Главный комплекс гистосовместимости и половой отбор

Гены главного комплекса гистосовместимости (MHC) кодируют белки клеточной поверхности, которые облегчают иммунный ответ организма на патогены, а также его способность избегать нападения на собственные клетки. Эти гены поддерживали необычайно высокий уровень аллельного разнообразия на протяжении веков и в разных популяциях. Это означает, что для каждого гена MHC в популяции постоянно существует множество аллелей (или вариантов гена), и многие люди гетерозиготны по MHC локусам (это означает, что они обладают двумя разными аллелями для данного локуса гена).
Огромный источник генетических вариаций , влияющих на приспособленность организма, проистекает из коэволюционной гонки вооружений между хозяевами и паразитами. Есть две гипотезы, объясняющие большое разнообразие MHC, которые не являются взаимоисключающими. Во-первых, существует отбор людей, обладающих разнообразным набором аллелей MHC, что может произойти, если гетерозиготы MHC более устойчивы к патогенам, чем гомозиготы - это называется преимуществом гетерозигот . Во-вторых, существует отбор, который проходит частотно-зависимый цикл — это называется гипотезой Красной Королевы .
Есть свидетельства того, что многие позвоночные, включая человека, выбирают себе партнеров на основе сигналов «совместимости» между их аллелями MHC, отдавая предпочтение партнерам с аллелями, отличными от их собственных, что приводит к образованию пар, которые имеют тенденцию производить более гетерозиготное потомство. Существует несколько предложенных гипотез, которые касаются того, как брачные предпочтения, связанные с MHC, могут быть адаптивными и как в MHC сохраняется необычно большое количество аллельного разнообразия. [ 1 ] [ 2 ]
Гипотезы
[ редактировать ]Согласно первой гипотезе, если особи, гетерозиготные по MHC, более устойчивы к паразитам, , чем гомозиготные то самкам выгодно выбирать партнеров с генами MHC, отличными от их собственных, и это приведет к появлению MHC-гетерозиготного потомства - это известное как дисассортативное спаривание . Гипотеза утверждает, что люди с гетерозиготным MHC будут способны распознавать более широкий спектр патогенов и, следовательно, вызывать специфический иммунный ответ против большего числа патогенов, имея таким образом преимущество в иммунитете . К сожалению, гипотеза о преимуществе MHC-гетерозигот не была должным образом проверена. [ 2 ] Гены иммунитета, не относящиеся к MHC, у разных видов демонстрируют недостаток гетерозигот или не имеют преимуществ. [ 3 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ] [ 8 ] У мышей повышенная гетерозиготность MHC снижает приспособленность, что ставит под сомнение эту гипотезу. MHC-гетерозиготные самки имели значительно более низкую приспособленность по сравнению с гомозиготными. [ 9 ] Этот вывод был повторен в другом исследовании на мышах и снова на рыбах. [ 10 ] [ 11 ] В некоторых случаях избыточная гетерозиготность может привести к снижению приспособленности. [ 12 ]
Гипотеза оптимальности утверждает, что слишком большая вариабельность MHC может привести к неспособности Т-клеток отличить себя от чужих и тем самым увеличить риск аутоиммунных заболеваний. Это обеспечит большую приспособленность людям, не имеющим большого разнообразия MHC. [ 6 ] [ 13 ] Аутоиммунные заболевания связаны с локусами MHC. У людей люди с большим разнообразием MHC имеют больший риск аутоиммунных заболеваний. Разнообразие MHC может быть низким, «поскольку чужеродные пептиды должны выделяться на собственном фоне». На индивидуальном уровне разнообразие MHC имеет тенденцию быть низким. У многих видов существует промежуточная гетерозиготность по MHC. Общие данные подтверждают, что промежуточная гетерозиготность MHC является наилучшей. [ 14 ]
Гипотеза Красной Королевы утверждает, что разнообразие MHC поддерживается паразитами. MHC индивидуумов Если аллели оказывают различную устойчивость к определенному паразиту, то аллель с наибольшей устойчивостью отдается предпочтение, отбирается и, следовательно, распространяется по всей популяции. Рекомбинация и мутация вызывают появление новых вариантов среди потомства, что может способствовать быстрой реакции на быстро развивающиеся паразиты или патогены с гораздо более коротким временем генерации. Однако, если этот конкретный аллель становится общим, давление отбора на паразитов, чтобы избежать распознавания по этому общему аллелю, возрастает. Полезная характеристика, которая позволяет паразиту избежать распознавания, распространяется и вызывает отбор против того, что раньше было устойчивым аллелем. Это позволяет паразиту избежать этого цикла частотно-зависимого отбора, и такой цикл в конечном итоге приводит к коэволюционной гонке вооружений, которая может поддерживать поддержание разнообразия MHC. Эта гипотеза имеет эмпирическую поддержку. [ 15 ] [ 2 ] [ 16 ]

Гипотеза избегания инбридинга имеет меньшее отношение к отношениям хозяин-паразит, чем гипотеза преимущества гетерозигот или гипотеза Красной Королевы. Чрезвычайное разнообразие MHC приведет к тому, что люди, имеющие общие аллели MHC, с большей вероятностью будут связаны между собой. В результате одной из функций MHC-дисассортативного спаривания будет предотвращение спаривания с членами семьи и любых вредных генетических последствий, которые могут возникнуть в результате. Гипотеза утверждает, что инбридинг увеличивает общую гомозиготность - не только локально в MHC, поэтому увеличение генетической гомозиготности может сопровождаться не только проявлением рецессивных заболеваний и мутаций, но также потерей любого потенциального преимущества гетерозигот. . [ 17 ] [ 2 ] Животные лишь в редких случаях избегают инбридинга. [ 18 ] Гипотеза избегания инбридинга была «исключена как объяснение наблюдаемой закономерности MHC-зависимых предпочтений партнера», поскольку родство не связано с выбором партнера. [ 19 ]
В ходе поиска потенциальных партнеров самкам будет полезно иметь возможность различать «плохие» гены, чтобы повысить здоровье и жизнеспособность своего потомства. Если выбор самки партнера происходит на основе «хороших» генов, то подразумевается, что среди самцов существуют генетические вариации. Более того, можно было бы предположить, что указанное различие в генах также будет влиять на разницу в приспособленности, которую потенциально можно было бы выбрать или отобрать для нее.
Как правило, крайний полиморфизм генов MHC выбирается в результате гонки вооружений между хозяином-паразитом ( гипотеза Красной Королевы ); однако дисассортативный выбор партнера может поддерживать генетическое разнообразие у некоторых видов. В зависимости от того, как паразиты изменяют отбор аллелей MHC, MHC-зависимый выбор партнера может повысить приспособленность потомства за счет повышения его иммунитета, как упоминалось ранее. Если это так, либо через гипотезу преимущества гетерозигот, либо через гипотезу Красной Королевы, то отбор также благоприятствует практикам спаривания, которые зависят от MHC.

Таким образом, выбор партнера - относительно MHC - вероятно, развился так, что самки выбирают самцов либо на основе разнообразных генов (гипотеза преимущества гетерозигот и предотвращения инбридинга), либо на «хороших» генах. Тот факт, что самки выбирают, является естественным выбором, поскольку для самок было бы выгодно иметь возможность выбирать самца, который приносил бы либо косвенную, либо прямую выгоду. В результате женского выбора половой отбор навязывается самцам. Об этом свидетельствует генетическая «реклама» — примером может служить существование преувеличенных черт, таких как сложные хвостовые перья самцов павлинов. Однако у людей выбор партнера осуществляют оба пола.
Связь между обонянием и MHC
[ редактировать ]Известно, что половой отбор на основе MHC задействует обонятельные механизмы у таких таксонов позвоночных, как рыбы, мыши, люди, приматы, птицы и рептилии. [ 1 ] На самом простом уровне люди уже давно знакомы с чувством обоняния, поскольку оно используется для определения приятности или неприятности ресурсов, еды и т. д. На более глубоком уровне было предсказано, что обоняние служит для личной идентификации людей на основе на генах MHC. [ 20 ]
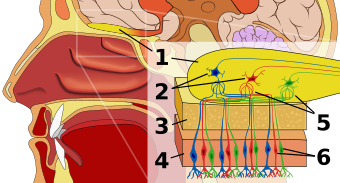
Хемосенсация, одно из самых примитивных чувств, превратилась в специализированную сенсорную систему. Люди могут не только обнаруживать, но также оценивать и реагировать на обонятельные сигналы окружающей среды (химические), особенно те, которые используются для вызова поведенческих и сексуальных реакций у других людей, также известных как феромоны . Феромоны сообщают о видовой принадлежности, поле и, что, возможно, самое важное, о генетической идентичности человека. Гены MHC обеспечивают основу, на которой развивается набор уникального обонятельного кодирования. [ 20 ]
Хотя точно неизвестно, как распознаются запахи, специфичные для MHC, в настоящее время считается, что белки, связанные с пептид-связывающей бороздкой MHC, могут производить запах. Каждый белок MHC связывается с определенной пептидной последовательностью, образуя набор уникально связанных комплексов пептид-MHC для каждого человека. Во время клеточного обмена комплекс MHC-пептид выделяется с поверхности клетки, а его фрагменты распределяются в жидкостях организма, таких как сыворотка крови, слюна и моча. Ученые полагают, что комменсальная микрофлора, микроорганизмы, выстилающие поверхности эпителия, открытые для внешней среды, такие как желудочно-кишечный тракт и влагалище, еще больше разлагают эти фрагменты, которые в результате этого процесса становятся летучими. Недавно было показано, что рецепторы вомероназального органа мышей активируются пептидами, имеющими характеристики, сходные с белками MHC; Надеемся, что дальнейшие исследования вскоре смогут прояснить точную трансформацию между генотипом MHC и обонятельным механизмом. [ 1 ] [ 20 ] [ 21 ]
Эмпирические данные
[ редактировать ]У людей
[ редактировать ]Сходство MHC у людей изучалось по трем основным направлениям: запах, привлекательность лица и фактический выбор партнера. [ 22 ] Исследования запаха обнаруживают предпочтения несходства MHC, но различаются в деталях, в то время как привлекательность лица благоприятствует сходству MHC, и фактические исследования спаривания различаются. [ 22 ]
Конкретные исследования
[ редактировать ]Некоторые исследования показывают, что предпочтения запахов и выбор партнера, связанные с MHC, демонстрируются людьми. Однако роль MHC в выборе партнера у человека относительно противоречива. Одно исследование, проведенное Ober et al. исследовали типы HLA у 400 пар в сообществе гуттеритов и обнаружили значительно меньше совпадений HLA между мужьями и женами, чем ожидалось, учитывая социальную структуру их сообщества. [ 23 ] С другой стороны, в том же исследовании 200 пар из южноамериканских племен не было обнаружено никаких доказательств выбора партнера на основе MHC. [ 23 ]
Другие исследования подходили к выбору партнера на основе предпочтений запаха. В одном исследовании, проведенном Ведекиндом и др., женщин попросили понюхать мужские подмышечные запахи, собранные на футболках, которые носят разные мужчины. Женщины, у которых была овуляция, оценили запахи мужчин, отличающихся по MHC, как более приятные, чем запахи мужчин, похожих по MHC. Более того, запахи мужчин, отличающихся по MHC, часто напоминали женщинам о нынешних или бывших партнерах, что позволяет предположить, что запах – особенно запах несходства по MHC – играет роль в выборе партнера. [ 24 ]
В другом исследовании, проведенном Ведекиндом и др., 121 женщину и мужчину попросили оценить приятность запаха потных футболок. Понюхав рубашки, выяснилось, что мужчины и женщины, которым напоминали своего партнера или бывшего партнера, имели значительно меньше общих аллелей MHC с владельцем, чем можно было бы случайно ожидать. Если выбор рубашек не был случайным, а на самом деле был выбран по разным MHC аллелям, это предполагает, что генетический состав MHC действительно влияет на выбор партнера. Более того, когда степень сходства между носителем и нюхателем была статистически учтена, значительного влияния MHC на предпочтение запаха больше не наблюдалось. Результаты показывают, что сходство или различие MHC определенно играет роль в выборе партнера. В частности, для этого выбирают MHC-дисассортативный выбор партнера и менее похожие комбинации MHC. [ 25 ] Одним из интересных аспектов эксперимента Ведекинда было то, что в отличие от женщин, обычно ездящих на велосипеде, женщины, принимающие оральные контрацептивы, предпочитали запахи MHC-подобных мужчин. Это предполагает, что таблетка может помешать адаптивному предпочтению несходства. [ 24 ] [ 25 ]
У приматов
[ редактировать ]Имеются доказательства выбора партнера, связанного с MHC, у других приматов. У серого мышиного лемура Microcebus murinus посткопулятивный выбор партнера связан с генетической конституцией. Отцы более MHC-отличны от матери, чем случайно протестированные мужчины. Отцы имеют больше различий в разнообразии аминокислот и микросателлитов, чем случайно протестированные мужчины. Предполагается, что это вызвано загадочным выбором женщины. [ 26 ]
У других животных
[ редактировать ]У мышей как самцы, так и самки выбирают MHC-разных партнеров. Мыши развивают способность идентифицировать членов семьи на раннем этапе роста и, как известно, избегают инбридинга с родственниками, что подтверждает гипотезу выбора партнера, опосредованного MHC, для предотвращения инбридинга. [ 2 ]
Рыбы — еще одна группа позвоночных, демонстрирующая выбор партнера, связанный с MHC. Ученые протестировали атлантического лосося Salmo salar , наблюдая за воздействием MHC на естественно нерестящегося лосося, обитающего в реке, по сравнению с искусственными скрещиваниями, которые проводились в инкубаториях. Логично, что искусственные скрещивания будут лишены преимуществ выбора партнера, которые естественным образом будут доступны. Результаты показали, что потомство искусственно выращенного лосося было более заражено паразитами: почти в четыре раза больше, чем потомство, выведенное естественным путем. Кроме того, дикое потомство было более MHC-гетерозиготным, чем искусственно выведенное потомство. Эти результаты подтверждают гипотезу гетерозиготного преимущества о половом отборе для выбора партнера, несходного по MHC. [ 27 ] На примере другой рыбы, трехиглой колюшки, было показано, что самки желают разнообразия MHC в своем потомстве, что влияет на их выбор партнера. [ 28 ]
Самки саваннских воробьев Passerculus Sandensis для спаривания выбирали самцов, не похожих по MHC. Самки с большей вероятностью будут вступать во внепарные отношения, если они находятся в паре с партнерами, похожими на MHC, и доступно больше непохожих партнеров. Точно так же разнообразие MHC у домашних воробьев Passer Domesticus предполагает, что происходит дисассортативный MHC-выбор партнера. [ 2 ]
Было показано, что MHC-опосредованный выбор партнера существует у шведских песчаных ящериц Lacerta agilis . Самки предпочитали ассоциироваться с образцами запахов, полученными от самцов, более отдаленно родственных по локусам MHC I. [ 29 ]
Несмотря на то, что многие виды социально моногамны, самки могут принимать или активно искать спаривание вне отношений; [ 30 ] Отцовство вне пары - это модель спаривания, которая, как известно, связана с выбором партнера, связанным с MHC. Птицы — одна из наиболее часто изучаемых групп животных, демонстрирующих такое сексуальное поведение. У алого чечевица Carpocus erythrinus самки гораздо реже осуществляли внепарное отцовство, когда их самки были MHC-гетерозиготными. [ 31 ] У сейшельской славки Acrocephalus sechellensis не было выявлено различий MHC между социальными партнерами. Однако, когда социальные партнеры женщин были похожи на MHC, они с большей вероятностью участвовали в отцовстве вне пары; в большинстве случаев самец внепары был значительно более отличен по MHC, чем социальный партнер. [ 32 ]
Выбор партнера, опосредованный MHC, может происходить после совокупления, на гаметическом уровне, посредством конкуренции сперматозоидов или загадочного выбора самки. Атлантический лосось — один из видов , Salmo salar у которого на конкуренцию сперматозоидов влияют изменения главного комплекса гистосовместимости, особенно аллелей класса I. Самцы атлантического лосося имеют более высокие показатели успешного оплодотворения при конкуренции за икру от самок, генетически сходных по генам класса I MHC. [ 33 ]
Еще одним видом, демонстрирующим загадочный выбор, связанный с MHC, является арктический гольц Salvelinus alpinus . Однако в этом случае кажется, что выбор сперматозоидов в большей степени зависит от яйцеклетки. Было обнаружено, что MHC-гетерозиготные самцы имеют значительно больший успех оплодотворения, чем MHC-гомозиготные самцы; Не было показано, что количество сперматозоидов, подвижность и скорость плавания существенно зависят от сходства или различия в MHC. Предполагается, что существует система хемоаттрактации, ответственная за то, что сама яйцеклетка способна различать и избирательно выбирать между MHC-гетерозиготными и MHC-гомозиготными самцами. [ 34 ]
В отличие от атлантического лосося и арктического гольца, рыжая джунглевая птица Gallus Gallus проявляет загадочное предпочтение самцов, а не самок. Самцы джунглевых птиц не проявили предпочтения, когда одновременно им были представлены как MHC-несходные, так и MHC-подобные самки. Тем не менее, они продемонстрировали загадочное предпочтение, отдав больше сперматозоидов тем из них, которые в большей степени отличались по MHC. [ 35 ]
Самцы песчаных ящериц Lacerta agilis ведут себя так же, как и самцы джунглевых птиц. Было показано, что первоначальное совокупление между самцом и самкой без каких-либо соперников продлевается, когда самец ощущает более высокую плодовитость самки. Однако вторые самцы корректировали продолжительность своего совокупления в зависимости от родства между самкой и первым самцом, которое, как полагают, определяется MHC-запахом копулятивной пробки. Более тесное генетическое родство между самцом и самкой песчаной ящерицы увеличивало шансы на успешное оплодотворение и вероятность отцовства второго самца. [ 36 ]
Абортивный отбор может быть формой загадочного женского выбора. Многие исследования на людях и грызунах показали, что самки могут самопроизвольно прерывать беременность, в которой потомство слишком похоже на MHC. [ нужна ссылка ] Кроме того, экстракорпоральное оплодотворение с большей вероятностью окажется неудачным, если пары имеют схожие гены MHC. [ нужна ссылка ]
MHC и сексуальный конфликт
[ редактировать ]Если самцы попытаются помешать выбору самки, спариваясь с самкой против ее воли, сексуальный конфликт может помешать выбору совместимости по генам MHC.
У чавычи Oncorhyncus tshawytscha самки действуют более агрессивно по отношению к MHC-подобным самцам, чем к MHC-несходным самцам, что предполагает наличие у самок выбора партнера. Более того, самцы направляли агрессию на MHC-подобных самок. Это сопровождалось притеснениями мужчин в отношении невосприимчивых женщин; однако существовала положительная корреляция между мужской агрессией и репродуктивным успехом. Способность самцов подавлять первоначальный выбор самки привела к тому, что потомство объектов мужской агрессии имело низкое генетическое разнообразие. Потомство с высоким генетическим разнообразием, по-видимому, появлялось только тогда, когда оперативное соотношение полов было ориентировано на самок, когда самки с большей вероятностью могли выбирать партнера, а самцы с меньшей вероятностью беспокоили самок. Эти результаты позволяют предположить, что сексуальный конфликт может мешать выбору партнера для женщин с «хорошими» генами MHC. [ 37 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Милински М., Гриффитс С., Вегнер К.М., Ройш Т.Б., Хаас-Ассенбаум А., Бём Т. (март 2005 г.). «Решения о выборе партнера самками колюшки предсказуемо модифицируются пептидными лигандами MHC» . Учеб. Натл. акад. наук. США 102 (12): 4414–8. Бибкод : 2005PNAS..102.4414M . дои : 10.1073/pnas.0408264102 . ПМК 555479 . ПМИД 15755811 .
- ^ Jump up to: а б с д и ж О'Двайер Т.В., Невитт Г.А. (июль 2009 г.). «Индивидуальное распознавание запахов у трубчатообразных цыплят: потенциальная роль главного комплекса гистосовместимости». Энн. Н-Й акад. наук. 1170 : 442–6. дои : 10.1111/j.1749-6632.2009.03887.x . ПМИД 19686174 . S2CID 10004939 .
- ^ Кемере, Эрван; Росси, Софи; Маленькая, Элоди; Маршан, Паскаль; Мерле, Джоэл; Игра, Иветт; Галан, Максим; Жило-Фромон, Эммануэль (10 марта 2020 г.). «Генетическая эпидемиология альпийского козерога – резервуара персистирующих и вирулентных вспышек бруцеллеза» . Научные отчеты . 10 (1): 4400. Бибкод : 2020NatSR..10.4400Q . дои : 10.1038/s41598-020-61299-2 . ISSN 2045-2322 . ПМК 7064506 . ПМИД 32157133 .
- ^ Миниас, Петр; Винклер, Михал (1 мая 2022 г.). «Баланс отбора генов врожденного иммунитета: поддержание адаптивного полиморфизма в толл-подобных рецепторах» . Молекулярная биология и эволюция . 39 (5): msac102. дои : 10.1093/molbev/msac102 . ISSN 1537-1719 . ПМК 9132207 . ПМИД 35574644 .
- ^ Моргер, Дженнифер; Байнок, Ярослав; Бойс, Келлиэнн; Крейг, Филип С.; Роган, Майкл Т.; Лунь, Чжао-Ронг; Спрячься, Джефф; Чиррен, Барбара (01 августа 2014 г.). «Природные полиморфизмы Toll-подобного рецептора 11 (TLR11) и Toll-подобного рецептора 12 (TLR12) не связаны с инфекцией Toxoplasma gondii у диких лесных мышей» . Инфекция, генетика и эволюция . 26 : 180–184. дои : 10.1016/j.meegid.2014.05.032 . ISSN 1567-1348 . ПМИД 24910107 .
- ^ Jump up to: а б Антонидес, Дженнифер; Матур, Самарт; Сундарам, Мекала; Риклефс, Роберт; ДеВуди, Дж. Эндрю (22 мая 2019 г.). «Иммуногенетический ответ бананаквита на фоне малярийных паразитов» . Эволюционная биология BMC . 19 (1): 107. Бибкод : 2019BMCEE..19..107A . дои : 10.1186/s12862-019-1435-y . ISSN 1471-2148 . ПМК 6529992 . ПМИД 31113360 .
- ^ Корнетти, Лука; Хилфикер, Даниэла; Лемуан, Мелисса; Чиррен, Барбара (6 августа 2018 г.). «Небольшие пространственные вариации риска заражения определяют эволюцию гена устойчивости к боррелиям у диких грызунов» . Молекулярная экология . 27 (17): 3515–3524. Бибкод : 2018MolEc..27.3515C . дои : 10.1111/mec.14812 . hdl : 10871/33429 . ISSN 0962-1083 . ПМИД 30040159 . S2CID 51711551 .
- ^ Нельсон-Флауэр, Марта Дж; Жермен, Райан Р.; Макдугалл-Шеклтон, Элизабет А; Тейлор, Сабрина С; Арсезе, Питер (27 июня 2018 г.). «Очищающий отбор в толл-подобных рецепторах певчих воробьев Melospiza melodia» . Журнал наследственности . 109 (5): 501–509. doi : 10.1093/jhered/esy027 . ISSN 0022-1503 . ПМИД 29893971 .
- ^ Ильмонен, Петтери; Пенн, Дастин Дж; Дамьянович, Кристи; Моррисон, Линда; Готби, Лалех; Поттс, Уэйн К. (1 августа 2007 г.). «Гетерозиготность по главному комплексу гистосовместимости снижает приспособленность у экспериментально инфицированных мышей» . Генетика . 176 (4): 2501–2508. doi : 10.1534/genetics.107.074815 . ISSN 1943-2631 . ЧВК 1950649 . ПМИД 17603099 .
- ^ Джо., Демас, Грегори Э. Нельсон, Рэнди (2012). Экоиммунология . Издательство Оксфордского университета. п. 238. ИСБН 978-0-19-987624-2 . OCLC 777401230 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Макклелланд, Эрин Э.; Грейнджер, Дональд Л.; Поттс, Уэйн К. (август 2003 г.). «Зависимая от комплекса гистосовместимости восприимчивость к Cryptococcus neoformans у мышей» . Инфекция и иммунитет . 71 (8): 4815–4817. дои : 10.1128/IAI.71.8.4815-4817.2003 . ISSN 0019-9567 . ПМК 166009 . ПМИД 12874366 .
- ^ Такахата, Наоюки (1994), Голдинг, Брайан (ред.), «Полиморфизм локусов MHC и изоляция иммунной системой у позвоночных» , Ненейтральная эволюция: теории и молекулярные данные , Бостон, Массачусетс: Springer US, стр. 233. –246, номер домена : 10.1007/978-1-4615-2383-3_19 , ISBN 978-1-4615-2383-3 , получено 28 июля 2022 г.
- ^ Новак, Массачусетс; Тарчи-Горнох, К; Остин, Дж. М. (15 ноября 1992 г.). «Оптимальное количество молекул главного комплекса гистосовместимости у человека» . Труды Национальной академии наук . 89 (22): 10896–10899. Бибкод : 1992PNAS...8910896N . дои : 10.1073/pnas.89.22.10896 . ISSN 0027-8424 . ПМК 50449 . ПМИД 1438295 .
- ^ Вулфинг, Бенно; Траульсен, Арне; Милински, Манфред; Бём, Томас (12 января 2009 г.). «Сохраняет ли внутрииндивидуальное разнообразие главного комплекса гистосовместимости золотую середину?» . Философские труды Королевского общества B: Биологические науки . 364 (1513): 117–128. дои : 10.1098/rstb.2008.0174 . ПМЦ 2666699 . ПМИД 18926972 .
- ^ Ламперт, КП; Фишер, П.; Шартл, М. (март 2009 г.). «Основная изменчивость комплекса гистосовместимости у клональной амазонской молли Poecilia formosa: число копий менее важно, чем генотип?» . Молекулярная экология . 18 (6): 1124–1136. Бибкод : 2009MolEc..18.1124L . дои : 10.1111/j.1365-294X.2009.04097.x . ПМИД 19226318 . S2CID 13068773 .
- ^ Шимкова, Андреа; Кошар, Мартин; Ветешник, Лукаш; Выскочилова, Мартина (14 июня 2013 г.). «Гены MHC и паразитизм у Carassius gibelio, диплоидно-триплоидного вида рыб с двойной стратегией воспроизводства» . Эволюционная биология BMC . 13 (1): 122. Бибкод : 2013BMCEE..13..122S . дои : 10.1186/1471-2148-13-122 . ISSN 1471-2148 . ПМЦ 3691641 . ПМИД 23768177 .
- ^ Вестемейер Р.Л., Браун Дж.Д., Симпсон С.А. и др. (ноябрь 1998 г.). «Отслеживание долгосрочного упадка и восстановления изолированного населения». Наука . 282 (5394): 1695–8. Бибкод : 1998Sci...282.1695W . дои : 10.1126/science.282.5394.1695 . ПМИД 9831558 . S2CID 19726112 .
- ^ Шлупп, Инго; Бербель-Фильо, Вальдир (3 июня 2021 г.). «Мнения преподавателей о метааналитических доказательствах того, что животные редко избегают инбридинга» . дои : 10.3410/ф.740048135.793586159 . S2CID 243521297 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Бернатчес, Л.; Лэндри, К. (май 2003 г.). «Исследования MHC на немодельных позвоночных: что мы узнали о естественном отборе за 15 лет?» . Журнал эволюционной биологии . 16 (3): 363–377. дои : 10.1046/j.1420-9101.2003.00531.x . ISSN 1010-061X . ПМИД 14635837 . S2CID 28094212 .
- ^ Jump up to: а б с Ямазаки К., Бошан Г.К., Сингер А., Бард Дж., Бойс Э.А. (февраль 1999 г.). «Типы запахов: их происхождение и состав» . Учеб. Натл. акад. наук. США 96 (4): 1522–5. Бибкод : 1999PNAS...96.1522Y . дои : 10.1073/pnas.96.4.1522 . ПМЦ 15502 . ПМИД 9990056 .
- ^ Бхутта М.Ф. (июнь 2007 г.). «Секс и нос: реакция феромонов человека» . JR Soc Med . 100 (6): 268–74. дои : 10.1177/014107680710000612 . ПМК 1885393 . ПМИД 17541097 .
- ^ Jump up to: а б Гавличек Дж., Робертс СК (май 2009 г.). «Выбор партнера у человека, коррелирующий с MHC: обзор». Психонейроэндокринология . 34 (4): 497–512. дои : 10.1016/j.psyneuen.2008.10.007 . ПМИД 19054623 . S2CID 40332494 .
- ^ Jump up to: а б Чайкс Р., Цао С., Доннелли П. (2008). «Зависит ли выбор партнера у людей от MHC?» . ПЛОС Генет. 4 (9): e1000184. дои : 10.1371/journal.pgen.1000184 . ПМК 2519788 . ПМИД 18787687 .
- ^ Jump up to: а б Робертс С.К., Гослинг Л.М., Картер В., Петри М. (декабрь 2008 г.). «Предпочтения запахов, коррелирующие с MHC, у людей и использование пероральных контрацептивов» . Учеб. Биол. наук. 275 (1652): 2715–22. дои : 10.1098/rspb.2008.0825 . ПМК 2605820 . ПМИД 18700206 .
- ^ Jump up to: а б Ведекинд К., Фюри С. (октябрь 1997 г.). «Предпочтения в запахах тела у мужчин и женщин: стремятся ли они к определенным комбинациям MHC или просто к гетерозиготности?» . Учеб. Биол. наук. 264 (1387): 1471–9. дои : 10.1098/rspb.1997.0204 . ПМК 1688704 . ПМИД 9364787 .
- ^ Швенсов Н., Эберле М., Зоммер С. (март 2008 г.). «Совместимость имеет значение: выбор партнера, связанный с MHC, у диких беспорядочных приматов» . Учеб. Биол. наук. 275 (1634): 555–64. дои : 10.1098/rspb.2007.1433 . ПМЦ 2596809 . ПМИД 18089539 .
- ^ Консуэгра С., Гарсия де Леанис С. (июнь 2008 г.). «Выбор партнера, опосредованный MHC, повышает устойчивость лосося к паразитам» . Учеб. Биол. наук. 275 (1641): 1397–403. дои : 10.1098/rspb.2008.0066 . ПМК 2602703 . ПМИД 18364312 .
- ^ Курц Дж., Кальбе М., Эшлиманн П.Б. и др. (январь 2004 г.). «Разнообразие главного комплекса гистосовместимости влияет на устойчивость к паразитам и врожденный иммунитет колюшек» . Учеб. Биол. наук. 271 (1535): 197–204. дои : 10.1098/rspb.2003.2567 . ПМК 1691569 . ПМИД 15058398 .
- ^ Олссон М., Мэдсен Т., Нордби Дж., Вапстра Э., Уйвари Б., Виттселл Х. (ноябрь 2003 г.). «Главный комплекс гистосовместимости и выбор партнера у песчаных ящериц» . Учеб. Биол. наук. 270 (Приложение 2): S254–6. дои : 10.1098/rsbl.2003.0079 . ПМК 1809963 . ПМИД 14667398 .
- ^ Сутер С.М., Кейзер М., Фейну Р., Мейер Д.Р. (ноябрь 2007 г.). «Самки тростниковой овсянки повышают приспособленность за счет спаривания дополнительных пар с генетически несходными самцами» . Учеб. Биол. наук. 274 (1627): 2865–71. дои : 10.1098/rspb.2007.0799 . ПМК 2288684 . ПМИД 17785270 .
- ^ Промерова Винклер. Возникновение внепарного отцовства связано с MHC-вариабельностью социального самца у алой чечевицы Carpodacus erythrinus. Журнал птичьей биологии 42, 5-10 (2011).
- ^ Ричардсон Д.С., Комдер Дж., Берк Т., фон Шанц Т. (апрель 2005 г.). «Модели социального выбора и выбора партнера вне пары на основе MHC у сейшельской славки» . Учеб. Биол. наук. 272 (1564): 759–67. дои : 10.1098/rspb.2004.3028 . ПМК 1602051 . ПМИД 15870038 .
- ^ Йейтс С.Е., Эйнум С., Флеминг И.А. и др. (февраль 2009 г.). «Икра атлантического лосося благоприятствует конкуренции сперматозоидов, имеющих схожие основные аллели гистосовместимости» . Учеб. Биол. наук. 276 (1656): 559–66. дои : 10.1098/rspb.2008.1257 . ПМЦ 2592554 . ПМИД 18854296 .
- ^ Скарштейн Ф. и др. (2005). «MHC и успех оплодотворения у арктического гольца (Salvelinus alpinus)». Поведенческая экология и социобиология . 57 (4): 374–380. дои : 10.1007/s00265-004-0860-z . S2CID 42727644 .
- ^ Джиллингем М.А., Ричардсон Д.С., Лёвли Х., Мойнихан А., Уорли К., Пиццари Т. (март 2009 г.). «Загадочное предпочтение самок, отличающихся по MHC, у самцов красной джунглевой птицы Gallus Gallus» . Учеб. Биол. наук. 276 (1659): 1083–92. дои : 10.1098/rspb.2008.1549 . ПМК 2679071 . ПМИД 19129124 .
- ^ Олссон М., Мэдсен Т., Уджвари Б., Вапстра Э. (апрель 2004 г.). «Плодовитость и MHC влияют на тактику эякуляции и предвзятость отцовства у песчаных ящериц». Эволюция . 58 (4): 906–9. дои : 10.1554/03-610 . hdl : 10536/DRO/DU:30066491 . ПМИД 15154566 . S2CID 198152712 .
- ^ Гарнер С.Р., Бортолуцци Р.Н., Хит Д.Д., Нефф Б.Д. (март 2010 г.). «Сексуальный конфликт препятствует выбору самки партнера из-за несходства главного комплекса гистосовместимости у чавычи» . Учеб. Биол. наук. 277 (1683): 885–94. дои : 10.1098/rspb.2009.1639 . ПМЦ 2842720 . ПМИД 19864282 .