Биолюминесцентные бактерии

Биолюминесцентные бактерии представляют собой светопродуктивные бактерии , которые преимущественно присутствуют в морской воде, морских отложениях, поверхности разлагающейся рыбы и в кишечнике морских животных. Несмотря на то, что бактериальная биолюминесценция, хотя и не так распространена, также обнаруживается у наземных и пресноводных бактерий. [ 1 ] Эти бактерии [ нужно разъяснения ] может быть свободная жизнь (например, Vibrio harveyi ) или в симбиозе с животными, такими как гавайский кальмар бобтейля ( Aliivibrio fischeri ) или наземные нематоды ( Photorhabdus luminescens ). Организма -хозяева обеспечивают эти бактерии [ нужно разъяснения ] Безопасный дом и достаточное питание. В обмене хозяева используют свет, произведенный бактериями для камуфляжа, добычи и/или спаривания. Биолюминесцентные бактерии развили симбиотические взаимоотношения с другими организмами, в которых оба участника выигрывают близко к одинаково. [ 2 ] Другая возможная причина, по которой бактерии используют реакцию люминесценции для чувствительности кворума , способность регулировать экспрессию генов в ответ на плотность бактериальных клеток. [ 3 ]
История
[ редактировать ]Записи биолюминесценции из -за бактерий существовали на протяжении тысячелетий. [ 4 ] Они появляются в фольклоре многих регионов, включая Скандинавию и индийский субконтинент. И Аристотель , и Чарльз Дарвин описали явление светящихся океанов. [ 4 ] С момента своего открытия менее 30 лет назад фермент люцифераза и ее регуляторный ген LUX привели к значительным достижениям в области молекулярной биологии, используя в качестве репортерного гена. [ 5 ] Люцифераза была впервые очищена от McElroy и Green в 1955 году. [ 6 ] Позже было обнаружено, что в люциферазе было две субъединицы, называемые субъединицами α и β. Гены, кодирующие эти ферменты, Luxa и Luxb, соответственно, были сначала изолированы в роскошном опероне Aliivibrio Fisheri [ 4 ] .
Цель био-люминесценции
[ редактировать ]с широкими биологическими целями Биологические цели био-люминесценции включают, но не ограничиваются притяжением товарищей, [ 7 ] защита от хищников и предупреждающие сигналы. В случае биолюминесцентных бактерий био-люминесценция в основном служит формой рассеивания. Было выдвинуто предположение, что кишечные бактерии (бактерии, которые выживают в кишках других организмов) - особенно те, которые распространены в глубине океана - используют био -люминесценцию в качестве эффективной формы распределения. [ 8 ] После того, как они пробились в пищеварительные тракты рыб и других морских организмов и выводятся в фекальных гранулах, биолюминесцентные бактерии способны использовать свои биолуминесцентные возможности для приманки в других организмах и быстрого проглатывания этих бактериальных фикальных пеллетов. [ 9 ] Таким образом, био-люминесценция бактерий гарантирует их выживание, настойчивость и рассеивание, поскольку они способны войти и обитать другие организмы.
Регуляция био-люминесценции
[ редактировать ]Регуляция био-люминесценции в бактериях достигается посредством регуляции окислительного фермента, называемого люциферазой . Важно, чтобы био-люминесцентные бактерии снижали скорость производства люциферазы, когда популяция в числе является скудной для сохранения энергии. Таким образом, бактериальная биолюминесценция регулируется с помощью химической связи, называемой чувствительностью кворума . [ 10 ] По сути, определенные сигнальные молекулы под названием аутоиндустровки [ 11 ] С специфическими бактериальными рецепторами активируются, когда плотность популяции бактерий достаточно высока. Активация этих рецепторов приводит к скоординированной индукции производства люциферазы, которая в конечном итоге дает видимую светильность . [ 12 ]
Биохимия био-люминесценции
[ редактировать ]
Химическая реакция, которая отвечает за био-люминесценцию, катализируется ферментом люциферазы . В присутствии кислорода люцифераза катализирует окисление органической молекулы, называемой люциферином . [ 13 ] Несмотря на то, что био-люминесценция в разнообразном диапазоне организмов, таких как бактерии, насекомые и динофлагелляты, функционируют в этом общем способе (используя люциферазу и люциферин), существуют различные типы систем люциферин-люциферазы. В частности, для бактериальной био-люминесценции биохимическая реакция включает окисление алифатического альдегида пониженным мононуклеотидом флавина . [ 14 ] Продукты этой реакции окисления включают окисленный мононуклеотид флавина, цепь жирных кислот и энергию в виде сине-зеленого видимого света. [ 15 ]
Реакция: fmnh 2 + o 2 + rcho → fmn + rcooh + h 2 o + light
Эволюция био-люминесценции
[ редактировать ]Из всех световых излучателей в океане био-люминесцентные бактерии являются наиболее распространенными и разнообразными. Тем не менее, распределение био-люминесцентных бактерий неравномерное, что предполагает эволюционные адаптации. Бактериальные виды в наземных родах, таких как Photorhabdus, являются био-люминесцентными. С другой стороны, морские роды с био-люминесцентными видами, такими как Vibrio и Shewanella onidensis, имеют разные тесно связанные виды, которые не являются излучателями света. [ 16 ] Тем не менее, все биолуминесцентные бактерии имеют общую последовательность генов: ферментативное окисление альдегида и снижение мононуклеотида флавина люциферазой , которые содержатся в опероне LUX. [ 17 ] Бактерии из различных экологических ниш содержат эту генную последовательность; Следовательно, идентичная последовательность генов, очевидно, предполагает, что био-люминесцентные бактерии являются результатом эволюционной адаптации.
Использовать в качестве лабораторного инструмента
[ редактировать ]После обнаружения оперона Lux использование биолюминесцентных бактерий в качестве лабораторного инструмента, как утверждается, произвело революцию в области микробиологии окружающей среды. [ 4 ] Применение биолюминесцентных бактерий включает биосенсоры для обнаружения загрязняющих веществ, измерения токсичности загрязняющих веществ [ 4 ] [ 18 ] [ 19 ] и мониторинг генетически инженерных бактерий, выпущенных в окружающую среду. [ 20 ] [ 21 ] [ 22 ] Биосенсоры, созданные путем размещения генной конструкции Lux под контролем индуцибельного промотора, могут использоваться для определения концентрации специфических загрязняющих веществ. [ 4 ] Биосенсоры также могут различать загрязнители, которые являются биодоступными, и теми, которые являются инертными и недоступными. [ 4 ] Например, Pseudomonas Fluorescens был генетически спроектирован, чтобы способствовать деградации салицилата и нафталина и используется в качестве биосенсора для оценки биодоступности салицилата и нафталина. [ 4 ] Биосенсоры также могут использоваться в качестве индикатора клеточной метаболической активности и для обнаружения присутствия патогенов. [ 4 ]
Эволюция
[ редактировать ]Светопроизводительная химия, стоящая за биолюминесценцией, зависит от линий биолюминесцентных организмов. [ 16 ] Основываясь на этом наблюдении, считается, что биолюминесценция развивалась независимо, по крайней мере, 40 раз. [ 16 ] В биолюминесцентных бактериях реклассификация членов группы видов вициро -фишера в качестве нового рода, Aliivibrio, привела к повышению интереса к эволюционному происхождению биолюминесценции [ 16 ] Полем Среди бактерий распределение биолюминесцентных видов является полифилетическим. Например, в то время как все виды в наземном роде Photorhabdus светятся, роды Aliivibrio, Photobacterium, Shewanella и Vibrio содержат как светящиеся, так и неястные виды. [ 16 ] Несмотря на биолюминесценцию у бактерий, не разделяющих общее происхождение, все они имеют общую последовательность генов. Появление высококонсервативного оперона Lux в бактериях из очень разных экологических ниш предполагает сильное избирательное преимущество, несмотря на высокие энергетические затраты на производство света. Считается, что восстановление ДНК является первоначальным селективным преимуществом для производства света у бактерий. [ 16 ] Следовательно, роскошный оперон, возможно, был потерян в бактериях, которые развивались более эффективные системы восстановления ДНК, но сохранялись в тех, где видимый свет стал селективным преимуществом. [ 16 ] [ 23 ] Считается, что эволюция чувствительности кворума дала дополнительное избирательное преимущество для производства света. Ощущение кворума позволяет бактериям сохранять энергию, гарантируя, что они не синтезируют светопроизводительные химические вещества, если не будет видна достаточная концентрация. [ 16 ]
Бактериальные группы, которые демонстрируют биолюминесценцию
[ редактировать ]Все виды бактерий, которые, как сообщалось, обладают биолюминесценцией, принадлежат семействам Vibrionaceae , Shewanellaceae или Enterobacteriaceae , все из которых отнесены к классовой гаммапротеобактерии . [ 24 ]
(Список от Dunlap and Henryk (2013), «светящиеся бактерии», прокариоты [ 24 ] )
Распределение
[ редактировать ]Биолюминесцентные бактерии наиболее распространены в морских средах во время весенних цветов, когда существуют высокие концентрации питательных веществ. Эти легкие организмы встречаются в основном в прибрежных водах возле оттока рек, таких как Северное Адриатическое море, Триесте, северо-западная часть Каспийского моря, побережье Африки и многих других. [ 25 ] Они известны как молочные моря . Биолюминесцентные бактерии также встречаются в пресной и наземной среде, но они менее распространены, чем в условиях морской воды. Они найдены во всем мире, как свободные, симбиотические или паразитные формы [ 1 ] и, возможно, как оппортунистические патогены . [ 24 ] Факторы, которые влияют на распределение биолюминесцентных бактерий, включают температуру, соленость, концентрацию питательных веществ, уровень рН и солнечное излучение. [ 26 ] Например, Aliivibrio Fischeri выгодно растет в средах, которые имеют температуру от 5 до 30 ° C, а pH - менее 6,8; Принимая во внимание, что фотобактерий фосфор процветает в условиях, которые имеют температуру от 5 до 25 ° С, а рН - менее 7,0. [ 27 ]
Генетическое разнообразие
[ редактировать ]Все биолюминесцентные бактерии имеют общую последовательность генов: оперон Lux , характеризующийся организацией гена LuxCdabe . [ 24 ] Коды люкс для люциферазы, в то время как LuxCDE коды для жирного акислого редуктазного комплекса, который отвечает за синтезирование альдегидов для биолюминесцентной реакции. Несмотря на эту общую генную организацию, вариации, такие как наличие других генов LUX, могут наблюдаться среди видов. Основываясь на сходстве в содержании и организации генов, оперон Lux может быть организован в следующие четыре различных типа: тип Aliivibrio / Shewanella , тип Photobacterium , тип вибрио / Candidatus photodesmus и тип Photorhabdus . Хотя эта организация следует по уровню классификации родов для членов Vibrionaceae ( Aliivibrio , Photobacterium и Vibrio ), его эволюционная история неизвестна. [ 24 ]
За исключением типа оперона Photorhabdus , все варианты оперона Lux , кодирующий флавин редуктазу содержат ген Luxg . [ 24 ] Большинство оперонов типа Aliivibrio / Shewanella содержат дополнительные регуляторные гены Luxi / Luxr , которые используются для аутоиндукции во время чувствительности кворума . [ 28 ] Тип оперона Photobacterum характеризуется наличием ребра , которые кодируют для рибофлавина, и образуют Lux-Rib оперон . Тип оперона Vibrio / Candidatus Photodesmus отличается как от типов оперонов Aliivibrio / Shewanella, так и от типов Photobacterium в том, что оперон не имеет регуляторных генов, непосредственно связанных с ним. [ 24 ]
Механизм
[ редактировать ]Все бактериальные люциферазы составляют приблизительно 80 кДа гетеродимеров, содержащих две субъединицы: α и β. Субъединица α отвечает за излучение света. [ 4 ] Гены Luxa и LuxB кодируют для субъединиц α и β, соответственно. В большинстве биолюминесцентных бактерий гены Luxa и LuxB окружены Luxc и Luxd и вниз по течению от Luxe . [ 4 ]
Биолюминесцентная реакция заключается в следующем:
Fmnh 2 + o 2 + r-so-> fmn + h 2 o + r-coh + свет (~ 495 нм)
Молекулярный кислород реагирует с FMNH 2 (снижение мононуклеотида флавина) и длинноцепочечным альдегидом для получения FMN (мононуклеотид флавина), водой и соответствующей жирной кислотой. Сине-зеленый световой излучение биолюминесценции, например, создаваемой фотобактерием фосфоры и Vibro Harveyi , является результатом этой реакции. [ 4 ] Поскольку выброс света включает в себя расходы на шесть молекул АТФ для каждого фотона, это энергетически дорогой процесс. По этой причине излучение света не экспрессируется в биолюминесцентных бактериях; Это выражается только тогда, когда физиологически необходимо.
Ощущение кворума
[ редактировать ]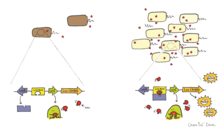
Биолюминесценция у бактерий может регулироваться с помощью явления, известного как аутоиндукция или чувствительность кворума . [ 4 ] Ощущение кворума является формой клеточной связи, которая изменяет экспрессию генов в ответ на плотность клеток. Аутоиндушер - это диффузированный феромон, продуцируемый конститутивно биолюминесцентными бактериями, и служит внеклеточной сигнальной молекулой. [ 4 ] Когда концентрация аутоиндуштора, секретируемой биолюминесцентными клетками в окружающей среде, достигает порога (выше 10 7 Клетки на мл), он индуцирует экспрессию люциферазы и других ферментов, участвующих в биолюминесценции. [ 4 ] Бактерии способны оценить их плотность, воспринимая уровень аутоиндуштора в окружающей среде и регулировать их биолюминесценцию, так что он выражается только тогда, когда существует достаточно высокая клеточная популяция. Достаточно высокая популяция клеток гарантирует, что биолюминесценция, продуцируемая клетками, будет видна в окружающей среде.
Хорошо известный пример чувства кворума - это то, что происходит между Аливибрио Фишери и его хозяином. Этот процесс регулируется Luxi и Luxr, кодируется Luxi и Luxr соответственно. Luxi является аутоинтазой, которая производит аутоиндустр (ИИ), в то время как LUXR функционирует как рецептор и транскрипционный фактор для оперона Lux. [ 4 ] Когда LUXR связывает ИИ, комплекс LUXR-AI активирует транскрипцию оперона LUX и вызывает экспрессию люциферазы . [ 28 ] Используя эту систему, A. fischeri показал, что биолюминесценция экспрессируется только тогда, когда бактерии связаны с хозяином и достигают достаточной плотности клеток. [ 29 ]
Другим примером чувствительности кворума биолюминесцентными бактериями является Vibrio Harveyi , который, как известно, свободно живет. В отличие от Aliivibrio Fischeri , V. Harveyi не обладают генами регулятора Luxi/Luxr и, следовательно, имеют другой механизм регуляции чувствительности кворума. Вместо этого они используют систему, известную как трехканальная система определения кворума. [ 30 ] Vibrio использует небольшие некодирующие РНК, называемые QRR RNA, для регулирования чувства кворума, используя их для контроля трансляции молекул энергии.
Роль
[ редактировать ]Использование биолюминесценции и ее биологического и экологического значения для животных, включая организмы -хозяева для симбиоза бактерий, широко изучалось. Биологическая роль и эволюционная история для специально биолюминесцентных бактерий по -прежнему остаются довольно таинственными и неясными. [ 4 ] [ 31 ] Тем не менее, существуют постоянно новые исследования для определения воздействия, которые бактериальная биолюминесценция может оказать на нашу постоянно меняющуюся среду и общество. Помимо многих научных и медицинских применений, ученые также недавно начали собираться вместе с художниками и дизайнерами, чтобы исследовать новые способы включения биолюминесцентных бактерий, а также биолюминесцентных растений, в городские источники света, чтобы уменьшить потребность в электричестве. [ 32 ] Они также начали использовать биолюминесцентные бактерии в качестве формы искусства и городского дизайна для чудеса и удовольствия от человеческого общества. [ 33 ] [ 34 ] [ 35 ]
Одним из объяснений роли бактериальной биолюминесценции является биохимический аспект. Несколько исследований показали биохимические роли пути люминесценции. Он может функционировать как альтернативный путь для электронного потока при низкой концентрации кислорода, что может быть выгодным, когда не доступен ферментируемый субстрат. [ 1 ] В этом процессе излучение света является побочным продуктом метаболизма.
Данные также свидетельствуют о том, что бактериальная люцифераза способствует устойчивости окислительного стресса. В лабораторной культуре Luxa и LUXB мутанты Vibrio Harveyi , которым не хватало активности люциферазы, показали нарушение роста при высоком окислительном стрессе по сравнению с диким типом. Мутанты LuxD , которые имели незатронутую люциферазу, но не смогли произвести люминесценцию, показали незначительную разницу. Это говорит о том, что люцифераза опосредует детоксикацию реактивного кислорода. [ 36 ]
Было также предполагается, что бактериальная биолюминесценция является источником внутреннего света при фотореактивации, процессе репарации ДНК, выполняемого фотолиазой. [ 37 ] Эксперименты показали, что не люминесцентные мутанты V. harveyi более чувствительны к УФ-облучению, что свидетельствует о существовании биолюминесцентной системы репарации ДНК. [ 23 ]
Другая гипотеза, называемая «гипотезой приманки», заключается в том, что бактериальная биолюминесценция привлекает хищников, которые помогут в их рассеивании. [ 37 ] Они либо напрямую принимают рыбу, либо косвенно принимаются зоопланктоном, который в конечном итоге будет потреблять более высокие трофические уровни. В конечном счете, это может позволить проникнуть в кишечник рыбы, богатую питательными веществами среда, где бактерии могут разделить, выделяться и продолжить свой цикл. Эксперименты с использованием Luminescent Photobacterium Leiognathi и не-люминесцентных мутантов показали, что люминесценция привлекает зоопланктон и рыбу, что подтверждает эту гипотезу. [ 37 ]
Симбиоз с другими организмами
[ редактировать ]Симбиотическая связь между гавайским боб хвостом кальмара эупримны и морской грамотрицательной бактерией Aliivibrio Fischeri была хорошо изучена. Два организма демонстрируют взаимные отношения, в которых биолюминесценция, продуцируемая A. fischeri, помогает привлечь добычу к хозяину кальмара, который обеспечивает богатые питательными веществами ткани и защищенную среду для A. fischeri . [ 38 ] Биолюминесценция, предоставленная A. fischeri, также помогает защите Squid E. Scolopes , обеспечивая камуфляж во время ночного питания. [ 39 ] После бактериальной колонизации специализированные органы кальмара претерпевают изменения в развитии, и становятся установленными отношениями. Кальмар выпускает 90% бактериальной популяции каждое утро, потому что ему больше не нужно производить биолюминесценцию при дневном свете. [ 4 ] Это изгнание приносит пользу бактериям, помогая в их распространении. Одно исключение одним кальмаром с боб хвоста производит достаточное количество бактериальных симбионтов, чтобы заполнить 10 000 м. 3 морской воды в концентрации, которая сопоставимо с тем, что находится в прибрежных водах. [ 39 ] Таким образом, по крайней мере, в некоторых местах обитания симбиотическая связь между А. Фишери и Э. Сколопес играет ключевую роль в определении численности и распределения Э. Сколопа . наблюдается более высокое содержание A. fischeri В окрестностях популяции E. scolopes , и это изобилие заметно уменьшается с увеличением расстояния от среды обитания хозяина. [ 39 ]
Биолюминесцентные виды фотобактерии также участвуют в взаимовыгодных ассоциациях с рыбой и кальмарами. [ 40 ] Плотные популяции P. kishitanii, P. leiogathi и могут жить в легких органах морской рыбы и кальмара и P. mandapamensis снабжены питательными веществами и кислородом для размножения [ 40 ] В обмен на предоставление биолюминесценции их хозяевам, что может помочь в специфической для пола передачи сигналов, предотвращения хищников, поиска или привлечения добычи и обучения. [ Цитация необходима ] Интересно, что Мейер-Рохов сообщил в 1976 году, что если рыба не может получить пищу и голодает, свет его биолюминесцентного симбионта становится все более тусклостью, пока световой излучение вообще не остановится. [ 41 ]
| В приведенной ниже таблице изображения справа указывают в синем месте местоположение светового органа разных семей симбиотически светящейся рыбы и кальмара. [ 42 ] E указывает на внешнее изгнание биолюминесцентных бактерий непосредственно в морскую воду. Я указывает внутреннее изгнание биолюминесцентных бактерий в пищеварительном тракте. (E) или (i) указывать предполагаемую локализацию изгнания. [ 43 ] |

|
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в Нилсон, KH; Гастингс, JW (1979-12-01). «Бактериальная биолюминесценция: ее контрольная и экологическая значимость» . Микробиологические обзоры . 43 (4): 496–518. doi : 10.1128/mmbr.43.4.496-518.1979 . ISSN 0146-0749 . PMC 281490 . PMID 396467 .
- ^ Макфолл-Нгай, Маргарет; Хит-Хекман, Элизабет А.К.; Джилетт, Амани А.; Пейер, Сюзанна М.; Харви, Элизабет А. (2012). «Секретные языки коэворированных симбиозов: понимание эупримнского сколопса - Вибрио Фишчери Симбиоз» . Семинары по иммунологии . 24 (1): 3–8. doi : 10.1016/j.smim.2011.11.006 . PMC 3288948 . PMID 22154556 .
- ^ Уотерс, Кристофер М.; Басслер, Бонни Л. (2005-10-07). «Ощущение кворума: коммуникация клеток к бактериям». Ежегодный обзор биологии клеток и развития . 21 (1): 319–346. doi : 10.1146/annurev.cellbio.21.012704.131001 . PMID 16212498 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий Nunes-Halldorson, Вания да Силва; Duran, Norma Leticia (2003-06-01). «Биолюминесцентные бактерии: роскошные гены экологического биозонсора» . Бразильский журнал микробиологии . 34 (2): 91–96. Doi : 10.1590/s1517-83822003000200001 . ISSN 1517-8382 .
- ^ «Репортеры люциферазы» . www.thermofisher.com . Получено 2017-03-24 .
- ^ МакЭлрой, WD; Зеленый (1954). «Ферментативные свойства бактериальной люциферазы». Архивы биохимии и биофизики . 56 (1): 240–255. doi : 10.1016/0003-9861 (55) 90353-2 . PMID 14377570 .
- ^ Херринг, Питер Дж. (2007-08-01). «Обзор. Секс с огнями? Обзор биолюминесцентного сексуального диморфизма в море» . Журнал Морской биологической ассоциации Соединенного Королевства . 87 (4): 829–842. Bibcode : 2007jmbuk..87..829h . doi : 10.1017/s0025315407056433 . ISSN 1469-7769 . S2CID 86616254 .
- ^ Руби, например; Морин, JG (1979-09-01). «Светящиеся кишечные бактерии морских рыб: изучение их распределения, плотности и дисперсии» . Прикладная и экологическая микробиология . 38 (3): 406–411. Bibcode : 1979apenm..38..406r . doi : 10.1128/aem.38.3.406-411.1979 . ISSN 0099-2240 . PMC 243508 . PMID 16345429 .
- ^ Руби, например; Морин, JG (1979-09-01). «Светящиеся кишечные бактерии морских рыб: изучение их распределения, плотности и дисперсии» . Прикладная и экологическая микробиология . 38 (3): 406–411. Bibcode : 1979apenm..38..406r . doi : 10.1128/aem.38.3.406-411.1979 . ISSN 0099-2240 . PMC 243508 . PMID 16345429 .
- ^ Уилсон, Тереза; Гастингс, Дж. Вудленд (2003-11-28). «Биолюминесценция». Ежегодный обзор биологии клеток и развития . 14 (1): 197–230. doi : 10.1146/annurev.cellbio.14.1.197 . PMID 9891783 .
- ^ Эберхард, А.; Burlingame, AL; Эберхард, C.; Кеньон, Гл; Нилсон, KH; Оппенгеймер, Нью-Джерси (1981-04-01). «Структурная идентификация аутоиндуировщика фотобактерии Фишчери люциферазы». Биохимия . 20 (9): 2444–2449. doi : 10.1021/bi00512a013 . ISSN 0006-2960 . PMID 7236614 .
- ^ Перес-Веласкес, Джудит; Gölgeli, Meltem; García-Contreras, Родольфо (2016-08-25). «Математическое моделирование бактериального определения кворума: обзор» (PDF) . Бюллетень математической биологии . 78 (8): 1585–1639. doi : 10.1007/s11538-016-0160-6 . HDL : 11693/36891 . ISSN 0092-8240 . PMID 27561265 . S2CID 3940300 .
- ^ Каскова, Зинаида М.; Царкова, Александра С.; Yampolsky, Ilia V. (2016-10-24). «1001 свет: люциферины, люциферазы, их механизмы действия и применения в химическом анализе, биологии и медицине». Химический Соц Преподобный 45 (21): 6048–6077. doi : 10.1039/c6cs00296j . ISSN 1460-4744 . PMID 27711774 .
- ^ Уайлс, Сиуси ; Фергюсон, Кэтрин; Стефаниду, Марта; Янг, Дуглас Б.; Робертсон, Брайан Д. (2005-07-01). «Альтернативная люцифераза для мониторинга бактериальных клеток в неблагоприятных условиях» . Прикладная и экологическая микробиология . 71 (7): 3427–3432. Bibcode : 2005apenm..71.3427W . doi : 10.1128/aem.71.7.3427-3432.2005 . ISSN 0099-2240 . PMC 1169068 . PMID 16000745 .
- ^ Фишер, Эндрю Дж.; Томпсон, Томас Б.; Тоден, Джеймс Б.; Болдуин, Томас О.; Реймент, Иван (1996-09-06). «Кристаллическая структура разрешения 1,5 Å бактериальной люциферазы в условиях низкой соли» . Журнал биологической химии . 271 (36): 21956–21968. doi : 10.1074/jbc.271.36.21956 . ISSN 0021-9258 . PMID 8703001 .
- ^ Jump up to: а беременный в дюймовый и фон глин час Widder, EA (2010-05-07). «Биолюминесценция в океане: происхождение биологического, химического и экологического разнообразия». Наука . 328 (5979): 704–708. Bibcode : 2010sci ... 328..704W . doi : 10.1126/science.1174269 . ISSN 0036-8075 . PMID 20448176 . S2CID 2375135 .
- ^ О'Кейн, Деннис Дж.; Prasher, Douglas C. (1992-02-01). «Эволюционное происхождение бактериальной биолюминесценции» . Молекулярная микробиология . 6 (4): 443–449. doi : 10.1111/j.1365-2958.1992.tb01488.x . ISSN 1365-2958 . PMID 1560772 . S2CID 42286135 .
- ^ Ptitsyn, LR; Horneck, G.; Комова, О.; Kozubek, S.; Красавин, EA; Bonev, M.; Реттберг, П. (1997-11-01). «Биосенсор для скрининга генотоксина окружающей среды, основанный на анализе SOS Lux в рекомбинантных клетках Escherichia coli» . Прикладная и экологическая микробиология . 63 (11): 4377–4384. Bibcode : 1997apenm..63.4377p . doi : 10.1128/aem.63.11.4377-4384.1997 . ISSN 0099-2240 . PMC 168758 . PMID 9361425 .
- ^ Widder, Edith A.; Фолс, Бет (март 2014 г.). «Обзор биолюминесценции для инженеров и ученых в биофотонике». IEEE Журнал отдельных тем в квантовой электронике . 20 (2): 232–241. Bibcode : 2014ijstq..20..232W . doi : 10.1109/jstqe.2013.2284434 . ISSN 1077-260X . S2CID 33866877 .
- ^ Махафи, WF; Бауске, Эм; Ван Вуурде, JW; Ван дер Вольф, JM; Ван Ден Бринк, М.; Kloepper, JW (1997-04-01). «Сравнительный анализ устойчивости к антибиотикам, иммунофлуоресцентного окрашивания колонии и трансгенного маркера (биолюминесценция) для мониторинга окружающей среды Rhizobacterium» . Прикладная и экологическая микробиология . 63 (4): 1617–1622. Bibcode : 1997apenm..63.1617m . doi : 10.1128/aem.63.4.1617-1622.1997 . ISSN 0099-2240 . PMC 168454 . PMID 9097457 .
- ^ Шоу, JJ; Dane, F.; Гейгер, Д.; Kloepper, JW (1992-01-01). «Использование биолюминесценции для обнаружения генетически инженерных микроорганизмов, выпущенных в окружающую среду» . Прикладная и экологическая микробиология . 58 (1): 267–273. Bibcode : 1992apenm..58..267s . doi : 10.1128/aem.58.1.267-273.1992 . ISSN 0099-2240 . PMC 195202 . PMID 1311542 .
- ^ Флемминг, Калифорния; Lee, H.; Trevors, JT (1994-09-01). «Биолюминесцентный наиболее широкий способ перечисления роскошного маркированного Pseudomonas aeruginosa ug2lr в почве» . Прикладная и экологическая микробиология . 60 (9): 3458–3461. Bibcode : 1994apenm..60.3458f . doi : 10.1128/aem.60.9.3458-3461.1994 . ISSN 0099-2240 . PMC 201832 . PMID 16349396 .
- ^ Jump up to: а беременный Czyz, A.; Wrobel, B.; Wegrzyn, G. (2000-02-01). «Биолюминесценция Vibrio Harveyi играет роль в стимуляции репарации ДНК» . Микробиология . 146 (2): 283–288. doi : 10.1099/00221287-146-2-283 . ISSN 1350-0872 . PMID 10708366 .
- ^ Jump up to: а беременный в дюймовый и фон глин Делонг, Эдвард Ф.; Stackebrandt, Erko; Лори, Стивен; Томпсон, Фабиано (2013). Прокариоты - Springer . doi : 10.1007/978-3-642-31331-8 . ISBN 978-3-642-31330-1 Полем S2CID 5215416 .
- ^ Стейплс, Роберт Ф.; States., United (1966). Детали - распределение и характеристики поверхностной биолюминесценции в океанах / Роберт Ф. Стейпле. - Библиотека наследия биоразнообразия . doi : 10.5962/bhl.title.46935 .
- ^ Soto, W.; Gutierrez, J.; Remmenga, MD; Nishiguchi, MK (2009-01-01). «Соленость и температура на физиологические реакции Vibrio fischeri из разнообразных экологических ниш» . Микробная экология . 57 (1): 140–150. Bibcode : 2009micec..57..140s . doi : 10.1007/s00248-008-9412-9 . ISSN 0095-3628 . PMC 2703662 . PMID 18587609 .
- ^ Уотерс, Пол; Ллойд, Дэвид (1985). «Соль, рН и температурная зависимости роста и биолюминесценции трех видов светящихся бактерий, проанализированных на градиентных пластинах» . Журнал общей микробиологии . 131 (11): 65–9. doi : 10.1099/00221287-131-11-2865 .
- ^ Jump up to: а беременный Bassler, BL (1999-12-01). «Как бактерии разговаривают друг с другом: регуляция экспрессии генов путем чувствительности кворума». Текущее мнение о микробиологии . 2 (6): 582–587. doi : 10.1016/s1369-5274 (99) 00025-9 . ISSN 1369-5274 . PMID 10607620 .
- ^ Fuqua, WC; Winans, SC; Гринберг, EP (1994-01-01). «Ощущение кворума у бактерий: семейство регуляторов транскрипции, реагирующих на плотность клеток» . Журнал бактериологии . 176 (2): 269–275. doi : 10.1128/jb.176.2.269-275.1994 . ISSN 0021-9193 . PMC 205046 . PMID 8288518 .
- ^ Декабря, Том; Продолжить, Нико; Соргес, Патрик; Штаммы, Вилли; Боссье, Питер (2008-01-01). «Ощущение кворума и гашение кворума в Vibbrio Harmorolyi: арендованный этаж in vivoy» . Журнал ISME . 2 (1): 19–26. Bibcode : 200888sme… 2… 19d . doi : 10 1038/ism.2007.92 . ISSN 1751-7362 . PMID 18180744 .
- ^ Ванньер, Томас; Хингам, Паскаль; Туррел, Флория; Танет, Лиза; Лескот, Магали; Timsit, Youi (2020-06-01). «Разнообразие и эволюция генов бактериальной биолюминесценции в глобальном океане» . NAR Геномика и биоинформатика . 2 (2): LQAA018. doi : 10.1093/nargab/lqaa018 . PMC 7671414 . PMID 33575578 .
- ^ Арддавани, Олимпия; Zerefos, Stelios; Дулос, Ламброс Т. (2020-01-01). «Переиздание внешнего освещения в рамках городского ландшафта: роль трансгенных биолюминесцентных растений в средиземноморских городских и пригородных освещениях» . Журнал чистого производства . 242 : 118477. Bibcode : 2020JCPRO.24218477A . doi : 10.1016/j.jclepro.2019.118477 . ISSN 0959-6526 .
- ^ Лац, Майкл I. (2019). «Создание бесконечного куба, выставка биолюминесценции» . Лимнология и океанографический бюллетень . 28 (4): 130–134. Bibcode : 2019limob..28..130L . doi : 10.1002/lob.10345 . ISSN 1539-6088 .
- ^ Дибас, Шерил Лин (2019). «Новая волновая наука и искусство встречаются в море». Океанография . 32 (1): 10–12. ISSN 1042-8275 . JSTOR 26604937 .
- ^ Фот, Маркус; Caldwell, Glenda Amayo (2018). «Более чем мужская медийная архитектура» (PDF) . Материалы 4 -й конференции по биеннале в области медиа -архитектуры . С. 66–75. doi : 10.1145/32843889.3284495 . ISBN 978-1-4503-6478-2 Полем S2CID 69160272 .
- ^ Szpilewska, Hanna; Чиз, Агата; Wegrzyn, Grzegorz (2003-11-01). «Экспериментальные данные физиологической роли бактериальной люциферазы в защите клеток от окислительного стресса». Текущая микробиология . 47 (5): 379–382. doi : 10.1007/s00284-002-4024-y . ISSN 0343-8651 . PMID 14669913 . S2CID 20132449 .
- ^ Jump up to: а беременный в Зарубин, Маргарита; Белкин, Шимшон; Ионеску, Майкл; Genin, Amatzia (2012-01-17). «Бактериальная биолюминесценция как приманка для морского зоопланктона и рыбы» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (3): 853–857. Bibcode : 2012pnas..109..853z . doi : 10.1073/pnas.1116683109 . ISSN 1091-6490 . PMC 3271926 . PMID 22203999 .
- ^ Нотон, Линн М.; Мандель, Марк Дж. (2012-03-01). «Колонизация эупримны Scolopes Squid от Vibrio fischeri » . Журнал визуализированных экспериментов (61): E3758. doi : 10.3791/3758 . ISSN 1940-087X . PMC 3399469 . PMID 22414870 .
- ^ Jump up to: а беременный в Руби, например; Ли, К.Х. (1998-03-01). «Вибрио-фишер-эупримна сколопскую ассоциацию органов: современные экологические парадигмы» . Прикладная и экологическая микробиология . 64 (3): 805–812. Bibcode : 1998apenm..64..805r . doi : 10.1128/aem.64.3.805-812.1998 . ISSN 0099-2240 . PMC 106330 . PMID 16349524 .
- ^ Jump up to: а беременный Urbanczyk, Генрик; Аст, Дженнифер С.; Dunlap, Paul V. (2011-03-01). «Филогения, геномика и симбиоз фотобактерии» . Обзоры микробиологии FEMS . 35 (2): 324–342. doi : 10.1111/j.1574-6976.2010.00250.x . ISSN 0168-6445 . PMID 20883503 .
- ^ Мейер-Рохов, Виктор Бенно (1976). «Потеря биолюминесценции в аномалопах Катоптрон из -за голода». Экспериментация . 32 (9): 1175–1176. doi : 10.1007/bf01927610 .
- ^ Nealson, KH и Hastings, JW (1979) «Бактериальная биолюминесценция: ее контрольная и экологическая значимость», Microbiol. Rev. , 43 : 496–518. Doi : 10.1128/mmbr.43.4.496-518.1979 .
- ^ Танет, Лиза; Мартини, Северин; Казалот, Лори; Тамбурини, Кристиан (2020). «Обзоры и синтезы: бактериальная биолюминесценция - экология и воздействие в биологическом углеродном насосе» . Биогеонов . 17 (14): 3757–3778. Бибкод : 2020bgeo ... 17.3757t . doi : 10.5194/bg-17-3757-2020 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .