долговечности видов» . элиф . 5 doi : 10.7554/elife.19130 . PMC 5148604 . PMID 27874830 .
- ^ Grube K, Bürkle A (декабрь 1992 г.). «Поли (ADP-рибоза) полимеразная активность в мононуклеарных лейкоцитах 13 видов млекопитающих коррелирует с видоспецифичной жизненной силой» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (24): 11759–11763. Bibcode : 1992pnas ... 8911759G . doi : 10.1073/pnas.89.24.11759 . PMC 50636 . PMID 1465394 .
- ^ Фрэнсис А.А., Ли В.Х., Риган Д.Д. (июнь 1981 г.). «Связь восстановления удаления ДНК ультрафиолетовых индуцированных поражений с максимальной продолжительностью жизни млекопитающих». Механизмы старения и развития . 16 (2): 181–189. doi : 10.1016/0047-6374 (81) 90094-4 . PMID 7266079 . S2CID 19830165 .
- ^ Treton JA, Courtois Y (март 1982 г.). «Корреляция между репарацией удаления ДНК и продолжительностью жизни млекопитающих в эпителиальных клетках линзы». Cell Biology International Reports . 6 (3): 253–260. doi : 10.1016/0309-1651 (82) 90077-7 (неактивный 2 сентября 2024 г.). PMID 7060140 .
{{cite journal}}: CS1 Maint: doi неактивен по состоянию на сентябрь 2024 года ( ссылка ) - ^ Maslansky CJ, Williams GM (февраль 1985 г.). «Ультрафиолетовый индуцированный светом синтез репарации ДНК в гепатоцитах из видов разных долговечностей». Механизмы старения и развития . 29 (2): 191–203. doi : 10.1016/0047-6374 (85) 90018-1 . PMID 3974310 . S2CID 23988416 .
- ^ Уокер В.Ф., Хомбергер Д.Г. (1998). Анатомия и рассечение плода свиньи (5 -е изд.). Нью -Йорк: WH Freeman and Company. п. 3. ISBN 978-0-7167-2637-1 Полем OCLC 40576267 .
- ^ ORR CM (ноябрь 2005 г.). «Антиатр, бьющий с суставами: тест с конвергенцией адаптации для предполагаемых чертовых особенностей африканских hominidae». Американский журнал физической антропологии . 128 (3): 639–658. doi : 10.1002/ajpa.20192 . PMID 15861420 .
- ^ Fish FE, Frappell PB, Baudinette RV, Macfarlane PM (февраль 2001 г.). «Энергетика наземного локомоции утконос Ornithorhynchus anatinus» (PDF) . Журнал экспериментальной биологии . 204 (Pt 4): 797–803. doi : 10.1242/jeb.204.4.797 . HDL : 2440/12192 . PMID 11171362 . Архивировано (PDF) из оригинала 14 марта 2024 года . Получено 25 января 2024 года .
- ^ Dhingra P (2004). «Сравнительный двуночный - как остальная часть животного царства ходит по двум ногам» . Антропологическая наука . 131 (231). Архивировано из оригинала 21 апреля 2021 года . Получено 11 марта 2017 года .
- ^ Александр Р.М. (май 2004 г.). «Двухвосткие животные и их отличия от людей» . Журнал анатомии . 204 (5): 321–330. doi : 10.1111/j.0021-8782.2004.00289.x . PMC 1571302 . PMID 15198697 .
- ^ Jump up to: а беременный Dagg AI (1973). «Походки у млекопитающих». Обзор млекопитающих . 3 (4): 135–154. doi : 10.1111/j.1365-2907.1973.tb00179.x .
- ^ Робертс Т.Д. (1995). Понимание баланса: механика осанки и передвижения . Сан -Диего: Нельсон Торнес. п. 211. ISBN 978-1-56593-416-0 Полем OCLC 33167785 .
- ^ Jump up to: а беременный в Cartmill M (1985). "Подъем". В Hildebrand M, Bramble DM, Liem KF, Wake DB (Eds.). Функциональная морфология позвоночных . Кембридж: Belknap Press. С. 73–88. ISBN 978-0-674-32775-7 Полем OCLC 11114191 .
- ^ Vernes K (2001). «Скоростная производительность северной летающей белки ( Glaucomys Sabrinus ) в зрелом смешанном лесу восточной Канады» . Журнал млекопитающих . 82 (4): 1026–1033. doi : 10.1644/1545-1542 (2001) 082 <1026: gpotnf> 2,0.co; 2 . S2CID 78090049 .
- ^ Барба Ла (октябрь 2011 г.). «Летучие мыши - единственные летающие млекопитающие» . Биоэриальная локомоция . Архивировано из оригинала 14 мая 2016 года . Получено 20 мая 2016 года .
- ^ «Летучие мыши в полете раскрывают неожиданную аэродинамику» . Scienceday . 2007. Архивировано из оригинала 19 декабря 2019 года . Получено 12 июля 2016 года .
- ^ Hedenström A, Johansson LC (март 2015 г.). «BAT Flight: аэродинамика, кинематика и морфология полета» (PDF) . Журнал экспериментальной биологии . 218 (Pt 5): 653–663. doi : 10.1242/jeb.031203 . PMID 25740899 . S2CID 21295393 . Архивировано (PDF) из оригинала 25 января 2024 года . Получено 25 января 2024 года .
- ^ «Летучие летучие мыши экономят энергию, рисуя крылья наверх» . Scienceday . 2012. Архивировано из оригинала 31 мая 2021 года . Получено 12 июля 2016 года .
- ^ Карен Т (2008). Вися с летучими мышами: экобаты, вампиры и кинозвезды . Альбукерке: Университет Нью -Мексико Пресс. п. 14. ISBN 978-0-8263-4403-8 Полем OCLC 191258477 .
- ^ Sterbing-D'Angelo S, Chadha M, Chiu C, Falk B, Xian W, Barcelo J, et al. (Июль 2011). «Датчики летучей мыши поддерживают управление полетом» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (27): 11291–11296. BIBCODE : 2011PNAS..10811291S . doi : 10.1073/pnas.1018740108 . PMC 3131348 . PMID 21690408 .
- ^ Damiani, R, 2003, самые ранние доказательства Cynodont Burrowing, The Royal Society Publishing, том 270, выпуск 1525
- ^ Шимер HW (1903). «Адаптация к водным, древесным, исчезающим и курсовым привычкам у млекопитающих. III. Фоссориальные адаптации» . Американский натуралист . 37 (444): 819–825. doi : 10.1086/278368 . JSTOR 2455381 . S2CID 83519668 . Архивировано из оригинала 9 апреля 2023 года . Получено 23 августа 2020 года .
- ^ Stanhope MJ, Waddell VG, Madsen O, De Jong W, Hedges SB, Cleven GC, et al. (Август 1998). «Молекулярные доказательства многочисленного происхождения насекомого и нового порядка эндемичных африканских насекомых млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (17): 9967–9972. Bibcode : 1998pnas ... 95.9967S . doi : 10.1073/pnas.95.17.9967 . PMC 21445 . PMID 9707584 .
- ^ Перри Да (1949). «Анатомическая основа плавания в китах». Журнал зоологии . 119 (1): 49–60. doi : 10.1111/j.1096-3642.1949.tb00866.x .
- ^ Fish Fe, Hui CA (1991). «Плавание дельфина - обзор» (PDF) . Обзор млекопитающих . 21 (4): 181–195. doi : 10.1111/j.1365-2907.1991.tb00292.x . Архивировано из оригинала (PDF) 29 августа 2006 года.
- ^ Marsh H (1989). «Глава 57: Dugongidae» (PDF) . Фауна Австралии . Тол. 1. Канберра: правительственные публикации Австралии. ISBN 978-0-644-06056-1 Полем OCLC 27492815 . Архивировано из оригинала (PDF) 11 мая 2013 года.
- ^ Jump up to: а беременный Берта А (2012). «Исправленное разнообразие: эволюция и адаптация». Возвращение в море: жизнь и эволюционные времена морских млекопитающих . Калифорнийский университет. С. 62–64. ISBN 978-0-520-27057-2 .
- ^ Jump up to: а беременный Fish Fe, Hurley J, Costa DP (февраль 2003 г.). «Маневренность от морского льва Zalophus californianus: поворот производительности нестабильного дизайна тела» . Журнал экспериментальной биологии . 206 (Pt 4): 667–674. doi : 10.1242/jeb.00144 . PMID 12517984 .
- ^ Jump up to: а беременный Ридман М. (1990). Черносы: Печать, морские львы и морские морски . Калифорнийский университет. ISBN 978-0-520-06497-3 Полем OCLC 19511610 .
- ^ Fish FE (1996). «Переходы от тяги на основе перетаскивания к движению на основе подъема в плавании млекопитающих» . Интегративная и сравнительная биология . 36 (6): 628–641. doi : 10.1093/icb/36.6.628 .
- ^ Fish Fe (2000). «Биомеханика и энергетика у водных и полуакватических млекопитающих: утконос к киту» (PDF) . Физиологическая и биохимическая зоология . 73 (6): 683–698. Citeseerx 10.1.1.734.1217 . doi : 10.1086/318108 . PMID 11121343 . S2CID 49732160 . Архивировано из оригинала (PDF) 4 августа 2016 года.
- ^ Eltringham SK (1999). «Анатомия и физиология». Бегемоты . Лондон: T & Ad Poyser Ltd. p. 8. ISBN 978-0-85661-131-5 Полем OCLC 42274422 .
- ^ «Бегемопотамус Hippopotamus amphibius » . National Geographic . Архивировано с оригинала 25 ноября 2014 года . Получено 30 апреля 2016 года .
- ^ Jump up to: а беременный Сейфарт Р.М., Чейни Д.Л., Марлер П. (1980). «Аварийные звонки Vervet Monkey: семантическое общение в бесплатном примате» . Поведение животных . 28 (4): 1070–1094. doi : 10.1016/s0003-3472 (80) 80097-2 . S2CID 53165940 . Архивировано с оригинала 12 сентября 2019 года . Получено 22 сентября 2018 года .
- ^ Zuberbühler K (2001). «Специфичные для хищника звонки в обезьянах Кэмпбелла, Cercopithecus Campbelli ». Поведенческая экология и социобиология . 50 (5): 414–442. Bibcode : 2001becos..50..414z . doi : 10.1007/s002650100383 . JSTOR 4601985 . S2CID 21374702 .
- ^ Slabbekoorn H, Smith TB (апрель 2002 г.). «Песня птица, экология и видообразование» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 357 (1420): 493–503. doi : 10.1098/rstb.2001.1056 . PMC 1692962 . PMID 12028787 .
- ^ Bannister JL (2008). «Бальные киты (мистицеты)» . В Perrin WF, Würsig B, TheWissen JG (Eds.). Энциклопедия морских млекопитающих (2 -е изд.). Академическая пресса. С. 80–89. ISBN 978-0-12-373553-9 .
- ^ Скотт Н. (2002). «Существа культуры? Обозначение культурных систем в китах и дельфинах» . Биоссака . 52 (1): 9–14. doi : 10.1641/0006-3568 (2002) 052 [0009: cocmtc] 2.0.co; 2 . S2CID 86121405 .
- ^ Boughman JW (февраль 1998 г.). «Вокальное обучение большим копьями летучими мышами» . Разбирательство. Биологические науки . 265 (1392): 227–233. doi : 10.1098/rspb.1998.0286 . PMC 1688873 . PMID 9493408 .
- ^ «Язык Prairie Dogs декодирован учеными» . CBC News. 21 июня 2013 года. Архивировано с оригинала 22 мая 2015 года . Получено 20 мая 2015 года .
- ^ Mayell H (3 марта 2004 г.). «Слоны называют большие расстояния в час» . National Geographic . Архивировано из оригинала 5 марта 2004 года . Получено 15 ноября 2016 года .
- ^ Смит Дж. М. , Харпер Д. (2003). Сигналы животных . Оксфордская серия по экологии и эволюции. Издательство Оксфордского университета. С. 61–63. ISBN 978-0-19-852684-1 Полем OCLC 54460090 .
- ^ Fitzgibbon CD, Fanshawe JH (1988). «Стоимость в газеле Томсона: честный сигнал состояния» (PDF) . Поведенческая экология и социобиология . 23 (2): 69–74. Bibcode : 1988becos..23 ... 69f . doi : 10.1007/bf00299889 . S2CID 2809268 . Архивировано из оригинала (PDF) 25 февраля 2014 года.
- ^ Bildstein KL (май 1983). «Почему белохвостые оленя помечают их хвосты». Американский натуралист . 121 (5): 709–715. doi : 10.1086/284096 . JSTOR 2460873 . S2CID 83504795 .
- ^ Гослинг Л.М. (январь 1982). «Переоценка функции маркировки запаха на территориях» . Zeitschrift für Tierpsychologie . 60 (2): 89–118. doi : 10.1111/j.1439-0310.1982.tb00492.x . Архивировано (PDF) из оригинала 27 марта 2018 года . Получено 12 октября 2019 года .
- ^ Zala SM, Potts WK, Penn DJ (март 2004 г.). «Эксплуаты по маркировке ароматов обеспечивают честные сигналы здоровья и инфекции». Поведенческая экология . 15 (2): 338–344. doi : 10.1093/beheco/arh022 . HDL : 10.1093/beheco/arh022 .
- ^ Джонсон Р.П. (август 1973 г.). «Маркировка запаха у млекопитающих». Поведение животных . 21 (3): 521–535. doi : 10.1016/s0003-3472 (73) 80012-0 .
- ^ Schevill WE, McBride AF (1956). «Доказательства эхолокации китообразными». Глубоководные исследования . 3 (2): 153–154. Bibcode : 1956dsr ..... 3..153s . doi : 10.1016/0146-6313 (56) 90096-X .
- ^ Уилсон В., Мосс С. (2004). Томас Дж. (Ред.). Эхолокация в летучих мышах и дельфинах . Чикагское университетское издательство. п. 22. ISBN 978-0-226-79599-7 Полем OCLC 50143737 .
- ^ AU WW (1993). Сонар дельфинов . Springer-Verlag. ISBN 978-3-540-97835-0 Полем OCLC 26158593 .
- ^ Сандерс Дж.Г., Бейхман А.С., Роман Дж., Скотт Дж.Дж., Эмерсон Д., Маккарти Дж.Дж., Гирги П.Р. (сентябрь 2015 г.). «Bleen Whales организуют уникальный кишечный микробиом с сходством как с плотоядными животными, так и с травоядными» . Природная связь . 6 : 8285. Bibcode : 2015natco ... 6.8285s . doi : 10.1038/ncomms9285 . PMC 4595633 . PMID 26393325 .
- ^ Speaksman Jr (1996). «Энергетика и эволюция размера тела у маленьких наземных млекопитающих» (PDF) . Симпозии зоологического общества Лондона (69): 69–81. Архивировано из оригинала (PDF) 2 июня 2021 года . Получено 31 мая 2016 года .
- ^ Jump up to: а беременный Уилсон Д.Е., Берни Д., ред. (2001). Животное: окончательное визуальное руководство по дикой природе мира . DK Publishing. С. 86–89. ISBN 978-0-7894-7764-4 Полем OCLC 46422124 .
- ^ Jump up to: а беременный Ван Валкенбург B (июль 2007 г.). «Deja Vu: эволюция кормления морфологий в плотоядных животных» . Интегративная и сравнительная биология . 47 (1): 147–163. doi : 10.1093/ICB/ICM016 . PMID 21672827 .
- ^ Sacco T, Van Valkenburgh B (2004). «Экоморфологические индикаторы кормления у медведей (Carnivora: Ursidae)». Журнал зоологии . 263 (1): 41–54. doi : 10.1017/s0952836904004856 .
- ^ Singer MS, Bernays EA (2003). «Понимание всеявенности нуждается в поведенческой перспективе» . Экология . 84 (10): 2532–2537. Bibcode : 2003ecol ... 84.2532S . doi : 10.1890/02-0397 .
- ^ Hutson JM, Burke CC, Haynes G (1 декабря 2013 г.). «Остеофагия и модификации костей жирафом и другими большими копылами». Журнал археологической науки . 40 (12): 4139–4149. Bibcode : 2013Jarsc..40.4139h . doi : 10.1016/j.jas.2013.06.004 .
- ^ "Почему кошки едят траву?" Полем Pet Md. Архивировано с оригинала 10 декабря 2016 года . Получено 13 января 2017 года .
- ^ Geiser F (2004). «Скорость метаболизма и снижение температуры тела во время спячки и ежедневного оцепенения». Ежегодный обзор физиологии . 66 : 239–274. doi : 10.1146/annurev.physiol.66.032102.115105 . PMID 14977403 . S2CID 22397415 .
- ^ Хамфрис М.М., Томас Д.В., Крамер Д.Л. (2003). «Роль доступности энергии в спячке млекопитающих: подход затрат и выгод». Физиологическая и биохимическая зоология . 76 (2): 165–179. doi : 10.1086/367950 . PMID 12794670 . S2CID 14675451 .
- ^ Барнс Б.М. (июнь 1989 г.). «Избегание замораживания у млекопитающего: температура тела ниже 0 градуса C в арктическом спящем спящем состоянии». Наука . 244 (4912): 1593–1595. Bibcode : 1989sci ... 244.1593b . doi : 10.1126/science.2740905 . PMID 2740905 .
- ^ Фриц Г. (2010). «Астивация у млекопитающих и птиц» . В Навасе, Калифорния, Карвалью JE (ред.). Астивация: молекулярные и физиологические аспекты . Прогресс в молекулярной и субклеточной биологии. Тол. 49. Springer-Verlag. С. 95–113. doi : 10.1007/978-3-642-02421-4 . ISBN 978-3-642-02420-7 .
- ^ Майер, с. 59
- ^ Grove JC, Grey La, La Santa Medina N, Sivakumar N, Ahn JS, Corpuz TV, Berke JD, Kreitzer AC, Knight Za (июль 2022 г.). «Подсистемы дофамина, которые отслеживают внутренние состояния» . Природа . 608 (7922): 374–380. Bibcode : 2022natur.608..374G . doi : 10.1038/s41586-022-04954-0 . PMC 9365689 . PMID 35831501 .
- ^ Jump up to: а беременный в дюймовый и Метла, с. 105
- ^ Смит, с. 238.
- ^ «Языки кошек используют сложную физику» . 12 ноября 2010 г.
- ^ Смит, с. 237
- ^ Майер, с. 54
- ^ "Как жирафы пьют воду?" Полем Февраль 2016 года.
- ^ Манн Дж, Паттерсон Эм (ноябрь 2013 г.). «Использование инструмента водными животными» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 368 (1630): 20120424. DOI : 10.1098/rstb.2012.0424 . PMC 4027413 . PMID 24101631 .
- ^ Raffaele P (2011). Среди великих обезьян: приключения на следе наших ближайших родственников . Нью -Йорк: Харпер. п. 83. ISBN 978-0-06-167184-5 Полем OCLC 674694369 .
- ^ Келер В. (1925). Менталитет обезьян . Ливершай. ISBN 978-0-87140-108-3 Полем OCLC 2000769 .
- ^ McGowan RT, Rehn T, Norling Y, Keeling LJ (май 2014). «Позитивный аффект и обучение: изучение« эффекта Эврики »у собак». Животное познание . 17 (3): 577–587. doi : 10.1007/s10071-013-0688-x . PMID 24096703 . S2CID 15216926 .
- ^ Карбовски J (май 2007 г.). «Глобальное и региональное метаболическое масштабирование мозга и его функциональные последствия» . BMC Biology . 5 (18): 18. Arxiv : 0705.2913 . BIBCODE : 2007ARXIV0705.2913K . doi : 10.1186/1741-7007-5-18 . PMC 1884139 . PMID 17488526 .
- ^ Марино Л (июнь 2007 г.). "Мозги Кито: Насколько они водные?" Полем Анатомическая запись . 290 (6): 694–700. doi : 10.1002/ar.20530 . PMID 17516433 . S2CID 27074107 . Архивировано из оригинала 20 марта 2020 года . Получено 5 октября 2019 года .
- ^ Gallop GG (январь 1970). «Шимпанзе: самопознание». Наука . 167 (3914): 86–87. Bibcode : 1970sci ... 167 ... 86G . doi : 10.1126/science.167.3914.86 . PMID 4982211 . S2CID 145295899 .
- ^ Plotnik JM, de Waal FB, Reiss D (ноябрь 2006 г.). «Самопознание в азиатском слоне» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 103 (45): 17053–17057. Bibcode : 2006pnas..10317053p . doi : 10.1073/pnas.0608062103 . PMC 1636577 . PMID 17075063 . Архивировано (PDF) из оригинала 25 января 2024 года . Получено 25 января 2024 года .
- ^ Роберт С. (1986). «Онтогенство зеркального поведения у двух видов великих обезьян». Американский журнал приматологии . 10 (2): 109–117. doi : 10.1002/ajp.1350100202 . PMID 31979488 . S2CID 85330986 .
- ^ Walraven V, Van Elsacker L, Verheyen R (1995). «Реакции группы пигмеи шимпанзе (Pan Paniscus) на их зеркальные изображения: доказательство самопознания». Приматы . 36 : 145–150. doi : 10.1007/bf02381922 . S2CID 38985498 .
- ^ Leakey R (1994). «Происхождение ума» . Происхождение человечества . Нью -Йорк: Basicbooks. п. 150. ISBN 978-0-465-05313-1 Полем OCLC 30739453 .
- ^ Арчер Дж. (1992). Этология и человеческое развитие . Роуман и Литтлфилд. С. 215–218. ISBN 978-0-389-20996-6 Полем OCLC 25874476 .
- ^ Jump up to: а беременный Мартен К., Псаракос С. (1995). «Свидетельство самосознания в узлетенозном дельфине ( Tursiops Truncatus )». В Parker St, Mitchell R, Boccia M (Eds.). Самосознание у животных и людей: перспективы развития . Кембридж: издательство Кембриджского университета. С. 361–379. ISBN 978-0-521-44108-7 Полем OCLC 28180680 .
- ^ Jump up to: а беременный Delfour F, Marten K (апрель 2001 г.). «Обработка зеркального изображения у трех видов морских млекопитающих: киты -убийцы (Orcinus orca), ложные киты -убийцы (псевдорка Crassidens) и Калифорнийские морские львы (Zalophus californianus)». Поведенческие процессы . 53 (3): 181–190. doi : 10.1016/s0376-6357 (01) 00134-6 . PMID 11334706 . S2CID 31124804 .
- ^ Джарвис Джу (май 1981). «Euscociality у млекопитающего: кооперативное размножение в голых колониях молей». Наука . 212 (4494): 571–573. Bibcode : 1981sci ... 212..571J . doi : 10.1126/science.7209555 . JSTOR 1686202 . PMID 7209555 . S2CID 880054 .
- ^ Jacobs DS, Bennett NC, Jarvis Ju, Crowe TM (1991). «Структура колонии и иерархия доминирования думараленда кролика Cryptomys Damarensis (Rodentia: Bathyergidae) из Намибии». Журнал зоологии . 224 (4): 553–576. doi : 10.1111/j.1469-7998.1991.tb03785.x .
- ^ Харди С.Б. (2009). Матери и другие: эволюционное происхождение взаимного понимания . Бостон: Belknap Press of Harvard University Press. С. 92–93.
- ^ Harlow HF, Suomi SJ (июль 1971 г.). «Социальное выздоровление с помощью выявленных изоляционных обезьян» . Труды Национальной академии наук Соединенных Штатов Америки . 68 (7): 1534–1538. Bibcode : 1971pnas ... 68.1534H . doi : 10.1073/pnas.68.7.1534 . PMC 389234 . PMID 5283943 .
- ^ Ван Шайк CP (январь 1999 г.). «Социально -экологическая социальная социальность у орангутанов». Приматы; Журнал приматологии . 40 (1): 69–86. doi : 10.1007/bf025557703 . PMID 23179533 . S2CID 13366732 .
- ^ Archie EA, Moss CJ, Alberts SC (март 2006 г.). «Связи, которые связывают: генетическое родство предсказывает деление и слияние социальных групп у диких африканских слонов» . Разбирательство. Биологические науки . 273 (1586): 513–522. doi : 10.1098/rspb.2005.3361 . PMC 1560064 . PMID 16537121 .
- ^ Смит Дж., Меменис С.К., Хоулкамп К.Е. (2007). «Выбор партнера, связанный с рангом, в Обществе делящихся пятен пятнистой гиены ( Crocuta Crocuta )» (PDF) . Поведенческая экология и социобиология . 61 (5): 753–765. Bibcode : 2007becos..61..753s . doi : 10.1007/s00265-006-0305-y . S2CID 24927919 . Архивировано из оригинала (PDF) 25 апреля 2014 года.
- ^ Matoba T, Kutsukake N, Hasegawa T (2013). Хейворд М (ред.). «Потирание головы и облизывание усиливает социальные связи в группе африканских львов в неволе, Пантера Лео» . Plos один . 8 (9): E73044. BIBCODE : 2013PLOSO ... 873044M . doi : 10.1371/journal.pone.0073044 . PMC 3762833 . PMID 24023806 .
- ^ Krützen M, Barré LM, Connor RC, Mann J, Sherwin WB (июль 2004 г.). " О, отец: где ты? ' - Оценка отцовства в открытом делящем обществе диких узких слоев дельфинов (Tursiops sp.) В Shark Bay, Западная Австралия ». Молекулярная экология . 13 (7): 1975–1990. Bibcode : 2004molec..13.1975K . doi : 10.1111/j.1365-294x.2004.02192.x . PMID 15189218 . S2CID 4510393 .
- ^ Мартин С. (1991). Тропические леса Западной Африки: экология - угрозы - сохранение . Спрингер. doi : 10.1007/978-3-0348-7726-8 . ISBN 978-3-0348-7726-8 .
- ^ Le Roux A, Cherry Mi, Gygax L (5 мая 2009 г.). «Поведение бдительности и фитнеса: сравнение одиночного пищи и облигационного млекопитающего, занимающего группу». Поведенческая экология и социобиология . 63 (8): 1097–1107. Bibcode : 2009becos..63.1097l . doi : 10.1007/s00265-009-0762-1 . S2CID 21961356 .
- ^ Палаги Э., Норсия I (2015). Самондс Ке (ред.). «Сезон мира: примирение у деспотического вида (Lemur Catta)» . Plos один . 10 (11): E0142150. BIBCODE : 2015PLOSO..1042150P . doi : 10.1371/journal.pone.0142150 . PMC 4646466 . PMID 26569400 .
- ^ East ML, Hofer H (2000). «Очередь мужского пола Гиена ( Crocuta Crocuta ) для статуса в социальных группах, где преобладают женщины» . Поведенческая экология . 12 (15): 558–568. doi : 10.1093/beheco/12.5.558 .
- ^ Samuels A, Silk JB , Rodman P (1984). «Изменения в ранжировании доминирования и репродуктивном поведении макак мужского капота ( излучение MACACA )». Поведение животных . 32 (4): 994–1003. doi : 10.1016/s0003-3472 (84) 80212-2 . S2CID 53186523 .
- ^ Delpietro HA, Russo RG (2002). «Наблюдения за общей летучей мышью вампиров ( Desmodus rotundus ) и волосатой летучим мышью вампира ( Diphylla ecaudata ) в плену». Биология млекопитающих . 67 (2): 65–78. Bibcode : 2002 Mambi..67 ... 65d . doi : 10.1078/1616-5047-00011 .
- ^ Kleiman DG (март 1977 г.). «Моногамия у млекопитающих». Ежеквартальный обзор биологии . 52 (1): 39–69. doi : 10.1086/409721 . PMID 857268 . S2CID 25675086 .
- ^ Голландия Б., Райс В.Р. (февраль 1998 г.). «Перспектива: половой отбор погони: антагонистическое соблазнение в сравнении с резистентностью» (PDF) . Эволюция; Международный журнал органической эволюции . 52 (1): 1–7. doi : 10.2307/2410914 . JSTOR 2410914 . PMID 28568154 . Архивировано из оригинала (PDF) 8 июня 2019 года . Получено 8 июля 2016 года .
- ^ Clutton-Brock Th (май 1989). «Системы спаривания млекопитающих» . Труды Королевского общества Лондона. Серия B, биологические науки . 236 (1285): 339–372. BIBCODE : 1989RSPSB.236..339C . doi : 10.1098/rspb.1989.0027 . PMID 2567517 . S2CID 84780662 .
- ^ Boness DJ, Bowen D, Buhleier BM, Marshall GJ (2006). «Система спаривания и спариваговая система из приготовленной на водном браке: уплотнение гавани, Phoca Vitulina » . Поведенческая экология и социобиология . 61 (1): 119–130. Bibcode : 2006becos..61..119b . doi : 10.1007/s00265-006-0242-9 . S2CID 25266746 .
- ^ Клопфер PH (1981). «Происхождение родительской помощи» . В губернике DJ (ред.). Родительская помощь у млекопитающих . Нью -Йорк: Plenum Press. ISBN 978-1-4613-3150-6 Полем OCLC 913709574 .
- ^ Murthy R, Bearman G, Brown S, Bryant K, Chinn R, Hewlett A, et al. (Май 2015). «Животные в медицинских учреждениях: рекомендации по минимизации потенциальных рисков» (PDF) . Инфекционная контроль и больничная эпидемиология . 36 (5): 495–516. doi : 10.1017/ICE.2015.15 . PMID 25998315 . S2CID 541760 . Архивировано (PDF) из оригинала 3 ноября 2023 года.
- ^ Гуманное общество Соединенных Штатов. «Статистика владения домашними животными США» . Архивировано из оригинала 7 апреля 2012 года . Получено 27 апреля 2012 года .
- ^ USDA. «Профиль индустрии кроликов США» (PDF) . Архивировано из оригинала (PDF) 7 августа 2019 года . Получено 10 июля 2013 года .
- ^ Макки Р (26 мая 2013 г.). «Доисторическое пещевое искусство в Дордоге» . Хранитель . Архивировано из оригинала 31 мая 2021 года . Получено 9 ноября 2016 года .
- ^ Джонс Дж (27 июня 2014 г.). «10 лучших портретов животных в искусстве» . Хранитель . Архивировано из оригинала 18 мая 2016 года . Получено 24 июня 2016 года .
- ^ «Охота на оленей в Соединенных Штатах: анализ демографии охотника и поведения в Национальный обследование рыболовства, охоты, охоты и ассоциированного с дикой природой 2001 года» за 2001 год » . Промысел и служба дикой природы (США). Архивировано с оригинала 13 августа 2016 года . Получено 24 июня 2016 года .
- ^ Шелтон Л (5 апреля 2014 г.). «Популярность охоты на отдых, взлетевшая» . Натчез -демократ . Большой вид на открытом воздухе. Архивировано с оригинала 12 декабря 2017 года . Получено 24 июня 2016 года .
- ^ Нгуен Дж., Уитли Р. (2015). Охота за едой: Руководство по сбору урожая, полевой заправки и приготовления дикой игры . F+W Media. С. 6–77. ISBN 978-1-4403-3856-4 Полем Главы о охоте на оленей, дикой свиней (кабан), кролика и белки.
- ^ "Скачки" . Энциклопдия Британская . Архивировано из оригинала 21 декабря 2013 года . Получено 6 мая 2014 года .
- ^ Гендерс R (1981). Энциклопедия гонок борзых . Пелхэм книги. ISBN 978-0-7207-1106-6 Полем OCLC 9324926 .
- ^ Плюс S (1993). «Роль животных в человеческом обществе». Журнал социальных вопросов . 49 (1): 1–9. doi : 10.1111/j.1540-4560.1993.tb00906.x .
- ^ Фаулер К.Дж. (26 марта 2014 г.). «10 лучших книг об умных животных» . Хранитель . Архивировано из оригинала 28 мая 2021 года . Получено 9 ноября 2016 года .
- ^ Gamble N, Yates S (2008). Изучение детской литературы (2 -е изд.). Лос -Анджелес: Мудрец. ISBN 978-1-4129-3013-0 Полем OCLC 71285210 .
- ^ «Книги для взрослых» . SEAL SETTERS . Архивировано с оригинала 11 июля 2016 года . Получено 9 ноября 2016 года .
- ^ Патерсон Дж. (2013). «Животные в кино и средствах массовой информации». Оксфордские библиографии . doi : 10.1093/obo/9780199791286-0044 .
- ^ Джонс C (2011). Крупный рогатый скот: история, миф, искусство . Лондон: Британская музейная пресса. ISBN 978-0-7141-5084-0 Полем OCLC 665137673 .
- ^ Ван Гулик Р.Х. Хаягрива: Мантраяновый аспект культуры лошадей в Китае и Японии . Брилл Архив. п. 9
- ^ Grainger R (24 июня 2012 г.). «Изображение льва в древних и современных религиях» . ТРЕВОГА. Архивировано с оригинала 23 сентября 2016 года . Получено 6 ноября 2016 года .
- ^ Diamond JM (1997). «Часть 2: рост и распространение производства продуктов питания» . Оружие, микробы и сталь: судьбы человеческих обществ . Нью -Йорк: WW Norton & Company. ISBN 978-0-393-03891-0 Полем OCLC 35792200 .
- ^ Ларсон Г., Бургер Дж (апрель 2013 г.). «Генетика населения на одомашнивание животных» (PDF) . Тенденции в генетике . 29 (4): 197–205. doi : 10.1016/j.tig.2013.01.003 . PMID 23415592 . Архивировано из оригинала (PDF) 8 июня 2019 года . Получено 9 ноября 2016 года .
- ^ Зедер Ма (август 2008 г.). «Бытие и раннее сельское хозяйство в Средиземноморском бассейне: происхождение, распространение и воздействие» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (33): 11597–11604. Bibcode : 2008pnas..10511597Z . doi : 10.1073/pnas.0801317105 . PMC 2575338 . PMID 18697943 .
- ^ «Графические детали, карты и инфографика. Подсчет цыплят» . Экономист . 27 июля 2011 года. Архивировано с оригинала 15 июля 2016 года . Получено 6 ноября 2016 года .
- ^ «Породы крупного рогатого скота сегодня» . Скот сегодня . Скот-today.com. Архивировано из оригинала 15 июля 2011 года . Получено 6 ноября 2016 года .
- ^ Люкфар С.Д., Чике Пр. «Стратегии разработки проекта кроликов в системах натурального сельского хозяйства» . Продовольственная и сельскохозяйственная организация . Архивировано с оригинала 6 мая 2016 года . Получено 6 ноября 2016 года .
- ^ Pond WG (2004). Энциклопедия науки о животных . CRC Press. С. 248–250. ISBN 978-0-8247-5496-9 Полем OCLC 57033325 . Архивировано из оригинала 23 января 2023 года . Получено 5 октября 2018 года .
- ^ Braaten AW (2005). "Шерсть" . В Steele V (ред.). Энциклопедия одежды и моды . Тол. 3. Томсон Гейл . С. 441–443 . ISBN 978-0-684-31394-8 Полем OCLC 963977000 .
- ^ Quiggle C (осень 2000). «Альпака: древняя роскошь». Включите вязание : 74–76.
- ^ «Дикие млекопитающие составляют всего несколько процентов млекопитающих в мире» . Наш мир в данных . Получено 8 августа 2023 года .
- ^ «Генетические исследования» . Доверие животных. Архивировано с оригинала 12 декабря 2017 года . Получено 6 ноября 2016 года .
- ^ «Разработка наркотиков» . Animal Research.info. Архивировано с оригинала 8 июня 2016 года . Получено 6 ноября 2016 года .
- ^ «Статистика ЕС показывает снижение числа исследований животных» . Говоря об исследованиях. 2013. Архивировано с оригинала 24 апреля 2019 года . Получено 6 ноября 2016 года .
- ^ Pilcher HR (2003). «Это нокаут» . Природа . doi : 10.1038/news030512-17 . Архивировано с оригинала 10 ноября 2016 года . Получено 6 ноября 2016 года .
- ^ «Поставка и использование приматов в ЕС» . Европейская ассоциация биомедицинских исследований. 1996. Архивировано из оригинала 17 января 2012 года.
- ^ Карлссон он, Шапиро С.Дж., Фара I, Хау Дж (август 2004 г.). «Использование приматов в исследованиях: глобальный обзор». Американский журнал приматологии . 63 (4): 225–237. doi : 10.1002/ajp.20054 . PMID 15300710 . S2CID 41368228 .
- ^ Weatherall D, et al. (2006). Использование нечеловеческих приматов в исследованиях (PDF) (отчет). Лондон: Академия медицинских наук. Архивировано из оригинала (PDF) 23 марта 2013 года.
- ^ Ritchie H , Roser M (15 апреля 2021 года). «Биоразнообразие» . Наш мир в данных . Архивировано из оригинала 11 декабря 2022 года . Получено 29 августа 2021 года .
- ^ Jump up to: а беременный Bar-On YM, Phillips R, Milo R (июнь 2018 г.). «Распределение биомассы на Земле» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (25): 6506–6511. BIBCODE : 2018PNAS..115.6506B . doi : 10.1073/pnas.1711842115 . PMC 6016768 . PMID 29784790 .
- ^ Цена E (2008). Принципы и применение поведения домашнего животного: вводной текст . Сакраменто: издательство Кембриджского университета. ISBN 978-1-84593-398-2 Полем OCLC 226038028 .
- ^ Taupitz J, Weschka M (2009). Шимбриды - химеры и гибриды в сравнительных европейских и международных исследованиях . Гейдельберг: Спрингер. п. 13. ISBN 978-3-540-93869-9 Полем OCLC 495479133 .
- ^ Chambers SM, Fain SR, Fazio B, Amaral M (2012). «Отчет о таксономии североамериканских волков из морфологического и генетического анализа» . Североамериканская фауна . 77 : 2. DOI : 10.3996/NAFA.77.0001 . Архивировано из оригинала 31 мая 2021 года . Получено 12 октября 2019 года .
- ^ Van Vuure T (2005). Позвольте аурохам - историю, морфологию и экологию вымершего дикого быка . Pensoft Publishers. ISBN 978-954-642-235-4 Полем OCLC 940879282 .
- ^ Mooney Ha, Cleland EE (май 2001 г.). «Эволюционное влияние инвазивных видов» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (10): 5446–5451. Bibcode : 2001pnas ... 98.5446m . doi : 10.1073/pnas.091093398 . PMC 33232 . PMID 11344292 .
- ^ Le Roux JJ, Foxcroft LC, Herbst M, Macfadyen S (январь 2015). «Генетический анализ показывает низкий уровень гибридизации между африканскими дикими кошками (Felis Silvestris Lybica) и домашними кошками (F. s. Catus) в Южной Африке» . Экология и эволюция . 5 (2): 288–299. Bibcode : 2015ECOEV ... 5..288L . doi : 10.1002/ECE3.1275 . PMC 4314262 . PMID 25691958 .
- ^ Уилсон А. (2003). Австралийский отчет о состоянии лесов . п. 107
- ^ Rhymer JM, Simberloff D (ноябрь 1996 г.). «Вымирание гибридизацией и интрогрессией». Ежегодный обзор экологии и систематики . 27 : 83–109. doi : 10.1146/annurev.ecolsys.27.1.83 .
- ^ Поттс Б.М. (2001). Barbour RC, Hingston AB (Eds.). Генетическое загрязнение от сельскохозяйственного лесного хозяйства с использованием видов и гибридов эвкалипта: отчет для совместного предприятия RIRDC/L & WA/FWPRDC . Сельская промышленная корпорация и корпорация разработок Австралии. ISBN 978-0-642-58336-9 Полем OCLC 48794104 .
- ^ Jump up to: а беременный Dirzo R, Young HS, Galetti M, Ceballos G, Isaac NJ, Collen B (июль 2014 г.). «Дефунация в антропоцене» (PDF) . Наука . 345 (6195): 401–406. Bibcode : 2014sci ... 345..401d . doi : 10.1126/science.1251817 . PMID 25061202 . S2CID 206555761 . Архивировано (PDF) из оригинала 7 августа 2019 года . Получено 25 января 2024 года .
- ^ Primack R (2014). Основы биологии сохранения (6 -е изд.). Sunderland, MA: Sinauer Associates, Inc. Издатели. С. 217–245. ISBN 978-1-60535-289-3 Полем OCLC 876140621 .
- ^ Vignieri S (июль 2014 г.). «Исчезающая фауна. Введение» . Наука . 345 (6195): 392–395. Bibcode : 2014sci ... 345..392V . doi : 10.1126/science.345.6195.392 . PMID 25061199 .
- ^ Burney Da, Flannery TF (июль 2005 г.). «Пятьдесят тысячелетий катастрофических вымираний после контакта с человеком» (PDF) . Тенденции в экологии и эволюции . 20 (7): 395–401. doi : 10.1016/j.tree.2005.04.022 . PMID 16701402 . Архивировано из оригинала (PDF) 10 июня 2010 года.
- ^ Diamond J (1984). «Исторические исчезновения: розеттский камень для понимания доисторических вымираний». В Martin PS, Klein RG (Eds.). Четвертичные вымирания: доисторическая революция . Тусон: Университет Аризоны Пресс. С. 824–862. ISBN 978-0-8165-1100-6 Полем OCLC 10301944 .
- ^ Уоттс J (6 мая 2019 г.). «Человеческое общество под срочной угрозой от потери естественной жизни Земли» . Хранитель . Архивировано из оригинала 14 июня 2019 года . Получено 1 июля 2019 года .
- ^ McGrath M (6 мая 2019). «Кризис природы: люди» угрожают 1M видам с исчезновением » . Би -би -си . Архивировано с оригинала 30 июня 2019 года . Получено 1 июля 2019 года .
- ^ Главный D (22 ноября 2013 г.). «7 знаковых животных, которые люди доходят до исчезновения» . Живая наука . Архивировано из оригинала 6 января 2023 года . Получено 25 января 2024 года .
- ^ Платт -младший (25 октября 2011 г.). «Браконьеры приводят Javan Rhino к вымиранию во Вьетнаме» . Scientific American . Архивировано с оригинала 6 апреля 2015 года.
- ^ Каррингтон Д. (8 декабря 2016 г.). «Жирафы сталкиваются с вымиранием после разрушительного снижения, предупреждают эксперты» . Хранитель . Архивировано из оригинала 13 августа 2021 года . Получено 4 февраля 2017 года .
- ^ Estrada A, Garber PA, Rylands AB, Roos C, Fernandez-Duque E, Di Fiore A, et al. (Январь 2017). «Предстоящий кризис вымирания мировых приматов: почему приматы имеют значение» . Наука достижения . 3 (1): E1600946. Bibcode : 2017scia .... 3E0946E . doi : 10.1126/sciadv.1600946 . PMC 5242557 . PMID 28116351 .
- ^ Флетчер М (31 января 2015 г.). «Панголины: почему это милое доисторическое млекопитающее сталкивается с вымиранием» . Телеграф . Архивировано из оригинала 10 января 2022 года.
- ^ Гринфилд П (9 сентября 2020 года). «Люди эксплуатируют и разрушают природу в беспрецедентных масштабах - сообщают» . Хранитель . Архивировано из оригинала 21 октября 2021 года . Получено 13 октября 2020 года .
- ^ Маккарти Д. (1 октября 2020 г.). «Ужасные потери дикой природы показывают, что началась игра вымирания - но еще не поздно для перемен» . Независимый . Архивировано из оригинала 7 апреля 2023 года . Получено 13 октября 2020 года .
- ^ Pennisi E (18 октября 2016 г.). «Люди охотятся на приматов, летучих мышей и других млекопитающих для вымирания» . Наука . Архивировано из оригинала 20 октября 2021 года . Получено 3 февраля 2017 года .
- ^ Ripple WJ, Abernethy K, Betts MG, Chapron G, Dirzo R, Galetti M, et al. (Октябрь 2016). «Охота в буштинге и риск вымирания для млекопитающих в мире» . Королевское общество открыто наука . 3 (10): 160498. BIBCODE : 2016RSOS .... 360498R . doi : 10.1098/rsos.160498 . HDL : 1893/24446 . PMC 5098989 . PMID 27853564 .
- ^ Williams M, Zalasiewicz J, Haff PK, Schwägerl C, Barnosky AD , Ellis EC (2015). «Антропоцена биосфера». Антропоценовый обзор . 2 (3): 196–219. Bibcode : 2015antrv ... 2..196w . doi : 10.1177/2053019615591020 . S2CID 7771527 .
- ^ Морелл V (11 августа 2015 г.). «Исследование предупреждает« Мясные поляры, которые могут ускорить вымирание видов по всему миру » . Наука . Архивировано с оригинала 20 декабря 2016 года . Получено 3 февраля 2017 года .
- ^ Machovina B, Feeley KJ, Ripple WJ (декабрь 2015 г.). «Сохранение биоразнообразия: ключом является сокращение потребления мяса». Наука общей среды . 536 : 419–431. Bibcode : 2015scten.536..419m . doi : 10.1016/j.scitotenv.2015.07.022 . PMID 26231772 .
- ^ Редфорд К.Х. (1992). «Пустой лес» (PDF) . Биоссака . 42 (6): 412–422. doi : 10.2307/1311860 . JSTOR 1311860 . Архивировано (PDF) из оригинала 28 февраля 2021 года . Получено 4 февраля 2017 года .
- ^ Peres CA, Nascimento HS (2006). «Влияние охоты на игры на каяпо на юго-восточной Амазонии: последствия для сохранения дикой природы в запасах коренных народов тропических лесов». Человеческая эксплуатация и сохранение биоразнообразия . Тол. 3. Springer. С. 287–313. ISBN 978-1-4020-5283-5 Полем OCLC 207259298 .
- ^ Altrichter M, Boaglio G (2004). «Распределение и относительная численность пекари в аргентинском чако: ассоциации с человеческими факторами». Биологическое сохранение . 116 (2): 217–225. Bibcode : 2004bcons.116..217a . doi : 10.1016/s0006-3207 (03) 00192-7 .
- ^ Гобуш К. «Влияние браконьерства на африканских слонов» . Центр биологии сохранения . Университет Вашингтона. Архивировано из оригинала 8 декабря 2021 года . Получено 12 мая 2021 года .
- ^ Alverson DL, Freeburg MH, Murawski SA, Pope JG (1996) [1994]. «Прилова морских млекопитающих» . Глобальная оценка рыболовства и выбросов . Рим: Продовольственная и сельскохозяйственная организация Организации Объединенных Наций. ISBN 978-92-5-103555-9 Полем OCLC 31424005 . Архивировано из оригинала 17 февраля 2019 года . Получено 25 января 2024 года .
- ^ Glowka L, Burhenne-Guilmin F, Synge HM, McNeely JA, Gündling L (1994). Экологическая политика МСОП и юридический документ . Руководство по соглашению о биоразнообразии. Международный союз сохранения природы. ISBN 978-2-8317-0222-3 Полем OCLC 32201845 .
- ^ "О МСКН" . Международный союз сохранения природы. 3 декабря 2014 года. Архивировано с оригинала 15 апреля 2020 года . Получено 3 февраля 2017 года .
- ^ Ceballos G, Ehrlich PR, Barnosky AD, García A, Pringle RM, Palmer TM (июнь 2015 г.). «Ускоренные современные потери видов, вызванные человеком: войти в шестое массовое вымирание» . Наука достижения . 1 (5): E1400253. BIBCODE : 2015SCIA .... 1E0253C . doi : 10.1126/sciadv.1400253 . PMC 4640606 . PMID 26601195 .
- ^ Фишер до, Бломберг С.П. (апрель 2011 г.). «Корреляты повторного открытия и обнаружения вымирания у млекопитающих» . Разбирательство. Биологические науки . 278 (1708): 1090–1097. doi : 10.1098/rspb.2010.1579 . PMC 3049027 . PMID 20880890 .
- ^ Ceballos G, Ehrlich AH, Ehrlich PR (2015). Аннигиляция природы: человеческое исчезновение птиц и млекопитающих . Балтимор: издательство Джона Хопкинса. п. 69. ISBN 978-1-4214-1718-9 .
- ^ Jiang, Z.; Харрис, RB (2016). « Элафур Давидиан » . МСОП красный список угрожаемых видов . 2016 : E.T7121A22159785. doi : 10.2305/iucn.uk.2016-2.rlts.t7121a22159785.en . Получено 12 ноября 2021 года .
- ^ Jump up to: а беременный McKinney ML, Schoch R, Yonavjak L (2013). «Сохранение биологических ресурсов» . Наука окружающей среды: системы и решения (5 -е изд.). Jones & Bartlett Learning. ISBN 978-1-4496-6139-7 Полем OCLC 777948078 .
- ^ Perrin WF, Würsig BF, TheWissen JG (2009). Энциклопедия морских млекопитающих . Академическая пресса. п. 404. ISBN 978-0-12-373553-9 Полем OCLC 455328678 .
- Начиная с этого редактирования , в этой статье используется контент из «анатомии и физиологии» , который лицензируется таким образом, что позволяет повторно использовать в соответствии с лицензией Creative Commons Attribution-Sharealike 3.0 , но не под GFDL . Все соответствующие условия должны соблюдаться.
Дальнейшее чтение
- Браун WM (2001). «Естественный отбор компонентов мозга млекопитающих». Тенденции в экологии и эволюции . 16 (9): 471–473. Bibcode : 2001tecoe..16..471b . doi : 10.1016/s0169-5347 (01) 02246-7 .
- McKenna MC, Bell SK (1997). Классификация млекопитающих выше уровня видов . Нью -Йорк: издательство Колумбийского университета. ISBN 978-0-231-11013-6 Полем OCLC 37345734 . [ Постоянная мертвая ссылка ]
- Новак Р.М. (1999). Уокерские млекопитающие мира (6 -е изд.). Балтимор: издательство Джона Хопкинса. ISBN 978-0-8018-5789-8 Полем OCLC 937619124 .
- Симпсон Г.Г. (1945). «Принципы классификации и классификация млекопитающих». Бюллетень Американского музея естественной истории . 85 : 1–350.
- Murphy WJ, Eizirik E, O'Brien SJ, Madsen O, Scally M, Doudy CJ, et al. (Декабрь 2001 г.). «Разрешение раннего плацентарного излучения млекопитающих с использованием байесовской филогенетики». Наука . 294 (5550): 2348–2351. Bibcode : 2001sci ... 294.2348m . doi : 10.1126/science.1067179 . PMID 11743200 . S2CID 34367609 .
- Springer MS, Stanhope MJ, Madsen O, De Jong WW (август 2004 г.). «Молекулы консолидируют плацентарное дерево млекопитающих» (PDF) . Тенденции в экологии и эволюции . 19 (8): 430–438. doi : 10.1016/j.tree.2004.05.006 . PMID 16701301 . S2CID 1508898 . Архивировано из оригинала (PDF) 29 июля 2016 года . Получено 21 января 2005 года .
- Воган Т.А., Райан Дж. М., Капзапльюски Нью -Джерси (2000). МАММАЛОГИ (4 -е изд.). Форт -Уэрт, Техас: Saunders College Publishing. ISBN 978-0-03-025034-7 Полем OCLC 42285340 .
- Кригс Джо, Чураков Г., Кифманн М., Джордан У, Брозиус Дж., Шмитц Дж. (Апрель 2006 г.). «Ретрополированные элементы как архивы эволюционной истории плацентарных млекопитающих» . PLOS Биология . 4 (4): E91. doi : 10.1371/journal.pbio.0040091 . PMC 1395351 . PMID 16515367 .
- MacDonald DW, Norris S (2006). Энциклопедия млекопитающих (3 -е изд.). Лондон: коричневая справочная группа. ISBN 978-0-681-45659-4 Полем OCLC 74900519 .
Внешние ссылки
- ASM Архивирована . база данных о разнообразии млекопитающих
- BioDistryMapping.org-Все заказы млекопитающих в мире с картами распределения архивировали 2016-09-26 на The Wayback Machine
- Палеоценовые млекопитающие архивировали 3 февраля 2024 года в The Wayback Machine , участке, покрывающем рост млекопитающих, палеоценовых млекопитающих.
- Эволюция млекопитающих архивировала 25 января 2024 года в The Wayback Machine , краткое введение в ранних млекопитающих, Enchantedlearning.com
- Европейский атлас млекопитающих Эмма Архивировал 25 января 2024 года на машине Wayback от Societas Europaea Mammalogica, Europeanmals.org.org
- Морские млекопитающие мирового архивирования 8 июня 2019 года в The Wayback Machine -обзор всех морских млекопитающих, включая описания, как полностью водные, так и полуаватные, noaa.gov
- Mammalogy.org Arackived 1 марта 2020 года. В The Wayback Machine Американское общество маммалогов было создано в 1919 году с целью содействия изучению млекопитающих, и этот веб -сайт включает библиотеку изображений млекопитающих










