Формация Джебель Катрани
| Формация Джебель Катрани | |
|---|---|
| Стратиграфический диапазон : Эоцен - Олигоцен. ~ | |
 | |
| Тип | Формирование |
| Подразделения | Нижняя последовательность (нижняя зона ископаемой древесины)
Баритовый песчаник Верхняя толща (верхняя зона ископаемых пород) |
| Подложки | Видан эль Фарас Базальт |
| Накладки | Каср из формации Сагха |
| Область | Фаюмская депрессия |
| Толщина | 340 м |
| Литология | |
| Начальный | песчаник , аргиллит |
| Расположение | |
| Область | Мухафаза Файюм |
| Страна | |
Формация Джебель-Катрани (также Гебель-Катрани , Габал-Катрани или Джебель-Катрани ) — геологическое образование , расположенное в провинции Файюм в центральном Египте . Он расположен между откосом Джебель-Катрани и откосом Каср-эль-Сагха , к северу от Биркет-Карун, озера недалеко от Файюма . Формация согласно перекрывает формацию Каср-эль-Сагха и увенчана базальтом Видан-эль-Фарас . Возраст формации является предметом споров, но самые последние исследования показывают, что она охватывает как поздние части эоцена , так и ранний олигоцен , охватывая границу между этими двумя периодами времени.
Геология и фауна этого образования дают хорошее представление об окружающей среде и животных, обитавших в этот период времени. Исследования показывают, что формация Джебель-Катрани представляла собой смесь субтропических и тропических лесов , низменных болот и топей , прудов и рек, которые впадали на север в море Тетис . Это подтверждается наличием водозависимой фауны, включая подокнемидидовых черепах , крокодилов , морских коров , различных рыб, якан , ранних родственников фламинго , скоп , цапель и китоглавов .
Помимо этого, летопись окаменелостей формации Джебель-Катрани особенно известна своей ценностью для понимания ранней эволюции многих современных групп млекопитающих. Приматы представлены более чем десятком родов несколько форм ранних слонов , из отложений обнаружено , включая наземную Phiomia и полуводную Moeritherium . Между тем, ископаемые грызуны формации считаются важным связующим звеном между африканскими фиоморфами ( ромашки , дикобразы Старого Света , землекопы и тростниковые крысы ) и кавиоморфами Южной Америки ( капибары , шиншиллы и дикобразы Нового Света ). Помимо этих первых членов групп, которые позже приобрели известность, формация также была домом для множества уникальных групп, которые больше не встречаются сегодня или встречаются в значительно уменьшенном разнообразии. Сюда входят загадочные, возможно, плотоядные птолемаиды , крупные гиенодонты , огромное количество весьма разнообразных гиракоид, включая виды размером с носорога, антракотериев и причудливых Эмбритопод Арсинотериум .
География и история
[ редактировать ]
Обнажения формации Джебель-Катрани присутствуют в северной части Фаюмской впадины к юго-западу от Каира . [ 1 ] Фаюмская впадина — оазис к западу от Нила на севере Египта . [ 2 ]
Образования Фаюма в течение значительного времени изучались многочисленными палеонтологами, причем исследования начались еще в XIX веке. За это время регион широко изучался учеными, включая, помимо прочего, Чарльза Уильяма Эндрюса , Генри Фэрфилда Осборна , Рене Фуртау и Эрнста Стромера . Среди первых названий того, что сейчас известно как формация Джебель-Катрани, была «Ряд Флувио-Марин», придуманная Хью Дж. Л. Биднеллом. Однако, несмотря на большую часть исследований, проведенных в конце 19-го и начале 20-го века, населенные пункты Фаюма в конечном итоге вошли в период безвестности после начала Первой мировой войны , которая продолжалась на протяжении всей середины 20-го века и Второй мировой войны . Исследования возобновились в 1960-х годах после экспедиции под руководством Элвина Лаверна Саймонса . [ 2 ] Раньше формации региона были известны прежде всего сохранением млекопитающих, но птицы также были известны по нескольким экземплярам, обнаруженным в начале 20 века. В 60-е годы улучшенные методы сбора и дополнительные экспедиции Йельского университета и Университета Дьюка собрали много дополнительного материала совместно с Египетской геологической службой и General Petroleum Company . [ 1 ] Примерно в 70-х годах масштабы экспедиции были расширены и теперь охватывают более разнообразные области исследований, что привело к более точному датированию слоев. [ 2 ]
Геология и стратиграфия
[ редактировать ]
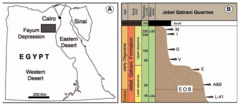
Эта формация перекрывает эоценовую формацию Каср-эль-Сагха и перекрывается олигоценовым базальтом Видан-эль-Фарас . Формация содержит по крайней мере два основных слоя, содержащих ископаемые: один в верхней последовательности, которая используется для обозначения верхних слоев формации, и второй в нижней последовательности. Эти два слоя в старых публикациях также назывались зонами верхней и нижней древесины. Обе толщи формации Джебель-Катрани отделены друг от друга так называемым баритовым песчаником, слоем толщиной 4–10 м (13–33 футов). [ 1 ] [ 2 ]
Датировка формации Джебель-Катрани исторически является предметом споров: некоторые исследования относят ее либо полностью к эоцену, либо к олигоцену, а некоторые утверждают, что она охватывает оба периода. Расмуссен и его коллеги, например, утверждали, что центральные баритовые песчаники, разделяющие оба слоя, содержащие ископаемые, отмечают точную границу эоцена и олигоцена . Частично это связано с фауной приматов формации: проплиопитециды и парапитецины появляются только в верхних местах. Между тем другие методы датирования в целом оказались неприменимы. По мнению Зейферта, фауна млекопитающих формации сама по себе слишком эндемична, а морские беспозвоночные отсутствуют, что препятствует биостратиграфическому датированию. Радиоизотопное датирование было проведено на вышележащем базальте Видан-эль-Фарас, однако результаты этого исследования позволили предположить, что возраст его нижних слоев составляет 23,6 миллиона лет, что намного моложе, чем предыдущие оценки формации Джебель-Катрани. Однако в публикации 2006 года Зейферт проводит сравнение между фаюмской фауной и летописью окаменелостей Формация Ашаук в Омане , в которой сохранились позвоночные, а также фораминиферы, полезные для датирования. Биостратиграфия и магнитостратиграфия фораминифер позволяют предположить, что исследованные местонахождения в Омане датируются примерно 31-31,5 млн лет назад (местонахождение Таках) и 33,7-33,3 млн лет назад (местонахождение Тайтинити). Зайферт утверждает, что фауна млекопитающих Омана наиболее близка к фауне млекопитающих из древнейших местонахождений нижней толщи формации Джебель-Катрани (L-41) и древнейших местонахождений верхней толщи (карьеры G и V). Даманы Thyrohyrax и Saghatherium встречаются в обеих формациях, как и примат Moeripithecus . В более общем плане обе формации сохраняют проплиопитециды и парапитециновые парапитециды , а также олигопитециды . В то время как первые две группы ограничены более молодой фаюмской толщей, последняя является наиболее распространенным семейством приматов в более древних отложениях. Если предположить традиционную интерпретацию формации Джебель-Катрани, это будет означать, что многие из этих таксонов должны были появиться в Египте на 2–4 миллиона лет раньше, чем в Омане, что Зейферт считает маловероятным. По их мнению, возраст формации составляет около 8 миллионов лет, а ее древнейшие местонахождения расположены в слоях позднего эоцена. Возраст местности BQ-2 оценивается в 37 миллионов лет (ранний Приабон ), а возраст L-41 попадает в возрастной диапазон 34,8–33,7 миллионов лет, временной интервал, включающий границу эоцена и олигоцена. Хотя Зейферт не смог опровергнуть самый ранний олигоценовый возраст, он утверждает, что поздний эоценовый возраст следует считать более вероятным, основываясь на несогласии, присутствующем чуть выше местности (которое Расмуссен также признал возможным кандидатом на EOB). Это означает, что только нижние 48 м (157 футов) формации имеют эоценовый возраст, включая участки BQ-2 и L-41. Остальная часть нижней толщи, как и вся верхняя толща, впоследствии попадет в олигоцен. Положение карьеров A и B относительно границы неоднозначно, однако, с другой стороны, карьер E, с другой стороны, без сомнения, по Зейферту считается олигоценовым (около 33 млн лет назад). [ 3 ]
Палеосреда
[ редактировать ]
Окружающая среда формации Джебель-Катрани была описана Бауном как низменная равнина от субтропического до тропического климата , который далее предполагает наличие ручьев и прудов. Основываясь на останках ископаемых птиц, которые включают окаменелости различных животных, тесно связанных с водой (скопы, ранние фламинго, яканы, цапли, аисты, бакланы и китоглавы), Расмуссен и его коллеги пришли к выводу, что окружающая среда представляет собой медленно движущуюся пресную воду с значительное количество водной растительности, что соответствует предыдущей гипотезе. Хотя литология предполагает, что большинство окаменелостей отложилось на песчаных отмелях после переноса течениями, авторы утверждают, что болота могли легко образоваться вдоль берегов реки, которая существовала в олигоцене, и могут быть причиной аргиллита, найденного в некоторых карьерах. Кроме того, они предполагают, что ископаемые птицы Фаюма из-за их сходства с современными группами следует считать более ценным индикатором окружающей среды по сравнению с ископаемыми млекопитающими, многие из которых принадлежали к семействам, не имеющим современных образцов. Отсутствие других птиц, типичных для такой среды, можно объяснить либо неточностью выборки, либо тем, что указанные группы просто еще не присутствовали в олигоценовой Африке. В целом Расмуссен и его коллеги сравнивают окружающую среду Джебель-Катрани с пресноводной средой обитания в современной Центральной Африке. [ 1 ] Обнаружение окаменелостей змееголов, похоже, подтверждает интерпретацию Расмуссена, поскольку сегодня род Parachanna предпочитает медленно текущие заводи с обилием растительности. Присутствовавшие в то же время другие рыбы, особенно Tylochromis , позволяют предположить, что также присутствовала глубокая открытая вода. Русла рек, возможно, были заросли тростником , папирусом и украшены плавающей растительностью, такой как кувшинки и сальвиния . [ 4 ] В статье 2001 года Расмуссен и др. утверждал, что песчаник и аргиллит формации, вероятно, образовались в результате разложения отложений системой речных каналов, которые впадали на запад в Тетис. Здесь они реконструировали окружающую среду как тропический низинный болотный лес, смешанный с болотами. Кроме того, они предполагают, что в окружающей среде бывали муссоны . [ 5 ] В целом это указывает на то, что этот регион был частью обширного пояса тропических лесов, который простирался через территорию нынешней Северной Африки, которая постепенно превращалась в открытые леса и даже в степи по мере продвижения вглубь страны. [ 2 ]
Палеобиота
[ редактировать ]Рыба
[ редактировать ]Актиноптеригии
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Alestidae плетет. [ 4 ] | Нижняя и верхняя последовательность | Карьер E, I, M, R & X | Различные зубы | Различные зубы, принадлежащие Characiformes , они общие для обоих ископаемых представителей. В формации присутствует большой диапазон морфотипов зубов, однако это может не отражать фактическое разнообразие присутствующих видов африканских тетра. | ||
| Amiiformes плетет. [ 4 ] | ||||||
| Опоздает [ 4 ] | Лейтс сп. | Нижняя и верхняя последовательность | Карьер E, I, M & V | Два черепа и другой изолированный материал. | Его отсутствие в озерной среде L-41 могло указывать на то, что вода была слишком мелкой. Современные лейты могут вырасти большими и предпочитают большие озера и реки. | |
| Парачанна [ 4 ] | П. фаюменсис | Нижняя и верхняя последовательность | Карьер Л-14, П&М | Различные черепные останки, включая зубную кость. | Змееголовая рыба, материал Джебель-Катрани, может свидетельствовать о том, что эта группа прибыла в Африку раньше, чем предполагалось ранее. | |
| Ткани Siluriformes . [ 4 ] | Нижняя и верхняя последовательность | Карьер L-41, B, G, E, I, J, K, M, P & V | Три черепа и различные посткраниальные останки, преимущественно плавниковые шипы. | Три известных черепа примерно одинакового размера, но могут не представлять один и тот же таксон. Посткраниальные элементы указывают на больший размерный диапазон. В некоторых аспектах сом Джебель-Катрани напоминает род Fajumia . | ||
| ср. Тилохромис [ 4 ] | Нижняя и верхняя последовательность | Карьер E, I и X | Нижняя челюсть и дополнительный материал, включая зубы. | Базальная цихлида . |
Хондриктиес
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Дасьятоидеа индет. [ 4 ] | ||||||
| Ткань Lamniformes . [ 4 ] |
Саркоптеригии
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Дипной интет. [ 4 ] | Карьер Л-41 |
Рептилии
[ редактировать ]| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Альбертвудемис [ 6 ] | А. черепаха | Нижняя последовательность | Местность к западу от карьеров AMNH. | Частичный пластрон с сочлененными периферийными элементами верхней раковины. | Боковая черепаха семейства Podocnemididae с высоким куполом, вероятно, напоминающая черепахи. Это может быть родственный таксон экземпляру UCMP 42008 из миоцена Кении. | |
| Эндрюсемис [ 6 ] [ 7 ] | А. лыбица | К северу от озера Карун | Остались различные оболочки. | Боковая черепаха семейства Podocnemididae, ранее известная как «Stereogenys» lybica . Образец голотипа, один из наиболее хорошо сохранившихся панцирей черепах из Фаюма, считался утерянным, прежде чем его заново открыли и использовали для выделения нового рода. Несколько дополнительных экземпляров известны из Джебель-Катрани и его нижележащих образований. | ||
| Крокодил | С. мегаринус [ 8 ] | Различные черепные и нижнечелюстные останки | Крупный и широкорылый вид крокодилов, ошибочно отнесенный к Crocodylus . Второй фаюмский крокодил, «Crocodylus» articeps , Кристофер Брошу считал более молодой особью этого вида. | |||
| Дакемис [ 6 ] [ 9 ] | Д. палеоморфа | Нижняя последовательность | Карьер Б | Почти полный череп | Боковая черепаха семейства Podocnemididae , которая может быть родственным таксоном экземпляру UCMP 42008. Вполне возможно, что Dacquemys представляет собой материал черепа Albertwoodemys . | |
| Эогавиалис [ 8 ] | Э. африканский | Ранний род гавиалоидов, распространённый в формации. Два вида первоначально были названы от Джебель-Катрани: Eogavialis gavialoides и Eogavialis tenuirostre (оба первоначально назывались Tomistoma ), однако недавние исследования показывают, что все виды Fayum Eogavialis, включая виды из других формаций в регионе, могут просто представлять один вид. [ 2 ] | ||||
| Гиганточерсина [ 10 ] | Г. аммон | Нижняя последовательность | Карьер А и Б | Несколько экземпляров, включая почти полный панцирь и пластрон, а также частичный таз. | Самая старая известная черепаха Африки. Оба названия были Testudo beadnelli и Testudo isis , но с тех пор стали синонимами G. ammon . [ 2 ] | |
| Неохелис [ 6 ] | Н. фаюменсис | Нижняя и верхняя последовательность | Карьер L-41, A, B, C, I, M, O, P & R | Различные остатки пластрона и панциря, а также материал черепа. | Боковая черепаха семейства Podocnemididae, похожая на Erymnochelys и ранее отнесенная к указанному роду. Типовой экземпляр, передний пластрон, потенциально утерян. Они являются одними из наиболее распространенных окаменелостей в карьерах L-41, I и M. | |
| Вараниды [ 2 ] | Неопределенный. | Верхняя последовательность | Карьер I | Пресакральный позвонок | I одиночная кость, описанная Smith et al. и первоначально отнесенный к роду Varanus . Более поздние исследования Холмса и др. предполагает, что это был стволовой варанид, отличный от других останков, обнаруженных в формации. | |
| Варан [ 2 ] | Неопределенный. | Верхняя последовательность | Карьер Я и М | Позвонки | Изолированные кости, описанные Holmes et al. и считается частью современного рода Varanus . Они утверждают, что это самый старый известный представитель рода и указывает на африканское происхождение современных варанов. |
Птицы
[ редактировать ]похожий на ястреба
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Accipitridae indet., aff. Галиет [ 1 ] | Нижняя последовательность | Карьер А | Дистальный конец предплюсны. | Хищная птица, сходная по размерам и морфологии с современными орланами. Ранние слои формации указывают на более близкую близость к берегу, возможно, поддерживая образ жизни, аналогичный современным видам Haliaeetus . | ||
| Pandionidae indet., aff. Пандион [ 1 ] | Верхняя последовательность | Карьер М | Дистальный конец плечевой кости. | Ископаемое, почти идентичное современной скопе , но меньшего размера. | ||
| Пандиониды? салфетка. [ 1 ] | Верхняя последовательность | Карьер I | Поврежденный карпометакарпус. | Хищная птица размером примерно с нынешнюю скопу, но более крепкая. |
Ржанкообразные
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Джанипес [ 1 ] | J. нимфеобаты | Верхняя последовательность | Карьер М | Дистальный конец предплюсны | Крупный вид яканы , превосходящий по размерам все современные таксоны, но все же меньший, чем Nupharanassa bulotorum . Фаюмские яканы уже проявляют признаки образа жизни, подобного образу жизни их современных сородичей, и впоследствии могут указывать на густую плавающую растительность. | |
| Количество быков | Верхняя последовательность | Карьер М | Множественные экземпляры с сохранением предплюсны. | Крупный вид яканы, на 30–35% крупнее самого крупного существующего вида — яканы с бронзовыми крыльями . | ||
| Н. толутария | Нижняя последовательность | Карьер Е | Дистальный конец предплюсны. | Меньший родственник Nupharanassa bulotorum , отличается гораздо меньшими размерами и более старшим возрастом. Это единственная известная якана в нижних слоях формации Джебель-Катрани. |
Ciconiiformes
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Палеоефиппиоринх [ 1 ] | Дитрих П. | Верхняя последовательность | Карьер М | Почти полный рострум с нижней челюстью и частичным черепом, а также дистальный конец правой большеберцовой кости. | Крупный аист, сравнимый по размеру с современными аистами джабиру и марабу . Хотя типовой материал имеет большое сходство с сохранившимся седлоклювом , тибиотарзус, найденный в другом карьере, не имеет сходства ни с одним современным таксоном и может принадлежать, а может и не принадлежать Palaeoephippiorhynchus . |  |
Кукулиформные
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Musophagidae indet., aff. Кринифер [ 1 ] | Верхняя последовательность | Карьер М | Дистальные концы предплюсне-плюсневой кости и плечевой кости. | Относительно крупный тип турако , один из самых ранних зарегистрированных представителей группы. Он имеет сходство с существующим родом Crinifer . |
Груиформы
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Gruidae плетет. [ 1 ] | Верхняя последовательность | Карьер М | Дистальный конец и частичный стержень предплюсны. | Птица, имеющая сходство с современными венценосными журавлями , журавлями рода Grus и хромым журавлями . По размерам он находится между размерами хромого журавля и журавля-красавки , хотя и ближе к первому. | ||
| Раллида надела. [ 1 ] | Верхняя последовательность | Карьер М | Дистальный конец предплюсны. | Возможно, это тип рельсового подшипника, сходный с родами Sarothrura , Coturnicops и Laterallus . |
Палеогнаты
| Имя | Разновидность | Член | Материал | Примечания | Изображение |
|---|---|---|---|---|---|
| Почетная сестра [ 1 ] [ 5 ] | Э. эокаен | Нижняя последовательность | Различные фрагментированные кости ног, включая дистальный конец большеберцовой кости и дистальный конец предплюсны. | Загадочная крупная наземная птица, по размеру сравнимая с современной реей . Первоначально считалось, что это разновидность бескилевых , но его связь с другими птицами неоднократно подвергалась сомнению. Stromeria fajumensis , обнаруженная в той же формации, считается синонимом Eremopezus . |
Пелеканиформы
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Ardeidae индет. [ 1 ] | Верхняя последовательность | Карьер Я и М | Частичный рострум, дистальная часть предплюсны ювенильного экземпляра и первая фаланга третьего пальца стопы. | Цапля среднего размера, сравнимая по размеру с красноватой цаплей и большой белой цаплей . Отсутствие перекрывающегося материала означает, что неизвестно, представляют ли эти окаменелости один или несколько видов. | ||
| Голиафия [ 1 ] | Г. Эндрюси | Нижняя и верхняя последовательность | Карьер М | Полная локтевая кость и дистальный конец предплюсне-плюсневой кости. | Крупный родственник современного китоглава . |  |
| Никтикоракс [ 1 ] | Н. сп. | Верхняя последовательность | Карьер М | Почти целые предплюсны и клювовидный отросток. | Останки птицы описаны как идентичные останкам нынешней ночной цапли с черной короной , что указывает на то, что ночные цапли отделились от других цапель по крайней мере 31 миллион лет назад. | |
| Ксенеродиопс [ 1 ] | X. миктер | Верхняя последовательность | Карьер Я и М | Почти полный рострум и плечевая кость без дистального конца. | Необычная цапля с тяжелым, загнутым вниз клювом. |
Феникоптериформы
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Относится к отряду Phoenicopteridae . Вид 1, аф. Палеолус [ 1 ] | Верхняя последовательность | Карьер М&Я | Множественные экземпляры, включая большеберцовую кость и части пястных костей. | Некоторые кости Fayum phoenicopteriform демонстрируют сходство как с производными фламинго, так и с более базальным Palaeolodus , которого Расмуссен и его коллеги считали Phoenicopterids, а не относили к отдельному семейству. Материал позволяет предположить, что это птица размером примерно с Palaelodus gracilipes . | ||
| Относится к отряду Phoenicopteridae. Вид 2 [ 1 ] | Верхняя последовательность | Карьер М | Фрагмент клювовидного отростка | Второй, более крупный феникоптериформ, по размеру ближе к Palaeolodus crassipes и малому фламинго . |
Сулиформы
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Фалакрокоракиды [ 1 ] | Неопределенный. | Верхняя последовательность | Карьер М | Дистальная предчелюстная кость. | Неопределенный баклан с более постепенно суживающимся и более сильно загнутым клювом. Среди современных бакланов он больше всего напоминает баклана гуанай . |
Млекопитающие
[ редактировать ]Афронасекомоядные
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Эохризохлорис [ 11 ] | Э. трибосфен | Нижняя последовательность | Карьер Е | Крошечный представитель афросорицид . | ||
| Геродот [ 12 ] | Х. паттерсони | Нижняя последовательность | Карьер Л-41 | Окаменелости зубной и верхней челюстей. | Разновидность слоновой землеройки . | |
| Джаухария [ 13 ] [ 11 ] | Дж. тенрекоидес | Нижняя последовательность | Карьер Е | Родственник тенреков и золотых кротов . | ||
| Метолдоботес [ 2 ] | М. стромери | Род слоновой землеройки. | ||||
| Тарнилестес [ 13 ] [ 11 ] | Q. Олигоцен | Верхняя последовательность | Карьер I | Нижняя челюсть плохой сохранности. | Самый производный представитель Afrosoricida в формации. | |
| В. Боуни | Нижняя последовательность | Карьер Л-41 | Стебель-афросорицид. | |||
| В. Расмуссени | Нижняя последовательность | Карьер Л-41 |
Парнокопытные
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Б. Эндрюси | Верхняя последовательность | Л-75 | Остатки зубов и зубных рядов. | Крупный антракотер с самых верхних уровней формации, он, возможно, был наиболее тесно связан с B. fraasi либо посредством анагенеза, либо путем отделения от линии B. fraasi. |
 | |
| Б. фраза | Верхняя последовательность | Карьер I | Вид Bothriogenys, происходящий от B. gorringei . Возможно, он либо разделился на две линии, либо дал начало B. andrewsi в результате анагенеза . | |||
| Б. к красным | Нижняя и верхняя последовательность | Карьер А, Б и М | Первоначально описано на основе нижней челюсти. | Самый старый вид Bothriogenys в Джебель-Катрани, возможно, разделился на две линии, давшие начало B. fraasi и B. Rugulosus . Возможно, это был полуводный браузер или травоядный, питавшийся в основном листвой. Материал, условно отнесенный к этому виду, хотя и типичен для нижней толщи, был обнаружен и в карьере М. | ||
| Б. ругулозус | Верхняя последовательность | Вид Bothriogenys , который, вероятно, произошел от B. gorringei . | ||||
| Наботерий [ 16 ] [ 18 ] [ 2 ] | Нет. Египетский | Частичный череп, несколько окаменелостей нижней и верхней челюстей, а также отдельные зубы. | Антракотер с хорошо развитыми клыками. Его зубной ряд лучше подходит для раздавливания фруктов, а не для разрезания растительности. | |||
| Тарниодон [ 14 ] [ 16 ] [ 19 ] | В. маленький | Нижняя последовательность | Голотип состоит из частичной нижней челюсти. | Самый маленький антракотер из формации Джебель-Катрани, менее распространенный, чем Bothriogenys . |
Рукокрылые
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Дофарелла [ 20 ] [ 21 ] | Д. сигей | Нижняя последовательность | Карьер Л-41 | Левая зубная кость. | Разновидность летучей мыши с ножнами . | |
| Khonsunycteris [ 20 ] | К. aegypticus | Нижняя последовательность | Карьер Л-41 | Левая зубная кость. | Разновидность вечерней летучей мыши . | |
| Фасматониктерис [ 22 ] [ 20 ] | П. фиоменсис | Верхняя последовательность | Карьер I | Стоматология осталась. | Разновидность летучей мыши-присоски . | |
| Филис [ 20 ] [ 21 ] | П. сфингис | Верхняя последовательность | Карьер I | Материал верхней и зубной кости. | мышь Летучая -филисид , родственная старшей Витватии из Биркет-Каруна. | |
| Сахарадерма [ 20 ] | С. псевдовампирус | Нижняя последовательность | Карьер Л-41 | Правильный зубной ряд. | Разновидность ложной летучей мыши-вампира . | |
| После вампиризма [ 20 ] [ 21 ] | V. orientalis | Верхняя последовательность | Большая плечевая кость. | Летучая мышь неопределенного родства. Это была первая ископаемая летучая мышь, обнаруженная в Африке, и самая крупная летучая мышь фаюмской последовательности весом до 120 г (4,2 унции). Это помещает его в диапазон размеров египетской летучей мыши . |
Эмбритоподы
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Арсинотерий [ 23 ] [ 24 ] | А. Зиттели | Нижняя и верхняя последовательность | Чуть выше карьера L-41 и карьера M, охватывая большую часть эпизода. | Крупный арсинотериид, эмбритопод известный своими двумя рогами. Известно большое количество материала, и в прошлом второй вид из Джебель-Катрани, A. andrewsi был предложен . Однако некоторые исследователи считают его синонимом A. zitteli , различия которого, возможно, вызваны половым диморфизмом. |  |
Гиенодонта
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| А. лептогнат | Нижняя последовательность | Карьер А | Тонкая нижняя челюсть. | Член Hyainailourinae , первоначально был описан как Pterodon leptognathus . Он был меньше, чем два других вида «Птеродон» из Джебель-Катрани и Метаптеродон . |  | |
| А. нефертитицион | Нижняя последовательность | Карьер Л-41 | Весь череп, а также небо и зубная кость. | Второй вид Akhnatenavus, меньший по размерам, чем A. leptognathus . Череп также, по-видимому, был намного короче, чем у типового вида. В результате расчетов средняя масса тела составила 19,2 кг (42 фунта), что примерно равно размеру евразийской рыси или росомахи . |  | |
| А. всегда | Нижняя последовательность | Описано по верхней челюсти. | был назван в 1910 году, Хотя A. altidens он не был должным образом описан до 1911 года. К этому виду также относили нижнюю челюсть, но Ланге-Бадре и Бёме сочли это решение сомнительным. | |||
| А. макрогнат | Нижняя последовательность | Карьер А и Б | Черепа и челюсти | Гиенодонт, первоначально описанный как разновидность птеродона . |  | |
| Брихотерий [ 25 ] [ 30 ] | Б. эфалмос | Нижняя последовательность | Карьер Л-41 | Множественные нижние челюсти и части головотрубки. | Тератодонтовый гиенодонт примерно такого же веса, как современная рыжая лисица или американский барсук. Хотя название было придумано в диссертации в 1994 году, официально оно не было описано до 2016 года. |  |
| Фалькатодон [ 31 ] [ 32 ] [ 27 ] | Ф. слесарь | Верхняя последовательность | Карьер V | Описано на основании левой зубной кости. | Выведенный Холройдом как разновидность метаптеродона , уже тогда было отмечено, что он, возможно, отличается от других. Позже Моралес и Пикфорд выделили его в отдельный род. В нижней последовательности может быть дополнительный материал из карьера B, однако отнесение указанных окаменелостей является лишь предварительным. |  |
| М. aegypticum | Верхняя последовательность | Карьер G | Зубные останки | Тератодонтиновый гиенодонт меньше Brychotherium . | ||
| М. Нананубис | Нижняя последовательность | Карьер Л-41 | Череп, нижние челюсти и множественные плечевые кости . | Меньший вид Masrasector из более древних отложений. Его предполагаемый размер тела находился в пределах размеров современных полосатых скунсов и мелкопятнистых генет . Кости конечностей указывают на то, что это был быстрый бегун, который мог охотиться на местных грызунов. | 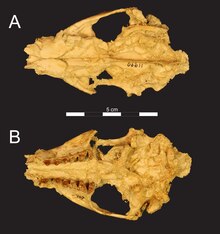 | |
| Метаптеродон [ 26 ] [ 27 ] [ 15 ] | М. брахицефал | Нижняя последовательность | Карьер А | Левая зубная кость. | Названный как вид Hyaenodon в конечном итоге поместил его в Metapterodon , Холройд . Хотя Моралес и Пикфорд ограничивают род миоценом и помещают другие виды Холройда в отдельные роды, они не упоминают M. brachycephalus . |  |
| Метазинопа [ 26 ] [ 25 ] [ 30 ] | М. фраза | Верхняя последовательность | Почти целая нижняя челюсть. | Возможный родственник тератодонтинов, хотя его точная связь с другими гиенодонтидами требует дополнительных исследований. Он был больше, чем Масрасектор . |  | |
| П. африканский | Нижняя последовательность | Карьер А | Мандибулы, трибуна и назначенный шейный позвонок. | Первый гиенодонт, описанный из Фаюма, более поздний анализ показал, что он был гораздо более производным, чем Pterodon dasyuroides , типовой вид. Следовательно, требуется другое название рода. |  | |
| П. фиоменсис | Нижняя последовательность | Карьер А | Нижняя челюсть меньше, чем у «P». африканский . | Как «П». африканский , «П.» phiomensis является гораздо более производным, чем типовой вид Pterodon , и поэтому требует другого названия рода. Более того, анализ показал, что он не был тесно связан с «П». африканский же. Он наиболее тесно связан с кладой, образованной Эхнатенавом , Исохьяенодоном и Хяинаилуросом . |  | |
| П. сп. | Нижняя последовательность | Карьер Е | Сломанный клык. | Изолированный зуб, по размеру похожий на «П». фиоменсис . | ||
| П. сиртос | Верхняя последовательность | Карьер М | Фрагмент правой верхней челюсти. | |||
| Квазиаптеродон [ 29 ] [ 2 ] [ 31 ] [ 33 ] | Q. минута | Верхняя последовательность | Первоначально названный как вид Apterodon наряду с A. altidens , более поздние исследования показали, что он явно отличается от этого рода. Вместо этого это может быть связано с «Sinope» ethiopica . | |||
| Сектизодон [ 31 ] [ 32 ] [ 27 ] | С. маркграфи | Левая верхняя челюсть. | S. markgrafi первоначально был назван Холройдом как вид Metapterodon поместили его в новый род Sectisodon в 1999 году, но в 2017 году Моралес и Пикфорд . | |||
| Синопа [ 34 ] [ 25 ] [ 29 ] [ 35 ] | С. эфиопская | Нижнечелюстная ветвь. | Гиенодонт первоначально описывался как вид Sinopa , хотя более поздние исследования ясно показывают, что он не принадлежит к американо-азиатскому роду. Его точное отношение к другим гиенодонтам остается неопределенным до дальнейшего изучения, однако было высказано предположение, что он является родственником Quasiapterodon . |
Холройд также идентифицировал несколько неопределенных птеродонтовых гиенодонтов из различных последовательностей формации Джебель-Катрани, но не идентифицировал их за пределами уровня подсемейства. [ 27 ]
Гиракоидея
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Антилохиракс [ 36 ] [ 37 ] | А. пептиденс | Нижняя последовательность | Карьер Л-41 | Мандибулы, раздробленный череп, а также различные неполные окаменелости черепа и посткраниальные элементы. | Титаногирацин, похожий на газель, описанный в 2000 году. Зубы и кости конечностей указывают на то, что это был беглый бродяга, похожий на современных антилоп и газелей. | |
| Б. фаюменсис [ 38 ] [ 39 ] [ 40 ] | Нижняя и верхняя последовательность | Описано по нижней челюсти. | Виды Bunohyrax представляли собой гиракоиды среднего размера, несколько напоминающие по зубному ряду свиньи. Возможно, они были родственниками Пахихиракса . | |||
| Б. майор [ 38 ] [ 40 ] | Верхняя последовательность | Описано по трем премолярам. | Первоначально названный Geniohyus major , B. major крупнее B. fajumensis . Кроме того, его зубы более бунодонтные. | |||
| Б. сп. [ 36 ] | Нижняя последовательность | Карьер Л-41 | ||||
| Г. дифик [ 41 ] | Нижняя последовательность | Среднерослый сорт. Гениохиус - один из самых редких гиракоидов Джебель-Катрани. | ||||
| Г. Магнус [ 40 ] [ 41 ] | Нижняя и верхняя последовательность | Карьер V | Различные останки черепа, включая фрагменты верхней челюсти и остатки зубов. | Это самый маленький вид Geniohyus , название которого появилось еще тогда, когда он еще считался частью Saghatherium . Его зубной ряд является промежуточным между другими Geniohyus и Bunohyrax . | ||
| После смерти Г. [ 38 ] [ 40 ] [ 41 ] | Нижняя последовательность | Описано по нижней челюсти. | Самый крупный из трёх названных видов Fayum Geniohyus и типовой вид. | |||
| Г. сп. [ 36 ] | Нижняя последовательность | Карьер Л-41 | Geniohyus получил название вида из-за их морфологического сходства со свиньями, а ранние исследования даже классифицировали их как свиней . | |||
| М. эокенус [ 36 ] [ 38 ] [ 39 ] [ 40 ] | Верхняя последовательность | Описано на основании предчелюстного и верхнечелюстного материала. Также включает остатки нижней челюсти. | Мегалогиракс демонстрирует сильный половой диморфизм в отношении размера тела. Это был общий род в формировании. | |||
| М. сп. [ 36 ] [ 38 ] | Нижняя последовательность | Карьер Л-41 | Безымянный вид Megalohyrax, сохранивший внутреннее нижнечелюстное окно, в отличие от более молодого M. eocaenus . | |||
| Пахихиракс [ 36 ] [ 38 ] [ 39 ] [ 41 ] | П. crassidentatus | Верхняя последовательность | Карьер I, L-46 и М | Черепа, челюсти и зубы, включая полный, но раздробленный череп молодой особи. | Гиракоид, напоминающий современного антракотера Bothriogenys . Возможно, это было полуводное травоядное животное. | |
| Св. древний [ 38 ] [ 43 ] [ 40 ] [ 41 ] | Нижняя последовательность | Описано по верхней челюсти. | У Saghatherium были тяжелые челюсти, и, возможно, он питался орехами и семенами. Кроме того, представители этого рода демонстрируют выраженный половой диморфизм и, возможно, имели сложные брачные ритуалы. Было показано, что вид S. sobrina просто представляет самок этого вида. |
 | ||
| С. плеч [ 41 ] | Верхняя последовательность | Карьер V | Зубные ряды и зубы. | Небольшой вид Saghatherium , отличающийся наличием внутренней камеры нижней челюсти. | ||
| С. боуни [ 36 ] [ 38 ] [ 43 ] | Нижняя последовательность | Карьер Л-41 | Распространенный вид Saghatherium, названный в 1991 году. | |||
| Селеногиракс [ 36 ] [ 38 ] [ 41 ] | С.Чатрати | Верхняя последовательность | Карьер V | Несколько мандибул. | Гиракоид назван в честь своих селенодонтных зубов (коренных зубов с серповидными гребнями), которые гораздо более развиты, чем у Titanohyrax . Эти зубы указывают на диету, которая требовала от них нарезать растительность, а не измельчать ее. Это небольшой даман, лишь немного крупнее Saghatherium , от которого он мог произойти. | |
| Т. заснул [ 38 ] [ 39 ] | Верхняя последовательность | Карьер I, G, L-46, M, P, R, V и X | В первую очередь остатки нижней челюсти и зубы, а также некоторые элементы конечностей и позвонков. | Тирохиракс является одним из самых мелких гиракоидов Фаюма и напоминает современного древесного дендрохиракса . Виды Thyrohyrax демонстрируют небольшой половой диморфизм, связанный с размером, однако самцы обладают увеличенными, похожими на бивни вторыми резцами и челюстями с камерами. | ||
| Т. литолагус [ 36 ] [ 43 ] | Нижняя последовательность | Карьер Л-41 | Родственник младшего Thyrohyrax pygmaeus , он был несколько крупнее T. meyeri . | |||
| Т. Мейер [ 36 ] [ 38 ] [ 43 ] | Нижняя последовательность | Карьер Л-41 | T. meyeri был очень похож на более молодого T. domorictus и, возможно, был его близким родственником. | |||
| Т. пигмей [ 38 ] [ 41 ] | Нижняя последовательность | Карьер А-1 | Передняя часть черепа с прилегающей нижней челюстью, а также несколько зубов от другого экземпляра. | Близкий родственник прямого потомка T.litholagus . Первоначально он был отнесен к Saghatherium magnum , затем описан как разновидность Megalohyrax, а затем переведен в Pachyhyrax и, в конечном итоге, в Thyrohyrax . | ||
| Т. Эндрюси [ 36 ] [ 40 ] [ 41 ] | Нижняя последовательность | Пять экземпляров, включая две челюсти и одну верхнюю челюсть. | Виды Titanohyrax являются крупнейшими гиракоидами Фаюмской впадины и образуют кладу Titanohyracinae с Antilohyrax . Считается, что, как и их меньший родственник, они были фоливорами. Нижние резцы T. andrewsi имели форму лопаточки, а не бивней. | |||
| Т. сужение [ 36 ] [ 40 ] [ 41 ] | Верхняя последовательность | Карьер V, R&I | Восемь экземпляров, включая остатки верхней, предчелюстной и нижней челюсти. | T. angustidens имеет наиболее сложную историю среди видов Titanohyrax , первоначально отнесенных к Megalohyrax и получивших название M. palaeotheroides , позже он был признан Titanohyrax и по-разному считался отдельным или синонимом T. andrewsi . Название T. palaeotheroides не сохранилось, так как это было nomen nudum . | ||
| Т. последний [ 36 ] [ 40 ] | Верхняя последовательность | Четыре сильно изношенных зуба. | Самый крупный вид Titanohyrax , оценка веса T. ultimus колеблется в пределах 600–1300 кг (1300–2900 фунтов), что делает его, возможно, таким же большим, как современный суматранский носорог . | |||
| Т. сп. [ 36 ] | Нижняя последовательность | Карьер Л-41 | Безымянный вид из карьера L-41. |
Марсупалия
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Ператерий [ 2 ] [ 44 ] | П. африканский | Верхняя последовательность | Карьер М | Несколько экземпляров, включая верхнюю и три нижних челюсти. | Примитивное сумчатое животное и самое базальное млекопитающее, встречающееся в отложениях свиты. Одно время он был известен под названием Катранитерий , однако с тех пор это имя считается младшим синонимом. |
Грызуны
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| А. до свидания | Нижняя последовательность | Карьер А, Б и Е | Этот вид также включает несколько экземпляров, ранее отнесенных к Phiomys andrewsi . Однако, поскольку оно не было адекватно описано, оно считается nomen nudum . | |||
| А. боуни | Нижняя последовательность | Карьер Л-41 | Полный верхний и нижний зубной ряд, а также части нижней челюсти и черепа. | Базальный представитель Phiomorpha и самый крупный представитель семейства Phiomyidae , хотя сейчас эта группа считается парафилетической . Это единственный вид Acritophiomys , который был правильно назван, поскольку и A. adaios , и A. woodi были описаны только в диссертации . | ||
| А. существует | Нижняя последовательность | Карьер Л-41 | Первый фиомидид, описанный из карьера L-41. Однако, поскольку оно не было адекватно описано, оно считается nomen nudum. | |||
| Биркамыс [ 45 ] [ 2 ] [ 47 ] | Б. ранний | Нижняя последовательность | Карьер Л-41 | Краниальные и нижнечелюстные останки с зубами, включая частично сохранившийся рострум. | Близкий родственник Мубхаммиса . | |
| Г. аегиптий | Нижняя последовательность | Карьер Е | Мандибулы и зубы | Типовой вид рода, описанный Вудом в 1960-х гг. По размеру он промежуточный среди фаюмских грызунов, крупнее маленького Phiocricetomys , но меньше Metaphiomys . Его филогения загадочна, и его по-разному относили к сестре дикобразов Старого Света или к Caviomorpha . | ||
| Г. Аслиус | Нижняя последовательность | Карьер Л-41 | Искаженный череп и различные фрагментарные останки. | Он демонстрирует несколько примитивных черт и может напоминать предковую форму этого рода. | ||
| Г. гилей | Нижняя последовательность | Карьер Л-41 | Уплощенный череп и различные дополнительные фрагменты. | G. hylaeus был первоначально назван Холройдом в докторской диссертации, что делало его nomen nudum до полного описания, написанного Салламом, Зейфертом и Саймонсом. | ||
| М. Биднелли | Верхняя последовательность | Левая нижняя челюсть и дополнительный материал. | Фиомидный грызун. | |||
| М. Шауби | Второй вид Metaphiomys, описанный Вудом в 1960-х годах. | |||||
| Монамис [ 46 ] [ 2 ] [ 49 ] | М. Саймонс | Верхняя последовательность | Карьер Я и М | Остатки нижней челюсти и зубы. | Ранее известный как Paraphiomys simonsi , более поздние исследования показали, что этот род отличается от гораздо более позднего миоценового Paraphiomys . | |
| Мубхаммис [ 2 ] [ 47 ] | М. Вадуменсис | Нижняя последовательность | Карьер Л-41 | Краниальные и нижнечелюстные элементы с зубцами. | Близкий родственник Биркамыса . | |
| Фиокрикетомия [ 45 ] [ 46 ] | П. минута | Верхняя последовательность | Карьер I | Нижняя челюсть с сохранением зубного ряда от 4-го временного премоляра до 2-го моляра. | Самый молодой известный представитель Phiocricetomyinae . | |
| П. Эндрюси | Нижняя последовательность | Карьер А и Б | Описано на основании частичных челюстей. | Фиомидный грызун. | ||
| П. парафиомиоидес | Фиомид, описанный Вудом в 1960-х годах. | |||||
| Таранимис [ 45 ] | Q. сафрутус | Нижняя последовательность | Карьер Л-41 | Остался череп и зубы. | Миниатюрный фиокристомииновый грызун. | |
| Талахфиомис [ 45 ] [ 46 ] [ 50 ] | Т. работал | Нижняя последовательность | Карьер Е | Временные премоляры и моляры. | Первоначально считалось, что это разновидность Phiomys , в докторской диссертации 1994 года он был назван Elwynomys . В соответствии с правилами ICZN , название Talahphiomys позже имело приоритет. |
Фолидота
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Фолидота [ 2 ] | Верхняя последовательность | Карьер М&Л-12 | Две кости пальцев. | Фрагментарные останки того, что могло быть ящером. Наличие термитников со следами раскопок может подтвердить это, однако идентичность этих костей пальцев не является общепризнанной и подвергается сомнению. |
Птолемаида
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| К.айшае | Верхняя последовательность | Карьер V | Остатки нижней челюсти и зубы. | Птолемаида в придачу. Типовой вид Клеопатродона . | ||
| С. робуста | Верхняя последовательность | Карьер I | Остатки нижней челюсти и зубы. | У C. Robusta были более крепкие челюсти, чем у C. ayeshae . | ||
| П. гранджери [ 51 ] | Верхняя последовательность | Карьер V | Множественные зубы и ветви нижней челюсти, а также почти полный, но искаженный череп. | Плотоядное афротерийное млекопитающее семейства Ptolemaiidae. Он крупнее старшего P. lyonsi . | ||
| П. лионси [ 53 ] | Нижняя последовательность | Карьер А | Материал нижней челюсти. | Плотоядный афротер, впервые описанный Осборном в 1908 году. | 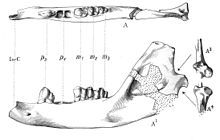 | |
| Карунавус [ 51 ] [ 52 ] | К. Мейери | Нижняя последовательность | Карьер А | Материал нижней челюсти подростка. | Птолемаида в придачу. Несмотря на более молодое животное, материал Q. meyeri крупнее материала Ptolemaia lyonsi , известного из того же местонахождения. Саймонс и Расмуссен предполагают, что это могло быть всеядное животное, похожее на енота. [ 2 ] |
Приматы
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Египтопитек [ 54 ] [ 2 ] | А. Зевксис | Верхняя последовательность | Фрагменты нижней челюсти и целые черепа мужчин и женщин. | Проплиопитецидный . примат из верхних толщ свиты Это может быть синонимом Проплиопитека . |  | |
| Афрамоний [ 2 ] | А. диидес | Нижняя последовательность | Карьер Л-41 | Три частичные челюсти. | Адаптивный примат | |
| Афротарсиус [ 54 ] | А. Чатрати | Верхняя последовательность | Карьер М | Единственная нижняя челюсть. | Примат спорной классификации. Ориентировочная масса тела составляла 130–300 г (4,6–10,6 унций). | |
| Анхомомис [ 3 ] [ 2 ] | А. валы | Нижняя последовательность | Карьер Л-41 | |||
| А. боуни | Верхняя последовательность | Карьер V |
 | |||
| А. Мустафаи | Верхняя последовательность | Карьер G | ||||
| А. фиоменсе | Верхняя последовательность | Различные останки, включая кости черепа и конечностей. | Примат-парапитецид, известный по богатой летописи окаменелостей. Apidium phiomense — типовой вид и один из первых приматов, описанных в Джебель-Катрани. | |||
| Арсиноя [ 55 ] [ 56 ] | А. дорогой | Нижняя последовательность | Карьер Л-41 | Нижняя челюсть с зубами | Парапитецидный примат. | |
| катопитек [ 55 ] [ 54 ] [ 57 ] | К. брауни | Нижняя последовательность | Карьер Л-41 | Более 16 экземпляров, включая черепа, челюсти и посткраниальные останки. | Очень распространенный вид олигопитецидов . По размеру он был похож на крупных приматов -каллитрихид и на более мелких ночных обезьян , беличьих обезьян и обезьян-тити . Его зубы предполагают диету из листьев. |  |
| Олигопитек [ 54 ] | О. саважей | Нижняя последовательность | Карьер Е | Частичная получелюсть и изолированные зубы. | Более редкий вид олигопитецидов. Представители этого семейства практически отсутствуют в верхней толще свиты, что может быть отсроченным результатом изменения климата в период эоцен-олигоценового перехода. Зубы олигопитека указывают на то, что он мог быть плодоядным. | |
| Олигопитециды [ 54 ] | Неопределенный. | Верхняя последовательность | Карьер М | Левая полунижняя челюсть с альвеолами для всех зубов и одним, почти целым коренным зубом. | Самый маленький известный олигопитецид и один из самых маленьких антропоидов. По массе тела он сравним с современными игрунками рода Callithrix и, как было отмечено, меньше толстохвостого карликового лемура . | |
| Парапитек [ 54 ] [ 2 ] | П. фраза | Верхняя последовательность | Полная нижняя челюсть. | Парапитецидный примат. Было предложено использовать синоним Apidium , однако эта точка зрения не является общепринятой. | ||
| Плезиопитек [ 56 ] | П. терраса | Нижняя последовательность | Карьер Л-41 | Нижняя челюсть и почти полный череп. | Загадочный примат, похожий на лемура, принадлежащий к семейству Plesiopithecidae . Это может быть связано с современным ай-ай . | |
| П. анкели | Верхняя последовательность | Карьер V | Стоматологический материал. | Проплиопитецидный примат из верхних толщ свиты. Moeripithecus и Aelopithecus считаются синонимами этого таксона. | 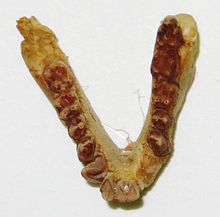 | |
| П. геккели | Множественные останки. | |||||
| Протеопитек [ 57 ] [ 2 ] | П. сильвия [ 55 ] | Нижняя последовательность | Карьер Л-41 | Фрагмент левой верхней челюсти и дополнительные останки, включая частичный череп. | Примат -протеопитецид . | |
| Q. базиодонтос | Нижняя последовательность | Карьер Л-41 | Нижнечелюстные останки | Первоначально описывался как Abuqatrania basiodontos . | ||
| К. Флигли | Верхняя последовательность | Карьер М | Фрагментарная полумандибула. | Примат-парапитецид с предполагаемой массой тела 300–500 г (11–18 унций). | ||
| В. множественное число | Нижняя последовательность | Карьер Е | Сохранились фрагментарные детали зубов. | Маленький примат-парапитецид весом менее 300 г (11 унций). | ||
| Серапия [ 55 ] [ 2 ] | С. эокена | Нижняя последовательность | Карьер Л-41 | Частичная нижняя челюсть | Примат-протеопитецид. | |
| Симонсиус [ 2 ] | С. гранджери | Верхняя последовательность | Карьер I | Фрагмент нижней челюсти. | Спорный род приматов-протеопитецидов. Первоначально он был назван разновидностью Parapithecus , но позже был выделен в отдельный род. Однако это не является общепринятым. |  |
| Вадилемур [ 2 ] | В. Элеганс | Нижняя последовательность | Карьер Л-41 | Возможен лорисиформ . |
Хоботки
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Меритериум [ 2 ] | Mo. trigodon | Правая ветвь нижней челюсти и другие остатки. | Небольшой, возможно, полуводный хоботок размером примерно с тапира. У M. trigodon есть некоторые особенности зубного ряда, которые могут указывать на то, что это может быть род, отличный от Moeritherium. Другой вид также был назван по формации, M. andrewsi , но современные исследования резко сократили количество действительных видов, и большинство из них теперь отнесены к типовому виду M. trigodon . |  | ||
| П. биднелли | Нижнечелюстная остается. | Ранний хоботок, названный Эндрюсом в 1901 году. Это типовой вид рода Palaeomastodon . В отличие от Moeritherium , палеомастодон и фиомия считаются наземными животными. |
 | |||
| П. минор | Вид, отличающийся от P. beadnelli меньшими размерами. Статус большинства видов палеомастодонов сложен и неоднократно обсуждался. | |||||
| П. маленький | Еще один небольшой вид Палеомастодона . | |||||
| П. Винтони | Морфологически сходный с P.minor, он был немного крупнее, но не приближался к размеру типового вида. Это самый распространенный вид в формации. | |||||
| Фиомия [ 2 ] | П. серриденс | Различные останки | Хотя в прошлом было названо несколько видов, Phiomia serridens в настоящее время является единственным действующим видом этого рода, произрастающим в Фаюме. Виды Palaeomastodon иногда также относили к Phiomia . |  |
Сирения
| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| Эосирен [ 59 ] [ 60 ] | Э. менты | Верхняя последовательность | Карьер О | Череп и ребра. | E. imenti является наиболее производным представителем рода Eosiren и имел удлиненную голову. |  |
Флора
[ редактировать ]| Имя | Разновидность | Член | Местность | Материал | Примечания | Изображение |
|---|---|---|---|---|---|---|
| бомбакоксилон [ 61 ] | Б. скала | Ископаемая древесина | « Бомбаковые ». | |||
| Терминалиоксилон [ 61 ] | Т. промежуточный | Ископаемая древесина | Белый мангровый лес . | |||
| Т. оригинал | Ископаемая древесина | Белый мангровый лес . | ||||
| Детариоксилон [ 61 ] | Д. aegyptiacum | Ископаемая древесина | Бобовые . | |||
| Легуминоксилон [ 61 ] | Л. сп. | Ископаемая древесина | Бобовые . | |||
| Фикоксилон [ 61 ] | Ф. бланкенхорни | Ископаемая древесина | Рис . | |||
| пальмоксилон [ 61 ] | П. ашерсони | Ископаемая древесина | Пальма . | |||
| П. геометрический | Ископаемая древесина | Пальма . | ||||
| П. лакуносум | Ископаемая древесина | Пальма . | ||||
| П. либикум | Ископаемая древесина | Пальма . | ||||
| П. пондичерриенсе | Ископаемая древесина | Пальма . | ||||
| Сапиндоксилон [ 61 ] | С. стромери | Ископаемая древесина | ягода Мыльная . | |||
| Стеркулиоксилон [ 61 ] | С. гиарабубенсе | Ископаемая древесина | « Стеркулиевые ». | |||
| Тамарикоксилон [ 61 ] | Т. африканский | Ископаемая древесина | Тамариск . | |||
| Фрагмиты [ 61 ] | П. южный | Окремненные стебли. | Тростник обычно встречается в сочетании с тамарикойлоном . | |||
| Аккростихум [ 61 ] | А. сп. | Окаменелые листья. | Папоротник Polypodiaceae . | |||
| Стенохлена [ 61 ] | С. сп. | Окаменелые листья. | Папоротник Polypodiaceae . | |||
| Сальвиния [ 61 ] | С. сп. | Окаменелые листья. | Папоротник сальвиниевые . | |||
| Он кладет на ладони . [ 61 ] | Ген. ты спишь салфетка. | Окаменелые листья. | Пальма . | |||
| Тифоидная ткань. [ 61 ] | Ген. ты спишь салфетка. | Окаменелые листья. | Рогоз, похожий на Typha. род | |||
| Кости [ 61 ] | Л.энгельхардти | Окаменелые листья. | Лавр . | |||
| кинометр [ 61 ] | К. сп. | Окаменелые листья. | Бобовые . | |||
| Фикус [ 61 ] | Ф. лейкоптероидес | Окаменелые листья. | Рис . | |||
| Ф. стромеры | Окаменелые листья. | Рис . | ||||
| Маэса [ 61 ] | М. Зиттели | Окаменелые листья. | Мирсин . | |||
| Нелумбо [ 61 ] | Н. сп. | Окаменелые листья. | Лотос . | |||
| ? нимфеиты [ 61 ] | ? Н. сп. | Окаменелые листья. | Водяная лилия . | |||
| Охновые ткани. [ 61 ] | Ген. ты спишь салфетка. | Окаменелые листья. | Охновые . | |||
| Относится к семейству Сапотовые . [ 61 ] | Ген. ты спишь салфетка. | Окаменелые листья. | Сапотовые . | |||
| Триплохитон [ 61 ] | Т. сп. | Окаменелые листья. | Стеркулиевые . | |||
| Двудольные [ 61 ] | Ген. ты спишь салфетка. | Окаменелые листья. | Семь неопределенных типов двудольных . | |||
| Эпипремнум [ 61 ] | Э. сп. | Ископаемые плоды. | Арум . | |||
| Он кладет на ладони . [ 61 ] | Ген. ты спишь салфетка. | Ископаемые плоды. | Пальмовый плод . | |||
| принадлежит к семейству Анакардиевые . [ 61 ] | Ген. ты спишь салфетка. | Ископаемые плоды. | Кешью . | |||
| Анонаспермум [ 61 ] | А. виды. | Ископаемые плоды. | Два вида сметанного яблока . | |||
| Канариум [ 61 ] | К. сп. | Ископаемые плоды. | Плод благовонного дерева . | |||
| Икациникария [ 61 ] | И. сп. | Ископаемые плоды. | Белая груша . | |||
| Эогипсера [ 61 ] | Э. сп. | Ископаемые плоды. | семя Лунное . | |||
| Секуридака [ 61 ] | С. терциария | Ископаемые плоды. | Молочница . | |||
| Тамарикс [ 61 ] | Т. сп. | Силицифицированные корни. | Тамариск . | |||
| Макроводоросли [ 61 ] | Ген. и спать салфетка. | Ископаемые таллоиды . | Водоросли . |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в Расмуссен, Д.Т.; Олсон, СЛ; Саймонс, Э.Л. (1987). «Ископаемые птицы из олигоценовой формации Джебель-Катрани, провинция Фаюм, Египет» . Смитсоновский вклад в палеобиологию . 62 (62): 1–20. дои : 10.5479/si.00810266.62.1 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь Кампуридис, П.; Хартунг, Дж.; Огюстен, Ф.Дж. (2023). «Эоцен-олигоценовые комплексы позвоночных Фаюмской впадины, Египет». Фанерозойская геология и природные ресурсы Египта . Достижения в области науки, технологий и инноваций. стр. 373–405. дои : 10.1007/978-3-030-95637-0_14 . ISBN 978-3-030-95636-3 .
- ^ Jump up to: а б с Зайферт Эрик Р. (январь 2006 г.). «Пересмотренные оценки возраста фауны млекопитающих позднего палеогена Египта и Омана» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (13): 5000–5005. Бибкод : 2006PNAS..103.5000S . дои : 10.1073/pnas.0600689103 . ПМЦ 1458784 . ПМИД 16549773 .
- ^ Jump up to: а б с д и ж г час я дж Мюррей, AM (2004). «Позднеэоценовый и раннеолигоценовый костистые кости и связанная с ними ихтиофауна формации Джебель-Катрани, Фаюм, Египет» . Палеонтология . 47 (3): 711–724. Бибкод : 2004Palgy..47..711M . дои : 10.1111/j.0031-0239.2004.00384.x . S2CID 140627361 .
- ^ Jump up to: а б Расмуссен, Д.Т.; Саймонс, Эл.; Хертель, Ф.; Джадд, А. (2001). «Задняя конечность гигантской наземной птицы из верхнего эоцена, Фаюм, Египет» . Палеонтология . 44 (2): 325–337. Бибкод : 2001Palgy..44..325R . дои : 10.1111/1475-4983.00182 . S2CID 130033734 .
- ^ Jump up to: а б с д Гаффни, ES; Мейлан, Пенсильвания; Вуд, Колорадо; Саймонс, Э.; де Алмейда Кампос, Д. (2011). «Эволюция бокошеих черепах: семейство Podocnemididae». Бюллетень Американского музея естественной истории . 350 : 1–237. дои : 10.1206/350.1 . hdl : 2246/6110 . S2CID 83775718 .
- ^ Перес-Гарсия, А. (2017). «Новая информация и установление нового рода египетской палеогеновой черепахи «Stereogenys» libyca (Podocnemididae, Erymnochelyinae)». Историческая биология . 31 (3): 1–10. дои : 10.1080/08912963.2017.1374383 . S2CID 90915135 .
- ^ Jump up to: а б Брочу, Калифорния; Джинджерич, П.Д. (2000). «Новый томистомовый крокодил из среднего эоцена (бартона) Вади-Хитан, провинция Фаюм, Египет» (PDF) . Материалы из Музея палеонтологии Мичиганского университета . 30 (10): 251–268.
- ^ Гаффни, ES; Дембье, Д.Д.; Саймонс, Эл.; Санчес-Вильягра, MR; Мейлан, Пенсильвания (2002). «Переописание черепа Даккемиса, подокнемидидной бокошейной черепахи из позднего эоцена Египта » Новитаты Американского музея (372): 1–16. doi : 10.1206/0003-0082(2002)372<0001:ROTSOD>2.0.CO;2 . S2CID 55263374 .
- ^ Холройд, Пенсильвания; Пархэм, Дж. Ф. (2003). «Древность африканских черепах». Журнал палеонтологии позвоночных . 23 (3): 688–690. Бибкод : 2003JVPal..23..688H . дои : 10.1671/1870 . S2CID 86348004 .
- ^ Jump up to: а б с д Зайферт, ER (2010). «Самые старые и самые молодые записи плацентарных афросорицидов из Фаюмской впадины на севере Египта» . Acta Palaeontologica Polonica . 55 (4): 599–616. дои : 10.4202/app.2010.0023 . S2CID 910171 .
- ^ Саймонс, Эл.; Холройд, Пенсильвания; Баун, ТМ (1991). «Раннетретичные землеройки-слоны из Египта и происхождение Macroscelidea» . Труды Национальной академии наук . 88 (21): 9734–9737. Бибкод : 1991PNAS...88.9734S . дои : 10.1073/pnas.88.21.9734 . ПМК 52794 . ПМИД 11607230 .
- ^ Jump up to: а б с Пикфорд, М. (2019). «Крошечные Tenrecomorpha (Млекопитающие) из эоцена Черной Вороны, Намибия» (PDF) . Сообщения Геологической службы Намибии . 21 : 15–25.
- ^ Jump up to: а б Силим, Ах; Саллам, HM; Хевайди, AGA; Ганнелл, Г.Ф.; Миллер, ER (2015). «Anthracotheres (Mammalia, Artiodactyla) из самого верхнего горизонта формации Джебель-Катрани, последний ранний олигоцен, Фаюмская впадина, Египет» . Египетский журнал палеонтологии . 15 : 1–11. ISSN 1687-4986 .
- ^ Jump up to: а б с Холройд, Пенсильвания; Саймонс, Эл.; Баун, ТМ; Полли, PD; Краус, MJ (1996). «Новые находки наземных млекопитающих из формации Каср-эль-Сагха верхнего эоцена, Фаюмская впадина, Египет» (PDF) . Палеовертебрата . 25 (2–4): 175–192.
- ^ Jump up to: а б с Силим, Ах; Абу Эль-Хейр, Джорджия (2022 г.). «Полный череп Bothriogenys fraasi (Mammalia, Artiodactyla, Anthracotheriidae) из раннего олигоцена, Фаюм, Египет » Геологический журнал . 57 (11): 4833–4841. дои : 10.1002/gj.4574 . S2CID 252244244 .
- ^ О'Лири, Массачусетс; Патель, бакалавр; Коулман, Миннесота (2012). «Эндокраниальная каменистая анатомия Bothriogenys (Mammalia, Artiodactyla, Anthracotheriidae), а также сравнение объема и плотности каменистых костей среди водных и наземных парнокопытных и внешних групп» . Журнал палеонтологии . 86 (1): 44–50. Бибкод : 2012JPal...86...44O . дои : 10.1666/10-091.1 . S2CID 85238250 .
- ^ Силим, Ах; Саллам, HM; Хевайди, AGA; Миллер, скорая помощь; Ганнелл, Г.Ф. (2016). «Новый антракотерий (парнокопытные) из раннего олигоцена, Фаюм, Египет, и разгадка тайны африканского рагатерия». Журнал палеонтологии . 90 (1): 170–181. Бибкод : 2016JPal...90..170S . дои : 10.1017/jpa.2016.13 . S2CID 132618649 .
- ^ Патрисия А. Холройд, Фабрис Лихоро, Грегг Ф. Ганнелл и Эллен Р. Миллер: Anthracotheriidae. В: Ларс Верделин и Уильям Джозеф Сандерс (Hrsg.): Кайнозойские млекопитающие Африки. Калифорнийский университет Press, Беркли, Лос-Анджелес, Лондон, 2010, S. 843–851.
- ^ Jump up to: а б с д и ж Симмонс, Северная Каролина; Зейферт, ER; Ганнелл, Г.Ф. (2016). «Новое семейство крупных всеядных летучих мышей (Mammalia, Chiroptera) из позднего эоцена Фаюмской впадины, Египет, с комментариями по использованию названия «Eochiroptera» » (PDF) . Американский музей Novitates (3857): 1–43. дои : 10.1206/3857.1 . ISSN 0003-0082 . S2CID 87290447 .
- ^ Jump up to: а б с Ганнелл, Г.Ф.; Саймонс, Эл.; Зайферт, ER (2008). «Новые летучие мыши (Mammalia: Chiroptera) из позднего эоцена и раннего олигоцена, Фаюмская впадина, Египет». Журнал палеонтологии позвоночных . 28 (1): 1–11. doi : 10.1671/0272-4634(2008)28[1:nbmcft]2.0.co;2 . S2CID 86125632 .
- ^ Ганнелл, Г.Ф.; Симмонс, Северная Каролина; Зайферт, ER (2014). «Новые Myzopodidae (Chiroptera) из позднего палеогена Египта: уточненный диагноз семейства и биогеографическое происхождение Noctilionoidea» . ПЛОС ОДИН . 9 (2): е86712. Бибкод : 2014PLoSO...986712G . дои : 10.1371/journal.pone.0086712 . ПМЦ 3913578 . ПМИД 24504061 .
- ^ Суд, Н. (1990). «Периотическая анатомия Arsinoitherium (Mammalia, Embrithopoda) и ее филогенетическое значение». Журнал палеонтологии позвоночных . 10 (2): 170–182. Бибкод : 1990JVPal..10..170C . дои : 10.1080/02724634.1990.10011806 .
- ^ Виалле, Н.; Мерзеро, Г.; Делмер, К.; Файст, М.; Жикель, С.; Мариво, Л.; Рамдаршан, А.; Виани-Лио, М.; Эссид, ЭМ; Марзуги, В.; Аммар, Гонконг; Табусе, Р. (2013). «Открытие млекопитающего эмбритопода (Arsinoitherium?) в позднем эоцене Туниса » Журнал африканских наук о Земле . 87 : 86–92. Бибкод : 2013JAfES..87...86В . doi : 10.1016/j.jafrearsci.2013.07.010 .
- ^ Jump up to: а б с д и ж Бортс, MR; Холройд, Пенсильвания; Зайферт, ER (2016). «Гиайнайлориновый и тератодонтиновый черепной материал из позднего эоцена Египта и применение экономных и байесовских методов к филогении и биогеографии Hyaenodonta (Placentalia, Mammalia)» . ПерДж . 4 : е2639. дои : 10.7717/peerj.2639 . ПМК 5111901 . ПМИД 27867761 .
- ^ Jump up to: а б с д и Осборн, HF (1909). «Новые хищные млекопитающие из Фаюмского олигоцена, Египет» (PDF) . Бюллетень Американского музея естественной истории . 26 : 415–424.
- ^ Jump up to: а б с д и ж Холройд, Пенсильвания (1999). «Новые Pterodontinae (Creodonta: Hyaenodontidae) из формации Джебель-Катрани позднего эоцена-раннего олигоцена, провинция Фаюм, Египет» . ПалеоБиос . 19 (2): 1–18.
- ^ Саймонс, Эл.; Джинджерич, П.Д. (1976). «Новый вид аптеродона (Mammalia, Creodonta) из верхнеэоценовой формации Каср-эль-Сагха в Египте» . Постилла Музея естественной истории Пибоди .
- ^ Jump up to: а б с Ланге-Бадре, Б.; Бёме, М. (2005). «Apterodon intermedius, sp. nov., новое европейское млекопитающее-креодонт из MP22 Эспенхайна (Германия)» . Анналы палеонтологии . 91 (4): 311–328. Бибкод : 2005AnPal..91..311L . дои : 10.1016/j.annpal.2005.08.001 .
- ^ Jump up to: а б с д и Бортс, MR; Зайферт, ER (2017). «Краниодентальная и плечевая морфология нового вида Masrasector (Teratodontinae, Hyaenodonta, Placentalia) из позднего эоцена Египта и двигательное разнообразие гиенодонтов» . ПЛОС ОДИН . 12 (4): e0173527. Бибкод : 2017PLoSO..1273527B . дои : 10.1371/journal.pone.0173527 . ПМК 5396875 . ПМИД 28422967 .
- ^ Jump up to: а б с д Бортс, MR; Стивенс, Нью-Джерси (2017). «Первый гиенодонт из позднеолигоценовой формации Нсунгве в Танзании: палеоэкологические взгляды на палеоген-неогеновый переход хищных животных» . ПЛОС ОДИН . 12 (10): e0185301. Бибкод : 2017PLoSO..1285301B . дои : 10.1371/journal.pone.0185301 . ПМК 5636082 . ПМИД 29020030 .
- ^ Jump up to: а б Моралес, Дж.; Пикфорд, М. (2017). «Новые гиенодонты (Ferae, Mammalia) из раннего миоцена Напака (Уганда), Кору (Кения) и Грилленталя (Намибия)». Ископаемый отпечаток . 73 (3–4): 332–359. дои : 10.2478/if-2017-0019 . hdl : 10261/195968 . S2CID 31350436 .
- ^ Соле, Ф.; Люлье, Дж.; Адачи, М.; Бенсала, М.; Махбуби, М.; Табусе, Р. (2014). «Hyaenodontidans из района Гоур-Лазиб (? Ранний эоцен, Алжир): выводы относительно систематики и происхождения Hyainailourinae и Teratodontinae» . Журнал систематической палеонтологии . 12 (3): 303–322. дои : 10.1080/14772019.2013.795196 . S2CID 84475034 .
- ^ Эндрюс, CW (1906). «Описательный каталог третичных позвоночных Фаюма, Египет» . Орден Попечителей Британского музея .
- ^ Морло, М.; Бастл, К.; Вэньхао, В.; Шааль, СФК (2014). «Первый вид Sinopa (Hyaenodontida, Mammalia) за пределами Северной Америки: значение для истории рода в эоцене Азии и Северной Америки» . Палеонтология . 57 (1): 111–125. Бибкод : 2014Palgy..57..111M . дои : 10.1111/пала.12052 . S2CID 129633205 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Расмуссен, Д.Т.; Саймонс, Э.Л. (2000). «Экоморфологическое разнообразие палеогеновых гиракоидов (млекопитающих): новый беглый обзор из Фаюма, Египет» . Журнал палеонтологии позвоночных . 20 (1): 167–176. doi : 10.1671/0272-4634(2000)020[0167:EDAPHM]2.0.CO;2 . S2CID 85172473 .
- ^ Де Блиё, Д.Д.; Саймонс, Э.Л. (2002). «Черепная и зубная анатомия Antilohyrax pectidens: гиракоид позднего эоцена (Mammalia) из Фаюма, Египет». Журнал палеонтологии позвоночных . 22 (1): 122–136. doi : 10.1671/0272-4634(2002)022[0122:CADAOA]2.0.CO;2 . S2CID 87901588 .
- ^ Jump up to: а б с д и ж г час я дж к л Де Блиё, Д.Д.; Баумринд, MR; Саймонс, Э.Л.; Чатрат, PS; Мейер, GE; Аттиа, Ю.С. (2006). «Половой диморфизм внутренней нижнечелюстной камеры Fayum Pliohyracidae (Mammalia)» . Журнал палеонтологии позвоночных . 26 (1): 160–169. doi : 10.1671/0272-4634(2006)26[160:SDOTIM]2.0.CO;2 . S2CID 85657104 .
- ^ Jump up to: а б с д и Мейер, GE (1973). «Новый даман олигоцена из формации Джебель-эль-Катрани, Фаюм, Египет» (PDF) . Музей естественной истории Пибоди, Йельский университет .
- ^ Jump up to: а б с д и ж г час я Табусе, Р. (2016). «Нижняя челюсть гиракоидного млекопитающего Titanohyrax andrewsi в коллекции Национального музея естественной истории, Париж (Франция) с переоценкой вида» . Палеовертебрата . 40 (1): e4. дои : 10.18563/pv.40.1.e4 .
- ^ Jump up to: а б с д и ж г час я дж Расмуссен, Д.Т.; Саймонс, Э.Л. (1988). «Новые олигоценовые гиракоиды Египта». Журнал палеонтологии позвоночных . 8 (1): 67–83. Бибкод : 1988JVPal...8...67R . дои : 10.1080/02724634.1988.10011684 .
- ^ Jump up to: а б Мацумото, Х. (1922). «Мегалохиракс, Эндрюс и Титаногиракс, генер. ноябрь. Пересмотр родов гиракоидов из Фаюма, Египет». Труды Зоологического общества (1921): 839–850.
- ^ Jump up to: а б с д Расмуссен, Д.Т.; Саймонс, EL (1991). «Старейшие египетские гиракоиды (Mammalia: Pliohyracidae): новые виды Saghatherium и Thyrohyrax из Фаюма». Новогодний ежегодник геологии и палеонтологии . 182 (2): 187–209. дои : 10.1127/njgpa/182/1991/187 .
- ^ Хукер, Джей-Джей; Санчес-Вильягра, MR; Гоин, Ф.Дж.; Саймонс, Э.Л.; Аттия, Ю.; Зайферт, ER (2008). «Происхождение афро-арабских сумчатых дидельфиморфов» . Палеонтология . 51 (3): 635–648. Бибкод : 2008Palgy..51..635H . дои : 10.1111/j.1475-4983.2008.00779.x . S2CID 129506768 .
- ^ Jump up to: а б с д и ж Аль-Ашкар, Сан-Франциско; Зейферт, ER; де Врис, Д.; Эль-Сайед, С.; Антар, М.С.; Саллам, Его Величество (2021 г.). «Новые грызуны-фиокристомиины (Hystricognathi) из формации Джебель-Катрани, Фаюмская впадина, Египет» . ПерДж . 9 : e12074. дои : 10.7717/peerj.12074 . ПМЦ 8533026 . ПМИД 34721955 .
- ^ Jump up to: а б с д и ж г Саллам, HM; Зейферт, ER; Саймонс, Э.Л. (2012). «Базальная фиоморфа (Rodentia, Hystricognathi) из позднего эоцена Фаюмской впадины, Египет» . Швейцарский журнал палеонтологии . 131 (2): 283–301. дои : 10.1007/s13358-012-0039-6 . S2CID 84516606 .
- ^ Jump up to: а б Саллам, HM; Зайферт, ER (2016). «Новые фиоморфные грызуны из позднего эоцена Египта и влияние филогенетических методов, основанных на байесовских часах, на оценку базальных взаимоотношений гистрикогнатов и биохронологию» . ПерДж . 4 : е1717. дои : 10.7717/peerj.1717 . ПМЦ 4782727 . ПМИД 26966657 .
- ^ Саллам, HM; Зейферт, ER; Саймонс, Э.Л. (2011). «Краниодентальная морфология и систематика нового семейства гистрикогнатных грызунов (Gaudeamuridae) из позднего эоцена и раннего олигоцена Египта» . ПЛОС ОДИН . 6 (2): e16525. Бибкод : 2011PLoSO...616525S . дои : 10.1371/journal.pone.0016525 . ПМК 3043065 . ПМИД 21364934 .
- ^ Саллам, HM; Зайферт, ER (2020). «Ревизия олигоцена 'Paraphiomys' и происхождение кроны Thryonomyoidea (Rodentia: Hystricognathi: Phiomorpha) вблизи границы олигоцена и миоцена в Африке» . Зоологический журнал Линнеевского общества . 190 (1): 352–371. doi : 10.1093/zoolinnean/zlz148 .
- ^ Джагер, Джей-Джей; Мариво, Л.; Салем, М.; Билал, А.А.; Бенамми, М.; Чаймани, Ю.; Дюринер, П.; Марандат, Б.; Метаис, Э.; Шустер, М.; Валентин, X.; Брюне, М. (2010). «Новые комплексы грызунов с эоценового откоса Дур-ат-Талах (Сахара в центральной Ливии): систематические, биохронологические и палеобиогеографические значения» (PDF) . Зоологический журнал Линнеевского общества . 160 (1): 195–213. дои : 10.1111/j.1096-3642.2009.00600.x .
- ^ Jump up to: а б с Баун, ТМ; Саймонс, Э.Л. (1987). «Новые олигоценовые Ptolemaiidae (Mammalia: ?Pantolesta) из формации Джебель Катрани, Фаюмская впадина, Египет». Журнал палеонтологии позвоночных . 7 (3): 311–324. Бибкод : 1987JVPal...7..311B . дои : 10.1080/02724634.1987.10011662 .
- ^ Jump up to: а б с Саймонс, Эл.; Баун, ТМ (1995). «Птолемаида, новый отряд млекопитающих — с описанием черепа Ptolemaia grangeri» . Proc Natl Acad Sci США . 92 (8): 3269–73. Бибкод : 1995PNAS...92.3269S . дои : 10.1073/pnas.92.8.3269 . ПМК 42147 . ПМИД 11607526 .
- ^ Осборн, HF (1908). «Новые ископаемые млекопитающие фаюмского олигоцена, Египет» . Бюллетень Американского музея естественной истории . 24 : 265–272.
- ^ Jump up to: а б с д и ж г час я Зейферт, ER; Саймонс, Э.Л. (2013). «Последний из олигопитецидов? Карликовый вид с самого молодого уровня приматов формации Джебель-Катрани, северный Египет». Журнал эволюции человека . 64 (3): 211–215. дои : 10.1016/j.jhevol.2012.10.011 . ПМИД 23416038 .
- ^ Jump up to: а б с д и Саймонс, Эл.; Зейферт, ER; Чатрат, PS; Аттиа, Ю. (2001). «Самая ранняя запись антропоида-парапитецида из формации Джебель-Катрани, Северный Египет». Фолиа Приматологическая . 72 (6): 316–331. дои : 10.1159/000052748 . ПМИД 11964500 . S2CID 34259124 .
- ^ Jump up to: а б Саймонс, Э.Л. (1992). «Разнообразие ранней третичной антропоидной радиации в Африке» . Труды Национальной академии наук . 89 (22): 10743–10747. Бибкод : 1992PNAS...8910743S . дои : 10.1073/pnas.89.22.10743 . ПМЦ 50418 . ПМИД 1438271 .
- ^ Jump up to: а б Саймонс, Э.Л. (1989). «Описание двух родов и видов позднеэоценовых Anthropoidea из Египта» . Труды Национальной академии наук . 86 (24): 9956–9960. Бибкод : 1989PNAS...86.9956S . дои : 10.1073/pnas.86.24.9956 . ПМК 298621 . ПМИД 2513576 .
- ^ Саймонс, Эл.; Кей, РФ (1983). «Катрания, новый базальный человекообразный примат из Фаюма, олигоцен Египта». Природа . 304 (5927): 624–626. Бибкод : 1983Natur.304..624S . дои : 10.1038/304624a0 . S2CID 4232974 .
- ^ Домнинг, Д.П.; Джинджерич, PD; Залмут, И.С. (2010). «14: Сирения». Кайнозойские млекопитающие Африки . стр. 147–160.
- ^ Домнинг, ДП; Джинджерич, доктор медицинских наук; Саймонс, Эл.; Анкель-Саймонс, ФА (1994). «Новый дюгонгид раннего олигоцена (Mammalia, Sirenia) из провинции Фаюм, Египет» (PDF) . Материалы из Музея палеонтологии Мичиганского университета . 29 (4).
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть Эль-Садави, МЫ (2006). «Об ископаемой флоре района Джебель-Катрани, Фаюм, Египет» . Тэкхольмия . 26 (1): 131–140. дои : 10.21608/taec.2006.12289 .