перекос GC

Перекос GC – это когда нуклеотидов гуанина и цитозина в избытке или недостатке в определенной области ДНК или РНК . Перекос GC также является статистическим методом измерения чрезмерной представленности гуанина для конкретной цепи. [1]
В условиях равновесия (без мутационного или селективного давления и при случайном распределении нуклеотидов внутри генома присутствует одинаковая частота четырех оснований ДНК ( аденина , гуанина , тимина и цитозина ). ) на обеих одиночных нитях молекулы ДНК [2] Однако у большинства бактерий (например, E. coli ) и некоторых архей (например, Sulfolobus solfataricus асимметричен ) нуклеотидный состав между ведущей и отстающей цепью : ведущая цепь содержит больше гуанина (G) и тимина (Т), тогда как отстающая цепь содержит больше аденина (А) и цитозина (С). [2] Это явление называется перекосом GC и AT , и была определена соответствующая статистика. [2] как:
Перекос GC = (G – C)/(G + C)
Перекос AT = (A − T)/(A + T)
Асимметричный нуклеотидный состав
[ редактировать ]Работа Эрвина Чаргаффа в 1950 году продемонстрировала, что в ДНК основания гуанин и цитозин обнаружены в равном количестве, а основания аденин и тимин обнаружены в равном количестве. Однако не было равенства между количеством одной пары и другой. [3] Открытие Чаргаффа называется правилом Чаргаффа или правилом четности 2 . [3] Три года спустя Уотсон и Крик использовали этот факт при выводе структуры ДНК, своей модели двойной спирали .
Естественным результатом правила четности 1 в состоянии равновесия, в котором нет мутаций и/или ошибок отбора ни в одной из двух цепей ДНК, является то, что при одинаковой скорости замещения комплементарные нуклеотиды в каждой цепи имеют одинаковую структуру. равные количества данного основания и его дополнения. [4] Другими словами, в каждой цепи ДНК частота встречаемости Т равна А, а частота встречаемости G равна С, поскольку скорость замещения предположительно одинакова. Это явление называется правилом четности 2 . Следовательно, второе правило четности существует только тогда, когда нет мутации или замены.
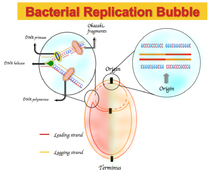
Любое отклонение от правила четности 2 приведет к асимметричному составу оснований, который отличает ведущую цепь – то есть цепь ДНК, которая реплицируется в прямом направлении – от отстающей цепи. Эта асимметрия называется перекосом GC или AT. [2]
В геномах некоторых бактерий наблюдается обогащение гуанина над цитозином и тимина над аденином на ведущей цепи и наоборот для отстающей цепи. Спектры асимметрии нуклеотидного состава варьируются от -1, что соответствует G = 0 или A = 0, до +1, что соответствует T = 0 или C = 0. [2] Следовательно, положительный перекос GC представляет богатство G по сравнению с C, а отрицательный перекос GC представляет богатство C по сравнению с G. В результате можно ожидать увидеть положительный перекос GC и отрицательный перекос AT в ведущей цепи, а отрицательный перекос GC и положительный перекос АТ в отстающей цепи. [5] Перекос GC или AT меняет знак на границах двух репликоров , которые соответствуют началу или концу репликации ДНК. [2] [4] [5] Первоначально этот асимметричный состав нуклеотидов объяснялся различным механизмом репликации ДНК между ведущей и отстающей цепью. Репликация ДНК сама по себе является полуконсервативным и асимметричным процессом. [6] Эта асимметрия обусловлена формированием репликационной вилки и ее разделением на возникающие ведущие и отстающие цепи. Ведущая цепь синтезируется непрерывно и параллельно с ведущей цепью; отстающая цепь реплицируется через короткие фрагменты полинуклеотида ( фрагменты Оказаки ) в направлении от 5' к 3'. [6]
Графики перекоса вычислений и GC
[ редактировать ]Существует три основных подхода к расчету и графической демонстрации перекоса GC и его свойств.
Асимметрия GC
[ редактировать ]Первый подход — асимметрия GC и AT. [2] Жан Р. Лобри первым сообщил в 1996 году: [7] наличие композиционной асимметрии в геномах трех бактерий: E. coli , Bacillus subtilis и Haemophilus influenzae . Исходные формулы в то время назывались не перекосом, а скорее отклонением от [A] = [T] или [C] = [G]:
отклонение от [A] = [T] как (A − T)/(A + T);
отклонение от [C] = [G] как (C − G)/(C + G);
где A, T, G и C представляют частоту появления эквивалентного основания в определенной последовательности определенной длины. Стратегия скользящего окна используется для расчета отклонения от C по всему геному. На этих графиках положительное отклонение от C соответствует отстающей цепи, а отрицательное отклонение от C соответствует ведущей цепи. [8] Кроме того, место переключения знака отклонения соответствует началу координат или терминалу. Ось X представляет расположение хромосом, нанесенное на графике от 5 'до 3', а ось Y представляет значение отклонения. Основным недостатком этого метода является его свойство зависимости от размера окна. Поэтому выбор подходящего размера окна сильно влияет на результат сюжета. Другие методы следует сочетать с отклонениями, чтобы с большей точностью идентифицировать и определить местонахождение источника репликации ДНК.
перекос CGC
[ редактировать ]
Второй подход называется кумулятивным перекосом GC (перекос CGC). [9] Этот метод по-прежнему использует стратегию скользящего окна, но использует сумму соседних окон с произвольного начала. В этой схеме весь геном обычно изображается от 5 до 3 футов с использованием произвольного начала и произвольной цепи. На совокупном графике перекоса GC пики соответствуют точкам переключения (конец или начало координат).
В отличие от более ранней статьи Лобри, недавние реализации перекоса GC переворачивают исходное определение, переопределяя его следующим образом:
Перекос GC = (G - C)/(G + C).
При перевернутом определении перекоса GC максимальное значение совокупного перекоса соответствует терминалу, а минимальное значение соответствует источнику репликации.
Z-кривая
[ редактировать ]Последний подход — Z. кривая [10] В отличие от предыдущих методов, этот метод не использует стратегию скользящего окна и считается более эффективным при поиске источника репликации. [10] каждого основания В этом методе исследуется совокупная частота по отношению к основанию в начале последовательности. Кривая Z использует трехмерное представление со следующими параметрами:



Где , представляет собой избыток пурина над пиримидином, обозначает избыток кето над амино, и показывает взаимосвязь между слабыми и сильными водородными связями . и компоненты могут сами по себе обнаружить начало репликации и асимметричный состав цепей. Комбинацию этих методов следует использовать для предсказания начала и окончания репликации, чтобы компенсировать их слабость.






Механизм
[ редактировать ]В научном сообществе нет единого мнения относительно механизма, лежащего в основе смещения нуклеотидного состава внутри каждой цепи ДНК. Существуют две основные школы мысли, объясняющие механизм специфического нуклеотидного состава цепи у бактерий. [4]
Первый описывает смещение и асимметричное мутационное давление на каждую цепь ДНК во время репликации и транскрипции . [4] [11] Из-за асимметричного характера процесса репликации неравная частота мутаций и эффективность восстановления ДНК во время процесса репликации могут привести к большему количеству мутаций в одной цепи по сравнению с другой. [5] Более того, время, необходимое для репликации между двумя цепями, варьируется и может привести к асимметричному мутационному давлению между ведущей и отстающей цепями. [12] Помимо мутаций во время репликации ДНК, транскрипционные мутации могут привести к искажению нуклеотидного состава конкретной цепи. [5] Дезаминирование цитозина и, в конечном итоге, мутация цитозина на тимин в одной цепи ДНК может увеличить относительное количество гуанина и тимина по отношению к цитозину и аденину. [5] У большинства бактерий большая часть генов закодирована в ведущей цепи. [4] Например, ведущая цепь Bacillus subtilis кодирует 75% генов. [5] Кроме того, сообщалось об избытке дезаминирования и превращения цитозина в тимин в кодирующей цепи по сравнению с некодирующей цепью. [4] [5] [13] Одним из возможных объяснений является то, что нетранскрибируемая цепь ( кодирующая цепь ) во время процесса транскрипции является одноцепочечной; следовательно, он более уязвим к дезаминированию по сравнению с транскрибируемой цепью ( некодирующей цепью ). [5] [14] Другое объяснение состоит в том, что активность репарации дезаминирования во время транскрипции не происходит на кодирующей цепи. [5] Только транскрибируемая цепь получает пользу от этих событий репарации дезаминирования.
Вторая школа мысли описывает механизм перекоса GC и AT как результат разницы в селективном давлении между ведущей и отстающей цепями. [4] [5] [14] Изучение генома прокариот показывает предпочтение в положении третьего кодона G над C и T над A. [5] Эта дискриминация создает асимметричный нуклеотидный состав, если кодирующая цепь неравномерно распределена между ведущей и отстающей цепями, как в случае бактерий. Кроме того, было показано, что высоко транскрибируемые гены, такие как рибосомальные белки , у бактерий расположены в основном на ведущей цепи. [5] Следовательно, смещение в выборе кодона третьего положения G по сравнению с C может привести к перекосу GC. Кроме того, некоторые сигнальные последовательности богаты гуанином и тимином, например последовательности chi , и эти последовательности могут иметь более высокую частоту встречаемости в одной цепи по сравнению с другой. [4] [5]
И мутационное, и селективное давление могут независимо друг от друга вызывать асимметрию в цепях ДНК. Однако сочетание и кумулятивный эффект обоих механизмов является наиболее правдоподобным объяснением неравномерности GC и AT. [4] [14]
Использование
[ редактировать ]Доказано, что перекос GC полезен в качестве индикатора ведущей цепи ДНК, отстающей цепи, начала репликации и терминала репликации. [2] [4] [5] Большинство бактерий и архей содержат только одну точку начала репликации ДНК. [2] Перекос GC положителен и отрицателен в ведущей цепи и в отстающей цепи соответственно; поэтому ожидается, что будет наблюдаться изменение знака перекоса GC только в точке начала и конца репликации ДНК. [4] Перекос GC также можно использовать для изучения смещений цепей и связанных с ними механизмов путем расчета избытка одного основания над дополнительным основанием в разных средах. [4] [5] [14] Такие методы, как перекос GC, перекос CGC и кривая Z, представляют собой инструменты, которые могут дать возможность лучше исследовать механизм репликации ДНК в различных организмах.
Ссылки
[ редактировать ]- ^ Кеннеди, Шон П.; Нг, Вайлап Виктор; Зальцберг, Стивен Л.; Худ, Лерой; ДасСарма, Шиладитья (1 октября 2001 г.). «Понимание адаптации видов галобактерий NRC-1 к экстремальным условиям окружающей среды посредством компьютерного анализа последовательности их генома» . Геномные исследования . 11 (10): 1641–1650. дои : 10.1101/гр.190201 . ISSN 1088-9051 . ПМК 311145 . ПМИД 11591641 .
- ^ Jump up to: а б с д и ж г час я Лобри, Дж. Р. Паттерны асимметричных замен в двух цепях ДНК бактерий. Молекулярная биология и эволюция 13, 660-665 (1996).
- ^ Jump up to: а б Чаргафф, Э. Химическая специфичность нуклеиновых кислот и механизм их ферментативной деградации. Опыт 6, 201–209 (1950).
- ^ Jump up to: а б с д и ж г час я дж к л Нексулеа А. и Лобри Дж. Р. Новый метод оценки влияния репликации на асимметрию состава оснований ДНК. Молекулярная биология и эволюция 24, 2169-2179, doi:10.1093/molbev/msm148 (2007).
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Тилье, Э.Р. и Коллинз, Р.А. Вклад ориентации репликации, направления генов и сигнальных последовательностей в асимметрию базового состава в бактериальных геномах. Журнал молекулярной эволюции 50, 249–257 (2000).
- ^ Jump up to: а б Роча, Е.П. Репликационная организация бактериальных геномов. Microbiology 150, 1609-1627, doi:10.1099/mic.0.26974-0 (2004).
- ^ Лобри, младший (май 1996 г.). «Асимметричные закономерности замещения в двух цепях ДНК бактерий» . Молекулярная биология и эволюция . 13 (5): 660–665. doi : 10.1093/oxfordjournals.molbev.a025626 . ISSN 0737-4038 . ПМИД 8676740 .
- ^ "Пояснение статьи Лобри 1996 года"
- ^ Григорьев, А. Анализ геномов с помощью кумулятивных диаграмм перекоса. Nucleic Acids Research 26, 2286-2290 (1998).
- ^ Jump up to: а б Чжан Р. и Чжан К.Т. Истоки множественной репликации археи Halobacterium вида NRC-1. Сообщения о биохимических и биофизических исследованиях 302, 728-734 (2003).
- ^ Лобри, Дж. Р. и Суеока, Н. Асимметричное направленное мутационное давление у бактерий. Геномная биология 3, RESEARCH0058 (2002).
- ^ Эппингер М., Баар К., Раддац Г., Хьюсон Д.Х. и Шустер С.С. Сравнительный анализ четырех Campylobacterales. Обзоры природы. Microbiology 2, 872-885, doi:10.1038/nrmicro1024 (2004).
- ^ Марин, А. и Ся, X. Перекос GC в генах, кодирующих белки, между ведущими и отстающими цепями в бактериальных геномах: новые модели замещения, включающие смещение цепей. Журнал теоретической биологии 253, 508-513, doi:10.1016/j.jtbi.2008.04.004 (2008).
- ^ Jump up to: а б с д Чарнески К.А., Хонти Ф., Брайант Дж.М., Херст Л.Д., Фейл Э.Дж. Атипичный перекос AT в геномах фирмикутов является результатом отбора, а не мутации. PLoS Genetics 7(9):e1002283 (2011).
- Мьюз, HW и др. MIPS: анализ и аннотация белков целых геномов в 2005 г. Нуклеиновые кислоты Res 34, D169-172, doi:10.1093/nar/gkj148 (2006).