Озерный метаболизм

Метаболизм озера представляет собой озера баланс между фиксацией углерода ( валовая первичная продукция ) и биологическим окислением углерода ( дыхание экосистемы ). [1] Метаболизм всего озера включает в себя фиксацию и окисление углерода всеми организмами в озере , от бактерий до рыб , и обычно оценивается путем измерения изменений растворенного кислорода или углекислого газа в течение дня. [2]
Дыхание экосистемы, превышающее валовую первичную продукцию, указывает на то, что озеро получает органический материал из окружающего водосбора , например, из ручьев , грунтовых вод притоков или опада . Метаболизм озер часто контролирует выбросы углекислого газа из озер или приток в них, но он не учитывает всю динамику углекислого газа, поскольку поступление неорганического углерода из окружающего водосбора также влияет на углекислый газ внутри озер. [3] [4]
Концепция
[ редактировать ]Оценки метаболизма озера обычно основаны на измерении растворенного кислорода или углекислого газа или измерениях углерода или индикатора кислорода для оценки производства и потребления органического углерода. Кислород вырабатывается, а углекислый газ потребляется в процессе фотосинтеза , кислород потребляется, а углекислый газ образуется в результате дыхания. Здесь органическое вещество символизируется глюкозой, хотя химические вещества, образующиеся и вдыхаемые в результате этих реакций, сильно различаются.

Фотосинтез :
![{\displaystyle 6CO_{2}+6H_{2}O{\xrightarrow[{}]{light}}C_{6}H_{12}O_{6}+6O_{2}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/c23155445a04b2b9c1afe475229a6274d87e38b7)
Дыхание :
![{\displaystyle C_{6}H_{12}O_{6}+6O_{2}{\xrightarrow[{}]{}}6CO_{2}+6H_{2}O}](https://wikimedia.org/api/rest_v1/media/math/render/svg/619bc6294a1bbc343eac76ab799d767420c04e83)
Фотосинтез и производство кислорода происходят только при наличии света , тогда как потребление кислорода посредством дыхания происходит как при наличии, так и при отсутствии света. Термины метаболизма озера включают:
- GPP - валовая первичная продукция (например, общий фотосинтез)
- Р – общее дыхание
- - гетеротрофное дыхание
- - автотрофное дыхание
- NEP – чистая продукция экосистемы = GPP – R
- ЧПП – чистая первичная продукция = ГПП –



Методы измерения
[ редактировать ]Оценка метаболизма озера требует аппроксимации процессов, которые влияют на производство и потребление органического углерода организмами в озере. Циклические изменения в ежедневном масштабе происходят в большинстве озер на Земле, поскольку солнечный свет доступен для фотосинтеза и производства нового углерода только в течение части дня. Исследователи могут воспользоваться этой моделью суточных графиков для измерения скорости изменения самого углерода или изменений в растворенных газах, таких как углекислый газ или кислород, которые происходят в ежедневном масштабе. Хотя ежедневные оценки метаболизма являются наиболее распространенными, метаболизм всего озера можно интегрировать за более длительные периоды времени, такие как сезонные или годовые показатели, путем оценки углеродного баланса всего озера . В следующих разделах освещаются наиболее распространенные способы оценки метаболизма озер в различных временных и пространственных масштабах, а также рассматриваются некоторые допущения каждого из этих методов.

Методы свободной воды
[ редактировать ]Измерение суточных изменений растворенных газов в озере, также известное как метод «свободной воды», быстро стало наиболее распространенным методом оценки метаболизма озера после широкого внедрения автономных датчиков, используемых для измерения растворенного кислорода и углекислого газа в воде. . [6] [7] [8] Метод свободной воды особенно популярен, поскольку многие ежедневные оценки метаболизма озера можно собрать относительно дешево и дать представление о метаболических режимах в трудные для наблюдения периоды времени, например, во время штормов. Измеренные изменения содержания растворенного кислорода и углекислого газа в озере представляют собой сумму всего метаболизма организма, от бактерий до рыб, после учета абиотических изменений растворенных газов. Абиотические изменения растворенных газов включают обмен растворенными газами между атмосферой и поверхностью озера, вертикальный или горизонтальный унос воды с различными концентрациями (например, вода с низким содержанием кислорода ниже термоклина озера) или импорт и экспорт растворенных газов из впадающих потоков или выход из озера. Абиотические изменения в растворенных газах могут доминировать над изменениями в растворенных газах, если озеро имеет низкую скорость метаболизма (например, олиготрофное озеро, пасмурный день) или если происходит крупное событие, которое приводит к тому, что абиотические факторы превышают биотические (например, ветер, вызывающий перемешивание и унос воды с низким содержанием кислорода). Биотические сигналы в растворенных газах наиболее очевидны, когда светит солнце и происходит фотосинтез, приводящий к выработке растворенного кислорода и потреблению углекислого газа. Преобразование солнечной энергии в химическую энергию называется валовым первичным производством (GPP), а рассеивание этой энергии посредством биологического окисления углерода называется дыханием экосистемы (ER). Высокочастотные (например, с 10-минутным интервалом) измерения растворенного кислорода или углекислого газа можно перевести в оценки GPP, ER и разницы между двумя показателями, называемыми чистой экосистемной продукцией (NEP), путем сопоставления высокочастотных данных с моделями Озерный метаболизм. Основное уравнение для оценки метаболизма озера с помощью одного датчика, расположенного в верхнем перемешанном слое, измеряющего растворенный кислород:
ДО/ч = ГПП-ЭР+Ф
Где F – поток газов между озером и атмосферой. Дополнительные условия потока абиотического газа могут быть добавлены, если эти абиотические потоки считаются значимыми для озера (например, явления смешивания, приток газов потока). Атмосферный газообмен (F) редко измеряется напрямую и обычно моделируется путем оценки турбулентности поверхности озера в результате ветрового и конвективного перемешивания. Чаще всего F оценивается на основе измерений скорости ветра и атмосферного давления, а разные модели оценки F могут привести к существенно разным оценкам скорости метаболизма озера в зависимости от исследуемого озера. [9] Предполагается, что валовая первичная продукция равна нулю в ночное время из-за слабого освещения или его отсутствия, и, таким образом, ER можно оценить по изменениям растворенного кислорода (или углекислого газа) в ночное время после учета абиотических изменений растворенного кислорода. Валовую первичную продукцию можно оценить, предполагая, что ER одинаков в течение дня и ночи, и учитывая изменения растворенного кислорода в течение дня, однако это предположение может быть справедливым не для каждого озера. [10]
Получение высокого отношения сигнал/шум является ключом к получению точных оценок метаболизма озера с помощью метода свободной воды, и есть выбор, который исследователь должен сделать до сбора данных и во время анализа данных, чтобы обеспечить точные оценки. Место сбора растворенного газа (обычно в поверхностном перемешанном слое), количество датчиков по вертикали и горизонтали, [11] [12] [13] Необходимо учитывать частоту и продолжительность сбора данных, а также методы моделирования. [14]
Методы моделирования метаболизма свободной воды
[ редактировать ]Методы измерения свободной воды требуют математических моделей для оценки показателей метаболизма озера на основе высокочастотных измерений растворенного газа. Эти модели различаются по сложности: от простых алгебраических моделей до глубоко интегрированного моделирования с использованием более совершенных статистических методов. Для оценки GPP, R и NEP или параметров, относящихся к этим терминам метаболизма, использовалось несколько статистических методов.
Методы светлых и темных бутылок
[ редактировать ]Метод светлой и темной бутылки использует ту же концепцию, что и метод свободной воды, для оценки скорости метаболизма - GPP происходит только в течение дня при использовании солнечной энергии, тогда как ER происходит как при наличии, так и при отсутствии света. [15] В этом методе озерная вода инкубируется в двух отдельных бутылках: одна прозрачная и подвергается воздействию естественного или искусственного освещения, а другая изолируется от света путем обертывания бутылки фольгой, краской или другим способом. Затем в течение определенного периода времени (например, от нескольких часов до суток) измеряются изменения в фиксации углерода или растворенных газов, чтобы оценить скорость метаболизма для конкретной глубины озера или объединенной водной толщи озера. Фиксация углерода измеряется путем введения радиоактивного изотопа углерода. 14 С в светлые и темные бутылки и отбор проб из бутылей с течением времени – пробы фильтруют на фильтровальную бумагу и количество 14 Уровень углерода, включенный в клетки водорослей (и бактерий), оценивается путем измерения образцов на сцинтилляционном счетчике. Разница между светлой и темной бутылкой 14 С можно считать показателем первичной продуктивности; однако из-за нефотосинтетического поглощения CO 2 ведутся споры о том, следует ли использовать темные бутылки с 14 только легкую бутылку и бутылку, обработанную альгицидом DCMU Метод C или следует использовать . Скорость изменения растворенных газов, углекислого газа или кислорода, требует использования как светлых, так и темных бутылей для оценки скорости продуктивности и дыхания.
Методы углеродного баланса всего озера
[ редактировать ]Вероятно, наиболее трудоемким методом оценки показателей метаболизма озера является измерение всех поступлений и выбросов органического или неорганического углерода в озеро за сезон или год, что также известно как баланс углерода всего озера. Измерение всех поступлений и выбросов углерода в озеро и из озера можно использовать для оценки чистой продукции экосистемы (NEP). [16] [17] Поскольку NEP представляет собой разницу между валовым первичным производством и дыханием (NEP = GPP - R), его можно рассматривать как чистое биологическое преобразование неорганического углерода в органический углерод (и наоборот) и, таким образом, можно определить через массу всего озера. баланс неорганического или органического углерода. [16] NEP, оцененный по неорганическому (IC) или органическому углероду (OC), можно оценить как:


где E – экспорт ОС посредством речного переноса, а IC – через речной перенос и обмен углекислого газа (например, CO 2 , CH 4 ) между поверхностью озера и атмосферой ; S – запасы в озерных отложениях и толще воды ОС и толще воды ИЦ; и I представляет собой поступление OC и IC из речных вод, окружающих водно-болотных угодий и по воздуху (например, атмосферные осаждения , опад ). Озеро, которое получает больше OC из водораздела , чем экспортирует вниз по течению или накапливается в толще воды и отложениях (I oc > E oc + S oc ), указывает на то, что внутри озера произошло чистое преобразование OC в IC и, таким образом, оно является чистым гетеротрофным ( отрицательный НЭП). Аналогичным образом, озеро, которое накапливает и экспортирует больше IC, чем было получено из водораздела (S ic + E ic > I ic ), также указывает на чистое преобразование OC в IC внутри озера и, таким образом, является чистым гетеротрофным.
Методы донного метаболизма
[ редактировать ]Хотя метод свободной воды, вероятно, содержит некоторый сигнал бентосного метаболизма, выделение бентосного вклада в метаболизм всего озера требует методов, специфичных для бентоса. Аналогично описанным выше методам использования светлых и темных бутылок, можно собирать керны озерных отложений и использовать изменения в фиксации растворенного кислорода или углерода для оценки показателей первичной продуктивности и дыхания. Относительно новые методы описывают изоляцию границы раздела осадок-вода с помощью прозрачных куполов и измерение изменений содержания растворенного кислорода на месте, что представляет собой гибрид метода свободной воды и метода светло-темной бутылки. [18] Эти методы в бентосной камере in-situ позволяют относительно легко оценить метаболизм бентоса в течение нескольких дней, что помогает исследователю определить, как меняется метаболизм бентоса при изменении погодных условий и характеристик озера.
Предположения
[ редактировать ]Экстраполяция измерений, специфичных для участка или глубины, на все озеро может быть проблематичной, поскольку внутри озера могут наблюдаться значительные метаболические изменчивости как по вертикали, так и по горизонтали. [11] (см. раздел изменчивости). Например, многие исследования метаболизма озер имеют только одну эпилимнетическую оценку метаболизма, однако это может привести к переоценке метаболических характеристик озера, таких как NEP, в зависимости от отношения глубины смешанного слоя к глубине затухания света. [12] [19] Усреднение ежедневных оценок метаболизма за более длительные периоды времени может помочь преодолеть некоторые из этих проблем экстраполяции одного сайта. [11] но необходимо тщательно учитывать последствия метаболических оценок, а не экстраполировать измерения.
Отношение к избирателям
[ редактировать ]На скорость метаболизма организма, или скорость, с которой организмы ассимилируют, трансформируют и расходуют энергию, влияют несколько ключевых составляющих, а именно свет, питательные вещества, температура и органические вещества. Влияние этих компонентов на метаболизм организма в конечном итоге определяет метаболизм в масштабе всего озера и может определять, является ли озеро чистым источником или поглотителем углерода. В следующем разделе мы описываем взаимосвязь между этими ключевыми компонентами и метаболизмом на уровне организма и экосистемы. Хотя отношения между описанными здесь организмами и компонентами хорошо известны, взаимодействующее влияние компонентов на скорость метаболизма от организмов до озерных экосистем затрудняет прогнозирование изменений метаболизма между озерами или внутри озер с течением времени. Многие из этих сложных взаимодействующих эффектов будут обсуждаться в разделе о пространственной и временной изменчивости.
Температура
[ редактировать ]
Температура является сильным фактором, контролирующим скорость биохимических реакций и биологическую активность. Оптимальная температура варьируется для разных водных организмов, поскольку некоторые организмы более адаптированы к холоду, а другие предпочитают более теплую среду обитания. Встречаются редкие случаи крайней термической толерантности в гиперсоленых антарктических озерах (например, пруд Дон Хуан ) или горячих источниках (например, Флай Гейзер ); однако большинство озерных организмов на Земле обитают при температуре от 0 до 40 ° C. Скорость метаболизма обычно экспоненциально зависит от температуры, однако энергия активации первичной продуктивности и дыхания часто различается, при этом фотосинтез имеет более низкую энергию активации, чем аэробное дыхание. Эти различия в энергиях активации могут иметь последствия для чистого метаболического баланса в озерных экосистемах по мере потепления климата. Например, Шарфенбергер и др. (2019) [20] показывают, что повышение температуры воды в результате изменения климата может переключить озера из чистого автотрофного состояния в гетеротрофное из-за различий в энергии активации, однако температура, при которой они переключаются, зависит от количества доступных питательных веществ.
Питательные вещества
[ редактировать ]Количество материала, доступного для усвоения клетками организма, контролирует скорость метаболизма на уровне клеточной и озерной экосистемы. В озерах фосфор и азот являются наиболее распространенными лимитирующими питательными веществами первичной продукции и дыхания экосистемы. ограничивал количество фосфора в стиральных порошках . Фундаментальная работа по изучению положительной взаимосвязи между концентрацией фосфора и эвтрофикацией озер привела к принятию закона, который , помимо других правил, [21] [22] Хотя фосфор часто используется в качестве показателя продуктивности экосистемы озера, а избыток фосфора — в качестве индикатора эвтрофикации, многие исследования показывают, что метаболизм совместно ограничивается фосфором и азотом или только азотом. [23] Баланс между фосфором, азотом и другими питательными веществами, называемый экологической стехиометрией , может определять скорость роста организма и метаболизм всего озера посредством клеточных потребностей в этих важнейших питательных веществах, опосредованных особенностями жизненного цикла. Например, быстрорастущие ветвистоусые имеют гораздо более низкое соотношение азота и фосфора (N:P), чем копеподы , в основном из-за большого количества богатой фосфором РНК в их клетках, используемой для быстрого роста. Владоцеры, обитающие в озерах с высоким соотношением N:P относительно стехиометрии тела кладоцер, будут ограничены в росте и метаболизме, что будет влиять на метаболизм всего озера. Кроме того, каскадные эффекты манипуляций с пищевой сетью могут вызвать изменения в продуктивности из-за изменений в стехиометрии питательных веществ. Например, добавление рыбоядных животных может снизить давление хищников на быстрорастущие кладоцеры с низким содержанием N:P, популяция которых быстро увеличивается, сохраняет фосфор в своих клетках и может привести к ограничению фосфора в озере, что, как следствие, снижает первичную продуктивность всего озера.
Свет
[ редактировать ]Солнечная энергия необходима для преобразования углекислого газа и воды в органическое вещество, иначе известного как фотосинтез. Как и в случае с температурой и питательными веществами, разные водоросли имеют разную скорость метаболической реакции на увеличение освещения, а также разные оптимальные условия освещения для роста, поскольку некоторые водоросли более приспособлены к более темной среде, в то время как другие могут вытеснить их в более светлых условиях. Свет также может взаимодействовать с питательными веществами, влияя на реакцию продуктивности водорослей на увеличение освещенности. [24] Эти различные реакции на уровне организма распространяются и влияют на метаболизм на уровне экосистемы. [25] [26] Даже в озерах с низким содержанием питательных веществ , где питательные вещества, как ожидается, будут ограничивающим ресурсом для первичной продуктивности, свет все еще может быть ограничивающим ресурсом, что оказывает каскадное негативное воздействие на более высокие трофические уровни, такие как продуктивность рыбы. [27] Изменчивость освещенности в разных зонах озера и внутри озера с течением времени создает неоднородность продуктивности как в пространственном, так и во временном отношении.
Помимо контроля первичной продуктивности, солнечный свет также может влиять на скорость дыхания, частично окисляя органическое вещество, что облегчает бактериям расщепление и преобразование в углекислый газ. Это частичное фотоокисление существенно увеличивает количество органического вещества, доступного для минерализации. [28] В некоторых озерах полное или частичное фотоокисление может составлять большую часть превращения органического вещества в неорганическое, однако доля бактериального дыхания сильно варьируется в зависимости от озера.
Органический углерод
[ редактировать ]Первичным и вторичным потребителям в озерах необходимы органические вещества (растительного или животного происхождения) для поддержания функций организма. Органические вещества, включая листья деревьев, растворенные органические вещества и водоросли, предоставляют этим потребителям необходимые ресурсы и в процессе увеличивают скорость дыхания экосистемы озера за счет преобразования органических веществ в рост клеток и поддержание организма. Некоторые источники органических веществ могут влиять на доступность других компонентов. Например, растворенные органические вещества часто затемняют воду озера, что уменьшает количество света, доступного в озере, тем самым снижая первичную продукцию. Однако увеличение нагрузки органического вещества в озеро может также увеличить количество питательных веществ, связанных с органическим веществом, что может стимулировать первичное производство и дыхание. Повышенное содержание растворенных органических веществ может создать компромисс между увеличением ограничения света и освобождением от ограничения питательных веществ. Этот компромисс может создать нелинейные зависимости между первичной продуктивностью озера и нагрузкой растворенного органического вещества в зависимости от того, сколько питательных веществ связано с органическим веществом и насколько быстро растворенное органическое вещество блокирует свет в толще воды. [29] [30] [31] [32] Это связано с тем, что при низких концентрациях растворенного органического вещества по мере увеличения концентрации растворенного органического вещества увеличение количества связанных с ним питательных веществ увеличивает GPP. [29] [30] [31] Но поскольку количество растворенных органических веществ продолжает увеличиваться, уменьшение освещенности из-за потемнения воды в озере подавляет GPP, поскольку свет становится ограничивающим ресурсом для первичной продуктивности. [29] [30] [31] Предполагается, что различия в величине и расположении максимального GPP в ответ на повышенную нагрузку DOC возникают на основе соотношения DOC и питательных веществ, поступающих в озеро, а также влияния DOC на световой климат озера. [31] [33] Потемнение воды в озере также может изменить термический режим внутри озера, поскольку более темные воды обычно означают, что более теплые воды остаются в верхней части озера, а более прохладные — внизу. Это изменение в распределении тепловой энергии может повлиять на уровень продуктивности пелагических и бентосных организмов (см. «Температура» выше) и изменить стабильность водного столба, оказывая влияние на вертикальное распределение питательных веществ и, следовательно, оказывая влияние на вертикальное распределение скорости метаболизма.

Другие составляющие
[ редактировать ]Другие компоненты озера могут влиять на скорость метаболизма озера, включая, среди прочего, концентрацию CO 2 , pH, соленость и кремнезем. CO 2 может быть лимитирующим (или соограничивающим наряду с другими питательными веществами) ресурсом для первичной продуктивности. [35] и может способствовать более интенсивному цветению фитопланктона. [36] Некоторые виды водорослей, такие как хризофиты, могут не обладать механизмами концентрации углерода или способностью использовать бикарбонат в качестве источника неорганического углерода для фотосинтеза, поэтому повышенные уровни CO 2 могут увеличивать скорость их фотосинтеза. Во время цветения водорослей повышенное содержание растворенного CO 2 гарантирует, что CO 2 не является ограничивающим ресурсом для роста, поскольку быстрое увеличение производства истощает CO 2 и повышает pH. Изменения pH в коротких временных масштабах (например, субдневные) из-за скачков первичной продуктивности могут вызвать кратковременное снижение роста и дыхания бактерий, но в более длительных временных масштабах бактериальные сообщества могут адаптироваться к повышенному pH. [37] [38]
Соленость также может вызывать изменения в скорости метаболизма озер из-за воздействия солености на индивидуальную скорость метаболизма и состав сообщества. [39] [40] [41] Скорость метаболизма в озере может коррелировать как положительно, так и отрицательно с соленостью из-за взаимодействия солености с другими факторами метаболизма экосистемы, такими как скорость смыва или засухи. [42] Например, Морейра-Тюрк (2000). [43] обнаружили, что избыток осадков по сравнению с испарением приводит к снижению солености в прибрежной лагуне, увеличению нагрузки питательными веществами и увеличению первичной пелагической продуктивности. Положительная взаимосвязь между первичной продуктивностью и соленостью может быть индикатором изменений в доступности питательных веществ из-за увеличения притока. Однако соленость увеличивается из-за дорожных солей. [44] может вызвать токсичность у некоторых озерных организмов, [45] а крайние случаи повышения солености могут ограничить перемешивание озер, что может изменить распределение скорости метаболизма в водной толще озера.
Пространственная и временная изменчивость
[ редактировать ]Скорость метаболизма в озерах и водохранилищах контролируется многими факторами окружающей среды, такими как доступность света и питательных веществ, температура и режим перемешивания водного столба. Таким образом, пространственные и временные изменения этих факторов вызывают пространственную и временную изменчивость скорости метаболизма, и каждый из этих факторов влияет на метаболизм в разных пространственных и временных масштабах.
Пространственные вариации внутри озер
[ редактировать ]Переменный вклад различных озерных зон (т.е. литоральной , пресноводной , бентосной ) в метаболизм всего озера зависит главным образом от неоднородности биомассы водорослей и бактерий, а также от наличия света и питательных веществ. Что касается организмов, участвующих в обмене веществ в каждой из этих зон, в метаболизме лимнетиков преобладают фитопланктон, зоопланктон и бактериальный метаболизм с низким вкладом эпифитов и рыб. Бентосный метаболизм может получать большой вклад от макрофитов , макро- и микроводорослей, беспозвоночных и бактерий. Бентосный метаболизм обычно наиболее высок в мелководных прибрежных зонах или в мелководных озерах с прозрачной водой, в которых свет достигает дна озера и стимулирует первичную продукцию. В темных или мутных глубоких озерах первичная продукция может быть ограничена более мелкими водами, а аэробное дыхание может быть снижено или вообще отсутствовать в более глубоких водах из-за образования бескислородных глубоких зон.
Степень пространственной неоднородности скорости метаболизма внутри озера зависит от морфометрии озера, характеристик водосбора (например, различий в землепользовании по всему водосбору и поступлениях из ручьев) и гидродинамических процессов. Например, озера с более интенсивными гидродинамическими процессами, такими как сильное вертикальное и латеральное перемешивание, более однородны по латерали и вертикали в отношении скорости метаболизма, чем сильно стратифицированные озера. С другой стороны, озера с более развитой литоралью имеют большую метаболическую гетерогенность по латерали, чем озера с более круглой формой и низкой долей мелководных литоралей.
Ослабление света, происходящее в толще воды, в сочетании с термической и химической стратификацией и турбулентностью, вызванной ветром или конвекцией, способствуют вертикальному распределению питательных веществ и организмов в толще воды. В стратифицированных озерах органическое вещество и питательные вещества, как правило, более сконцентрированы в более глубоких слоях, тогда как свет более доступен в более мелких слоях. Вертикальное распределение первичной продукции отвечает балансу между доступностью света и питательных веществ, в то время как дыхание происходит более независимо от света и питательных веществ и более однородно с глубиной. [46] Это часто приводит к сильной связи валовой первичной продукции (GPP) и дыхания экосистемы (ER) в поверхностных слоях озера, но к более слабой связи на больших глубинах. Это означает, что скорость ЭР сильно зависит от первичной продукции в более мелких слоях, тогда как в более глубоких слоях она становится более зависимой от смеси органического вещества из наземных источников и осаждения частиц водорослей и органического вещества, образующегося в более мелких слоях. В озерах с низкой концентрацией питательных веществ в поверхностных водах и с проникновением света под перемешанный слой первичная продукция выше на промежуточных глубинах, где достаточно света для фотосинтеза и более высокая доступность питательных веществ. [46] С другой стороны, малопрозрачные полимиктические озера имеют более высокую первичную продукцию в приповерхностных слоях, обычно с чистым автотрофным балансом (GPP > ER) между первичной продукцией и дыханием. [12]
С латеральной точки зрения неоднородность озер обусловлена различиями в скорости метаболизма в лимнетических зонах открытой воды и литоральных зонах с более преобладанием бентоса. Прибрежные районы обычно более сложны и неоднородны, отчасти из-за их близости к наземной системе, но также из-за небольшого объема воды и высокого соотношения объема наносов к воде. Таким образом, прибрежные зоны более восприимчивы к изменениям температуры, поступлению питательных веществ и органических веществ из ландшафта и речных притоков, сдвиговому перемешиванию ветра и воздействию волн, затенению наземной растительности и взвешиванию отложений (рис. 1). Кроме того, прибрежные зоны обычно имеют более сложную среду обитания из-за присутствия макрофитов, которые служат убежищем, рассадником и местом питания для многих организмов. Следовательно, скорость метаболизма в прибрежных районах обычно имеет высокую краткосрочную изменчивость и обычно превышает скорость метаболизма в озерных водах. [47] [11]
Пространственные различия между озерами
[ редактировать ]Помимо пространственной изменчивости внутри озер, скорость метаболизма в целом и его движущие силы также различаются в зависимости от озера. Каждое озеро имеет уникальный набор характеристик в зависимости от его морфометрии, свойств водосбора и гидрологических характеристик. Эти особенности влияют на условия озера, такие как цвет воды, температура, питательные вещества, органические вещества, ослабление света, вертикальное и горизонтальное перемешивание, оказывая прямое и косвенное воздействие на метаболизм озера.
Поскольку озера различаются по состоянию их составляющих (например, освещенности, питательных веществ, температуры и органического вещества), возникают различия в величине и изменчивости скорости метаболизма между озерами. В предыдущем разделе ( «Связь с составляющими ») мы обсудили ожидаемые закономерности скорости метаболизма в ответ на изменчивость этих влиятельных составляющих. Здесь мы обсудим, как метаболизм всего озера варьируется в разных озерах из-за различий в этих компонентах, опосредованных различиями в морфометрии озера, свойствах водосбора и времени пребывания воды .
Морфометрия озера (например, размер и форма озера) и свойства водосбора (например, землепользование, площадь водосбора, климат и геологические характеристики) определяют поток внешних поступлений органических веществ и питательных веществ на единицу объема воды озера. По мере увеличения соотношения между размером водосбора и объемом воды в озере (коэффициент дренажа) поток питательных веществ и органических веществ из окружающего наземного ландшафта обычно увеличивается. [48] То есть небольшие озера с относительно большими водосборами будут получать больше внешних поступлений питательных веществ и органических веществ на единицу объема озера, чем большие озера с относительно небольшими водосборами, что приведет к увеличению как первичной продукции, так и скорости дыхания. Ожидается, что в озерах с небольшим коэффициентом дренажа (т.е. с относительно большой площадью поверхности озера по отношению к площади водосбора) метаболические процессы будут меньше зависеть от внешних факторов, поступающих из окружающего водосбора. Кроме того, небольшие озера менее подвержены ветровому перемешиванию и, как правило, имеют более высокий уровень поступления наземного органического вещества, что часто приводит к меньшей глубине смешивания и усиленному ослаблению света, что ограничивает первичную продукцию верхними частями небольших озер. Что касается озер со схожими водосборными свойствами, то небольшие озера, как правило, более гетеротрофны (GPP < ER), чем большие озера, поскольку их более высокая скорость дыхания подпитывается более высоким содержанием аллохтонного органического вещества (т.е. синтезируемого внутри водосборной площади, но за пределами водоема). поступает в систему и опережает первичную добычу, которая ограничена более мелкими слоями озер.

Свойства водосбора, а именно земельный покров, землепользование и геологические характеристики, влияют на метаболизм озера посредством воздействия на качество органического вещества и питательных веществ, поступающих в озеро, а также воздействия ветра. Качество органического вещества может влиять на ослабление света и, наряду с воздействием ветра, на распределение тепла и света по всей толще воды озера. Озера в ландшафтах, где преобладает сельское хозяйство, имеют более высокие поступления питательных веществ и более низкие поступления органических веществ по сравнению с озерами с аналогичным коэффициентом дренажа, но в ландшафтах, где преобладают леса. Таким образом, ожидается, что озера в ландшафтах с преобладанием сельского хозяйства будут иметь более высокие темпы первичной продукции, большее цветение водорослей и чрезмерную биомассу макрофитов по сравнению с озерами в ландшафтах с преобладанием лесов ( рисунок ). Однако влияние размера и типа водосбора является сложным и интерактивным. Относительно небольшие лесные озера более затенены и защищены от воздействия ветра, а также получают большое количество аллохтонного органического вещества. Таким образом, небольшие лесные озера, как правило, более гуминовые с неглубоким перемешанным слоем и пониженной светопроницаемостью. Высокие поступления аллохтонного органического вещества (продуцируемого за пределами озера) стимулируют гетеротрофные сообщества, такие как бактерии, зоопланктон и рыбы, повышая скорость дыхания во всем озере. Следовательно, небольшие лесные озера с большей вероятностью будут чистыми гетеротрофными, при этом темпы ER превышают темпы первичной продукции в озере. С другой стороны, лесные озера с низким коэффициентом дренажа получают относительно меньше питательных веществ и органических веществ, что обычно приводит к образованию озер с чистой водой и низкими показателями GPP и ER ( таблица ).

Еще одним важным различием между озерами, которое влияет на изменчивость метаболизма озер, является время пребывания воды в системе, особенно среди озер, которые интенсивно управляются человеком . Изменения уровня озера и скорости смыва влияют на концентрации питательных веществ и органических веществ, численность организмов и скорость экологических процессов, таких как фотодеградация окрашенных органических веществ , тем самым влияя на величину и изменчивость скорости метаболизма. Бессточные озера или озера с промежуточным гидравлическим временем пребывания (HRT) обычно имеют высокое время удержания питательных веществ и органических веществ в системе, что способствует росту первичных продуцентов и бактериальной деградации органических веществ. [49] Таким образом, ожидается, что эти типы озер будут поддерживать относительно более высокие и менее изменчивые показатели GPP и ER, чем озера с коротким временем пребывания в том же трофическом статусе. С другой стороны, ожидается, что в озерах с длительной ГГТ скорость метаболизма будет снижена из-за меньшего поступления в озеро питательных веществ и органических веществ. Наконец, лентические системы, которые имеют частые и интенсивные изменения уровня воды и ускоренную скорость промывки, имеют динамику, близкую к лоточным системам , с обычно низкими показателями GPP и ER из-за того, что питательные вещества, органические вещества и водоросли вымываются из системы во время интенсивного смыва. события промывки.
Временные вариации в суточном масштабе
[ редактировать ]В суточном масштабе на показатели GPP больше всего влияет суточный цикл фотосинтетически активной радиации, тогда как на ER в значительной степени влияют изменения температуры воды. [50] Кроме того, скорость ER также связана с количеством или качеством органического субстрата и относительным вкладом автотрофного и гетеротрофного дыхания, о чем свидетельствуют исследования закономерностей ночного дыхания (например, Sadro et al. 2014). [10] ). Например, дыхание бактериопланктона может быть выше в течение дня и в первые часы ночи из-за большей доступности лабильного растворенного органического вещества, вырабатываемого фитопланктоном. С восходом солнца в озере быстро увеличивается первичная продукция, что часто делает его автотрофным (NEP > 0) и снижает содержание растворенного CO 2 , образовавшегося в результате минерализации углерода, произошедшей в ночное время. Такое поведение продолжается до тех пор, пока не будет достигнут пик NEP, обычно около максимальной доступности света. Кроме того, существует тенденция к неуклонному снижению NEP между часами максимальной освещенности и до восхода солнца следующего дня.
Ежедневные различия в поступающем свете и температуре из-за различий в погоде, таких как облачность и штормы, влияют на скорость первичной продукции и, в меньшей степени, на дыхание. [51] Эти погодные изменения также вызывают кратковременную изменчивость глубины смешанного слоя, что, в свою очередь, влияет на питательные вещества, органическое вещество и доступность света, а также на вертикальный и горизонтальный газообмен. Глубокое перемешивание снижает доступность света, но также увеличивает доступность питательных веществ и органических веществ в верхних слоях. Таким образом, влияние краткосрочной изменчивости глубины смешанного слоя на валовую первичную продукцию (ВПП) будет зависеть от того, какие факторы являются ограничивающими для каждого озера в данный период. Таким образом, более глубокий слой смешивания может либо увеличить, либо уменьшить скорость GPP в зависимости от баланса между питательными веществами и ограничением фотосинтеза светом ( рис. ).
Реакция скорости метаболизма столь же динамична, как физические и химические процессы, происходящие в озере, но изменения биомассы водорослей менее изменчивы, включая рост и потерю в течение более длительных периодов. Высокая доступность света и питательных веществ связана с образованием цветения водорослей в озерах; во время этого цветения ставки GPP очень высоки, и ставки ER обычно увеличиваются почти так же, как и ставки GPP, а баланс GPP и ER близок к 1. Сразу после цветения ставки GPP начинают снижаться, но ставки ER продолжают повышаться из-за высокая доступность лабильных органических веществ, что может привести к быстрому снижению концентрации растворенного кислорода в толще воды, что приведет к гибели рыбы.
Временные вариации в годовом масштабе
[ редактировать ]Сезонные изменения в метаболизме могут быть вызваны сезонными изменениями температуры, ледяного покрова, осадков, смешивания и стратификации динамики , а также сукцессии сообществ (например, контроль фитопланктона с помощью зоопланктона). [52] ). Сезонные изменения в метаболизме озер будут зависеть от того, как времена года изменяют поступление питательных веществ и органических веществ, а также от доступности света, а также от того, какие факторы ограничивают скорость метаболизма в каждом озере.
Свет является основным фактором метаболизма озера, поэтому сезонность уровня освещенности является важным фактором сезонных изменений скорости метаболизма озера. Таким образом, ожидается, что показатели GPP будут более выраженными в такие сезоны, как весна и лето, когда уровень освещенности выше, а дни длиннее. Особенно это выражено для озер с ограниченным светом ГПП, например, более мутных или окрашенных озер. Сезонность уровня освещенности также влияет на показатели ER. Скорость дыхания экосистемы обычно сочетается со скоростью GPP, поэтому в сезоны с более высоким GPP также будут наблюдаться более высокие скорости ER, связанные с увеличением количества органического вещества, производимого в озере. Кроме того, в сезоны с более высоким уровнем освещенности фотодеградация органического вещества более выражена, что стимулирует микробную деградацию, усиливая интенсивность гетеротрофного дыхания.

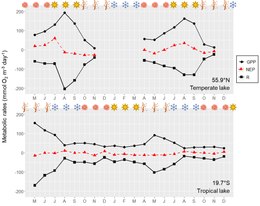
Большинство озер в мире замерзают зимой. [53] период низкой освещенности, когда лед и снежный покров ограничивают проникновение света в толщу воды. Ограничение освещенности происходит главным образом из-за снежного покрова, а не льда, что делает первичную продукцию очень чувствительной к снежному покрову в этих озерах. [54] Помимо ограничения света, низкие температуры подо льдом также снижают скорость метаболизма, но недостаточно, чтобы остановить метаболические процессы. Таким образом, метаболический баланс обычно отрицательный в течение большей части ледового сезона, что приводит к истощению растворенного кислорода. Мелкие озера в засушливом климате зимой не имеют снежного покрова или имеют очень небольшой снежный покров, таким образом, устойчивое первичное производство подо льдом может быть достаточным для предотвращения истощения растворенного кислорода, как сообщили Сонг и другие. [54] в монгольском озере. Несмотря на высокую долю озер в мире, которые замерзают зимой, исследований метаболизма озер под льдом проводилось мало, в основном из-за технических трудностей с отбором проб. [53] [55] [54] Озера, расположенные ближе к экватору, испытывают меньшую сезонность в отношении интенсивности освещения и продолжительности светового дня, чем озера в более высоких широтах (умеренный и полярный пояса). Таким образом, озера в более высоких широтах с большей вероятностью будут испытывать световое ограничение первичной продукции в сезоны низкой освещенности (зима и осень). Сезонные различия температуры в тропиках также не так важны, как для озер более высоких широт. Таким образом, прямое влияние сезонных колебаний температуры на скорость метаболизма более важно в озерах более высоких широт, чем в тропических озерах ( рисунок ). В свою очередь, тропические и субтропические озера с большей вероятностью будут иметь сезонные колебания в зависимости от динамики стратификации и перемешивания, а также режима осадков (влажные и засушливые сезоны), чем из-за четырех астрономических или метеорологических сезонов (весна, лето, осень и зима). по сравнению с озерами более высоких широт.

Сезонные изменения температуры и количества осадков приводят к сезонным изменениям устойчивости водной толщи. В периоды низкой стабильности водного столба более глубокий смешанный слой (полное или частичное перемешивание водного столба, в зависимости от озера) увеличивает поступление питательных веществ и органических веществ из более глубоких слоев и за счет ресуспендирования отложений, что снижает доступность света. И наоборот, в периоды сильной стабильности водного столба внутренняя нагрузка питательных веществ, органических веществ и связанных с ними бактерий в водный столб подавляется, в то время как потеря водорослей из-за опускания увеличивается. Более того, доступность света в этот период выше из-за фотообесцвечивания, меньшего ресуспендирования осадков и меньшей глубины перемешивания, что подвергает фитопланктон воздействию более богатой светом среды. Во многих озерах по всему миру зарегистрированы более высокие показатели ER в период низкой стабильности водного столба как следствие более высокой доступности органического вещества и более высокой биомассы бактерий, связанных с этим органическим веществом. [56] [57] [58] Однако реакция первичной продуктивности на эти сезонные изменения в разных озерах ведет себя по-разному. Как было сказано ранее, реакция скорости метаболизма на эти изменения будет зависеть от ограничивающих факторов первичной продукции в каждом озере ( рисунок ). В периоды низкой стабильности водного столба апвеллинг вод, богатых питательными веществами, может привести к более высоким показателям пелагического GPP, как это наблюдалось в некоторых тропических озерах. [59] [60] И наоборот, в периоды низкой стабильности водного столба показатели GPP могут быть ограничены из-за низкой освещенности, как это наблюдалось в некоторых озерах умеренного и субтропического пояса. [61] [62] Чистый метаболический баланс обычно более отрицателен в периоды дестратификации, даже в озерах, в которых хорошо смешанный сезон является наиболее продуктивным периодом. Несмотря на высокий GPP в этих системах, темпы ER также повышаются за счет увеличения доступности запасов органического вещества из отложений и более глубоких вод.
Сезонные различия в количестве осадков также влияют на скорость метаболизма. Увеличение количества осадков способствует поступлению органических веществ и питательных веществ в озера, что может стимулировать уровень ER и стимулировать или подавлять уровень GPP, в зависимости от баланса между увеличением количества питательных веществ и снижением доступности света. С другой стороны, уменьшение количества осадков влияет и на лимнологические условия, снижая уровень воды и тем самым увеличивая концентрацию питательных веществ и хлорофилла, а также изменяя термическую устойчивость водных сред. Эти изменения могут также повысить ставки ER и GPP. Таким образом, степень реакции скорости метаболизма на сезонные изменения количества осадков будет зависеть от морфометрии озера, свойств водосбора, а также интенсивности и продолжительности выпадения осадков. Озера, часто подвергающиеся сильным штормам, такие как районы тайфунов в северо-западной части Тихого океана, подвергаются интенсивным дождям, которые могут длиться несколько дней. [63] В эти сезоны штормов ожидается снижение скорости обмена веществ из-за уменьшения солнечного света и смывания воды и организмов. Ожидается, что это снижение будет более выраженным в показателях GPP, чем в показателях ER, что приведет к более гетеротрофному NEP (GPP < ER). Например, в субтропическом озере на Тайване во время сезонов тайфунов наблюдалось разделение показателей GPP и ER после сдвига пула органического вещества с автохтонного (органическое вещество, образующееся в озере) на аллохтонное (органическое вещество). добывается за пределами озера). [64] Это говорит о том, что показатели ER были более устойчивы к воздействию тайфуна, чем показатели GPP.
Межгодовые колебания
[ редактировать ]Межгодовая изменчивость скорости метаболизма может быть обусловлена обширными изменениями в водосборе или направленным и циклическим изменением климата и климатическими нарушениями, такими как явления, связанные с южным колебанием Эль-Ниньо (ЭНСО) . Эти изменения в водосборе, температуре воздуха и осадках в разные годы влияют на скорость метаболизма, изменяя поступление питательных и органических веществ в озеро, ослабление света, динамику перемешивания и прямую зависимость метаболических процессов от температуры.
Увеличение количества осадков увеличивает внешнюю нагрузку органических веществ, биогенных веществ и отложений в озерах. Более того, учащение сбросов, вызванное увеличением количества осадков, также может изменить динамику перемешивания и вызвать физическое смывание организмов. При этом уменьшение количества осадков, связанное с высокими скоростями испарения, также влияет на лимнологические условия за счет снижения уровня воды и тем самым увеличения концентрации питательных веществ и хлорофилла, а также изменения термической устойчивости водных сред. В теплые годы более высокая стабильность водного столба ограничивает поступление питательных веществ и органических веществ в фотическую зону. Напротив, в более холодные годы менее стабильный водный столб усиливает повторное взвешивание отложений и поступление питательных веществ и органических веществ из более глубоких вод. Это снижает доступность света, но увеличивает доступность питательных веществ и органических веществ. Таким образом, влияние межгодовых различий осадков и температуры на скорость метаболизма будет зависеть от интенсивности и продолжительности этих изменений, а также от того, какие факторы лимитируют ГПП и ЭР в каждом водоеме.
В озерах с ограничением питательных веществ и органического вещества GPP и ER более влажные годы могут повысить показатели GPP и ER из-за более высокого поступления питательных веществ и органических веществ из ландшафта. Это будет зависеть от того, будут ли наземные ресурсы быстро доступны первичным продуцентам и гетеротрофным сообществам или они попадут в озеро через более глубокие воды, в которых метаболические процессы очень слабы или вообще отсутствуют. В этом случае входные данные будут доступны только в следующем событии смешивания водного столба. Таким образом, увеличение скорости метаболизма за счет осадков зависит также от динамики стратификации и перемешивания, гидрологии и морфометрии озера. С другой стороны, более засушливые годы также могут привести к увеличению показателей GPP и ER, если это сопровождается более низким уровнем воды, что приведет к более высоким концентрациям питательных веществ и органических веществ. Более низкий уровень воды связан с менее стабильной толщей воды и большей близостью к отложениям, что приводит к увеличению поступления питательных и органических веществ из более глубоких вод. Также к эффекту концентрации приводит снижение уровня воды за счет испарения воды. В свою очередь, в теплые годы толща воды более стабильна, а глубина слоя смешения меньше, что снижает внутреннее поступление питательных веществ и органического вещества в смешанный слой. Скорость метаболизма в этом сценарии будет ниже в верхнем смешанном слое. В озерах с В фотической зоне, простирающейся глубже смешанного слоя, скорость метаболизма будет выше на промежуточных глубинах, совпадающих с глубокими максимумами хлорофилла.
В озерах, первичная продукция которых ограничена в основном наличием света, увеличение количества осадков может привести к снижению доступности света, что связано с увеличением растворенного органического вещества и общего количества взвешенных веществ. Следовательно, увеличение количества осадков будет связано с более низкими уровнями GPP, что приведет к снижению скорости дыхания, связанной с автохтонным производством, что приведет к разделению уровней GPP и ER. [65] Кроме того, повышенная доступность аллохтонного органического вещества во влажные годы может привести к более высокому ER и, следовательно, к отрицательному метаболическому балансу (NEP <0). [49]
Изменения годового количества осадков также могут влиять на пространственную изменчивость скорости метаболизма внутри озер. Уильямсон и сотрудники, [49] например, обнаружили, что в гиперэвтрофном водоеме в Северной Америке относительная пространственная изменчивость скоростей GPP и ER была выше в засушливый год по сравнению с влажным. Это предполагает более высокую значимость внутренних процессов, таких как внутренняя нагрузка, поглощение питательных веществ, седиментация и ресуспендирование, для скорости метаболизма в засушливые годы.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Уинслоу, Люк А.; Цварт, Джейкоб А.; Батт, Райан Д.; Дуган, Хилари А.; Вулвей, Р. Истин; Корман, Джессика Р.; Хэнсон, Пол С.; Прочтите, Джордан С. (январь 2016 г.). «LakeMetabolizer: пакет R для оценки метаболизма озера по кислороду свободной воды с использованием различных статистических моделей». Внутренние воды . 6 (4): 622–636. дои : 10.1080/iw-6.4.883 . ISSN 2044-2041 . S2CID 54811593 .
- ^ Хэнсон, Пол С.; Баде, Даррен Л.; Карпентер, Стивен Р.; Крац, Тимоти К. (2003). «Метаболизм озера: взаимосвязь с растворенным органическим углеродом и фосфором» . Лимнология и океанография . 48 (3): 1112–1119. Бибкод : 2003LimOc..48.1112H . дои : 10.4319/lo.2003.48.3.1112 . ISSN 1939-5590 .
- ^ Богард, Мэтью Дж.; Джорджио, Пол А. дель (2016). «Роль метаболизма в модуляции потоков CO 2 в бореальных озерах». Глобальные биогеохимические циклы . 30 (10): 1509–1525. Бибкод : 2016GBioC..30.1509B . дои : 10.1002/2016GB005463 . ISSN 1944-9224 . S2CID 133462828 .
- ^ Уилкинсон, Грейс М.; Буэло, Кэл Д.; Коул, Джонатан Дж.; Пейс, Майкл Л. (2016). «Экзогенно произведенный CO 2 удваивает выброс CO 2 из трех северных озер с умеренным климатом» . Письма о геофизических исследованиях . 43 (5): 1996–2003. Бибкод : 2016GeoRL..43.1996W . дои : 10.1002/2016GL067732 . ISSN 1944-8007 .
- ^ Перейти обратно: а б Ивон-Дюроше, Габриэль; Кэффри, Джейн М.; Ческатти, Алессандро; Доссена, Маттео; Джорджио, Поль дель; Газоль, Хосеп М.; Монтойя, Хосе М.; Пумпанен, Юкка; Штер, Питер А.; Триммер, Марк; Вудворд, Гай (2012). «Согласование температурной зависимости дыхания в разных временных масштабах и типах экосистем» . Природа . 487 (7408): 472–476. Бибкод : 2012Natur.487..472Y . дои : 10.1038/nature11205 . ISSN 1476-4687 . ПМИД 22722862 . S2CID 4422427 .
- ^ Портер, Джон Х.; Хэнсон, Пол С.; Лин, Чау-Чин (01 февраля 2012 г.). «Остаться на плаву в потоке сенсорных данных» . Тенденции в экологии и эволюции . 27 (2): 121–129. дои : 10.1016/j.tree.2011.11.009 . ISSN 0169-5347 . ПМИД 22206661 .
- ^ Коул, Джонатан Дж.; Пейс, Майкл Л.; Карпентер, Стивен Р.; Китчелл, Джеймс Ф. (декабрь 2000 г.). «Сохранение чистой гетеротрофии в озерах во время добавления питательных веществ и манипуляций с пищевой сетью» . Лимнология и океанография . 45 (8): 1718–1730. Бибкод : 2000LimOc..45.1718C . дои : 10.4319/lo.2000.45.8.1718 . ISSN 0024-3590 .
- ^ Хофманн, Хилмар; Энсинас-Фернандес, Хорхе; Тенгберг, Андерс; Атаманчук Дарья; Питерс, Фрэнк (21 декабря 2016 г.). «Метаболизм озера: сравнение скоростей метаболизма в озере, рассчитанных с помощью метода Diel CO 2 и обычного метода Diel O 2 » . ПЛОС ОДИН . 11 (12): e0168393. Бибкод : 2016PLoSO..1168393P . дои : 10.1371/journal.pone.0168393 . ISSN 1932-6203 . ПМК 5176309 . ПМИД 28002477 .
- ^ Дуган, Хилари А.; Вулвей, Р. Истин; Сантосо, Арианто Б.; Корман, Джессика Р.; Джеймс, Алин; Нодин, Эмили Р.; Патил, Виджай П.; Цварт, Джейкоб А.; Брентруп, Дженнифер А.; Хетерингтон, Эми Л.; Оливер, Саманта К. (1 января 2016 г.). «Последствия выбора модели газового потока для интерпретации метаболического баланса в 15 озерах» . Внутренние воды . 6 (4): 581–592. дои : 10.1080/IW-6.4.836 . ISSN 2044-2041 .
- ^ Перейти обратно: а б Садро, Стивен; Холтгрив, Гордон В.; Соломон, Кристофер Т.; Кох, Грегори Р. (2014). «Широко распространенная изменчивость ночных моделей дыхания экосистем связана с градиентами растворенного органического вещества, времени пребывания и продуктивности в глобальном наборе озер» . Лимнология и океанография . 59 (5): 1666–1678. Бибкод : 2014LimOc..59.1666S . дои : 10.4319/lo.2014.59.5.1666 . ISSN 1939-5590 .
- ^ Перейти обратно: а б с д Богерт, Мэтью К. Ван де; Баде, Даррен Л.; Карпентер, Стивен Р.; Коул, Джонатан Дж.; Пейс, Майкл Л.; Хэнсон, Пол С.; Лангман, Оуэн К. (2012). «Пространственная неоднородность сильно влияет на оценки метаболизма экосистем в двух северных озерах с умеренным климатом» . Лимнология и океанография . 57 (6): 1689–1700. Бибкод : 2012LimOc..57.1689V . дои : 10.4319/lo.2012.57.6.1689 . ISSN 1939-5590 .
- ^ Перейти обратно: а б с Обрадор, Биль; Штер, Питер А.; Кристенсен, Джеспер ПК (2014). «Вертикальные закономерности обмена веществ в трех контрастных стратифицированных озерах» . Лимнология и океанография . 59 (4): 1228–1240. Бибкод : 2014LimOc..59.1228O . дои : 10.4319/lo.2014.59.4.1228 . ISSN 1939-5590 .
- ^ Гилинг, Даррен П.; Штер, Питер А.; Гроссарт, Ганс Петер; Андерсен, Миккель Рене; Берер, Бертрам; Эскот, Кармело; Эврендилек, Фатих; Гомес-Генер, Луис; Хонти, Марк; Джонс, Ян Д.; Каракая, Нусрет (2017). «Кунемся глубже: метаболические процессы в металимнионе стратифицированных озер» (PDF) . Лимнология и океанография . 62 (3): 1288–1306. Бибкод : 2017LimOc..62.1288G . дои : 10.1002/lno.10504 . ISSN 1939-5590 . S2CID 56273601 .
- ^ Штер, Питер А.; Бэйд, Даррен; Богерт, Мэтью К. Ван де; Кох, Грегори Р.; Уильямсон, Крейг; Хэнсон, Пол; Коул, Джонатан Дж.; Крац, Тим (2010). «Озерный метаболизм и кислородная техника: состояние науки» . Лимнология и океанография: Методы . 8 (11): 628–644. дои : 10.4319/lom.2010.8.0628 . ISSN 1541-5856 . S2CID 95079460 .
- ^ Бендер, Майкл; Гранде, Карен; Джонсон, Кеннет; Марра, Джон; Уильямс, Питер Дж. ЛеБ.; Зибурт, Джон ; Пилсон, Майкл; Лэнгдон, Крис; Хичкок, Гэри (сентябрь 1987 г.). «Сравнение четырех методов определения продукции планктонного сообщества1». Лимнология и океанография . 32 (5): 1085–1098. Бибкод : 1987LimOc..32.1085B . дои : 10.4319/lo.1987.32.5.1085 . ISSN 0024-3590 .
- ^ Перейти обратно: а б Стецс, Эдвард Г.; Стригль, Роберт Г.; Эйкен, Джордж Р.; Розенберри, Дональд О.; Винтер, Томас К. (2009). «Гидрологическая поддержка потока углекислого газа, выявленная углеродным балансом всего озера» . Журнал геофизических исследований: Биогеонауки . 114 (Г1): G01008. Бибкод : 2009JGRG..114.1008S . дои : 10.1029/2008JG000783 . ISSN 2156-2202 .
- ^ Ловетт, Гэри М.; Коул, Джонатан Дж.; Пейс, Майкл Л. (1 февраля 2006 г.). «Равно ли чистое производство экосистемы накоплению углерода в экосистеме?». Экосистемы . 9 (1): 152–155. дои : 10.1007/s10021-005-0036-3 . ISSN 1435-0629 . S2CID 5890190 .
- ^ Годвин, Шон С.; Джонс, Стюарт Э.; Вейдель, Брайан С.; Соломон, Кристофер Т. (2014). «Концентрация растворенного органического углерода контролирует первичную продукцию бентоса: результаты исследований в камерах in situ в озерах с северной умеренной зоной». Лимнология и океанография . 59 (6): 2112–2120. Бибкод : 2014LimOc..59.2112G . дои : 10.4319/lo.2014.59.6.2112 . ISSN 1939-5590 . S2CID 35841740 .
- ^ Гилинг, Даррен П.; Штер, Питер А.; Гроссарт, Ганс Петер; Андерсен, Миккель Рене; Берер, Бертрам; Эскот, Кармело; Эврендилек, Фатих; Гомес-Генер, Луис; Хонти, Марк; Джонс, Ян Д.; Каракая, Нусрет (2017). «Кунемся глубже: метаболические процессы в металимнионе стратифицированных озер» (PDF) . Лимнология и океанография . 62 (3): 1288–1306. Бибкод : 2017LimOc..62.1288G . дои : 10.1002/lno.10504 . ISSN 1939-5590 . S2CID 56273601 .
- ^ Шарфенбергер, Ульрике; Джеппесен, Эрик; Беклиоглу, Мерьем; Сёндергорд, Мартин; Ангелер, Дэвид Г.; Чакироглу, Айше Идил; Дракаре, Стина; Хейзлар, Йозеф; Махди, Олдуши; Папастергиаду, Ева; Шорф, Михал (2019). «Влияние трофического статуса, уровня воды и температуры на метаболизм и метаболический баланс мелкого озера: стандартизированный общеевропейский эксперимент в мезокосме» . Лимнология и океанография . 64 (2): 616–631. Бибкод : 2019LimOc..64..616S . дои : 10.1002/lno.11064 . hdl : 11511/39476 . ISSN 1939-5590 . S2CID 91853932 .
- ^ «USGS WRI99-4007 Обзор мер по контролю за фосфором в Соединенных Штатах и их влияние на качество воды» . pubs.usgs.gov . Проверено 28 июня 2020 г.
- ^ Когава, Ана Каролина; ЦЕРНИКО, Беатрис Гамберини; ду Коуто, Леандро Джованни Домингуш; Сальгадо, Эрида Регина Нуньес (2017). «Синтетические моющие средства: 100 лет истории» . Саудовский фармацевтический журнал . 25 (6): 934–938. дои : 10.1016/j.jsps.2017.02.006 . ISSN 1319-0164 . ПМЦ 5605839 . ПМИД 28951681 .
- ^ Элзер, Джеймс Дж.; Бракен, Мэтью Э.С.; Клеланд, Эльза Э.; Грюнер, Дэниел С.; Харпол, В. Стэнли; Хиллебранд, Гельмут; Нгаи, Жаклин Т.; Сиблум, Эрик В.; Шурин, Джонатан Б.; Смит, Дженнифер Э. (2007). «Глобальный анализ ограничения азота и фосфора первичными продуцентами в пресноводных, морских и наземных экосистемах». Экологические письма . 10 (12): 1135–1142. Бибкод : 2007EcolL..10.1135E . дои : 10.1111/j.1461-0248.2007.01113.x . hdl : 1903/7447 . ISSN 1461-0248 . ПМИД 17922835 . S2CID 12083235 .
- ^ Личман, Елена ; Штайнер, Дэниел; Боссард, Питер (2003). «Фотосинтетическая и ростовая реакция трех пресноводных водорослей на ограничение фосфора и продолжительность светового дня». Пресноводная биология . 48 (12): 2141–2148. дои : 10.1046/j.1365-2427.2003.01157.x . ISSN 1365-2427 .
- ^ Штер, Питер А.; Бригенти, Людмила С.; Хонти, Марк; Кристенсен, Джеспер; Роуз, Кевин С. (2016). «Глобальные закономерности светового насыщения и фотоингибирования первичной продукции озера». Внутренние воды . 6 (4): 593–607. дои : 10.1080/IW-6.4.888 . ISSN 2044-2041 . S2CID 54945617 .
- ^ Цварт, Джейкоб А.; Соломон, Кристофер Т.; Джонс, Стюарт Э. (2015). «Характеристики фитопланктона предсказывают функционирование экосистемы в глобальном наборе озер». Экология . 96 (8): 2257–2264. дои : 10.1890/14-2102.1 . ISSN 1939-9170 . ПМИД 26405750 .
- ^ Карлссон, Ян; Быстрём, Пяр; Эш, Дженни; Эш, Пер; Перссон, Леннарт; Янссон, Матс (2009). «Световое ограничение бедных питательными веществами озерных экосистем» . Природа . 460 (7254): 506–509. Бибкод : 2009Природа.460..506К . дои : 10.1038/nature08179 . ISSN 1476-4687 . ПМИД 19626113 . S2CID 4360994 .
- ^ Кори, Роуз М.; Уорд, Коллин П.; Крамп, Байрон К.; Клинг, Джордж В. (2014). «Солнечный свет контролирует переработку углерода в толще воды в арктических пресных водах» . Наука . 345 (6199): 925–928. Бибкод : 2014Sci...345..925C . дои : 10.1126/science.1253119 . ISSN 0036-8075 . ПМИД 25146289 . S2CID 206556629 .
- ^ Перейти обратно: а б с Сикелл, Дэвид А.; Лапьер, Жан-Франсуа; Спроси, Дженни; Бергстрем, Анн-Кристин; Дейнингер, Энн; Родригес, Патрисия; Карлссон, Ян (2015). «Влияние растворенного органического углерода на первичную продукцию северных озер» . Лимнология и океанография . 60 (4): 1276–1285. Бибкод : 2015LimOc..60.1276S . дои : 10.1002/lno.10096 . hdl : 11336/5573 . ISSN 1939-5590 . S2CID 84008509 .
- ^ Перейти обратно: а б с Сикелл, Дэвид А.; Лапьер, Жан-Франсуа; Карлссон, Ян (14 июля 2015 г.). «Компромисс между светом и доступностью питательных веществ в зависимости от градиента концентрации растворенного органического углерода в шведских озерах: последствия для моделей первичного производства» . Канадский журнал рыболовства и водных наук . 72 (11): 1663–1671. дои : 10.1139/cjfas-2015-0187 . hdl : 1807/69824 .
- ^ Перейти обратно: а б с д Сикелл, Дэвид А.; Лапьер, Жан-Франсуа; Черувелил, Кендра Спенс (2018). «География круговорота углерода в озерах» . Письма по лимнологии и океанографии . 3 (3): 49–56. дои : 10.1002/lol2.10078 . ISSN 2378-2242 . S2CID 133973800 .
- ^ Келли, Патрик Т.; Соломон, Кристофер Т.; Цварт, Джейкоб А.; Джонс, Стюарт Э. (2018). «Система понимания изменений в пелагической валовой первичной продукции озерных экосистем». Экосистемы . 21 (7): 1364–1376. дои : 10.1007/s10021-018-0226-4 . ISSN 1435-0629 . S2CID 31266760 .
- ^ Бергстрем, Анн-Кристин; Карлссон, Ян (2019). «Свет и питательные вещества контролируют реакцию биомассы фитопланктона на глобальные изменения в северных озерах» . Биология глобальных изменений . 25 (6): 2021–2029. Бибкод : 2019GCBio..25.2021B . дои : 10.1111/gcb.14623 . ISSN 1365-2486 . ПМИД 30897262 . S2CID 85446214 .
- ^ Келли, Патрик Т.; Соломон, Кристофер Т.; Цварт, Джейкоб А.; Джонс, Стюарт Э. (01 ноября 2018 г.). «Система понимания изменений в пелагической валовой первичной продукции озерных экосистем». Экосистемы . 21 (7): 1364–1376. дои : 10.1007/s10021-018-0226-4 . ISSN 1435-0629 . S2CID 31266760 .
- ^ Янссон, Матс; Карлссон, Ян; Йонссон, Андерс (2012). «Перенасыщение углекислым газом способствует первичной продукции в озерах». Экологические письма . 15 (6): 527–532. дои : 10.1111/j.1461-0248.2012.01762.x . ISSN 1461-0248 . ПМИД 22420750 .
- ^ Верспаген, Иоланда М.Х.; Ваал, Дедмер Б. Ван де; Финке, Ян Ф.; Виссер, Петра М.; Донк, Эллен Ван; Хейсман, Джефф (13 августа 2014 г.). «Повышение уровня CO 2 усилит цветение фитопланктона в эвтрофных и гипертрофных озерах» . ПЛОС ОДИН . 9 (8): е104325. Бибкод : 2014PLoSO...9j4325V . дои : 10.1371/journal.pone.0104325 . ISSN 1932-6203 . ПМК 4132121 . ПМИД 25119996 .
- ^ Танк, Сюзанна Э.; Лесак, Лэнс Ф.В.; МакКуин, Дональд Дж. (2009). «Повышенный pH регулирует бактериальный круговорот углерода в озерах с высокой фотосинтетической активностью». Экология . 90 (7): 1910–1922. дои : 10.1890/08-1010.1 . ISSN 1939-9170 . ПМИД 19694139 .
- ^ Кристенсен, Джеспер, Пенсильвания; Санд-Йенсен, Кай; Штер, Питер А. (2013). «Колебания уровня воды контролируют химический состав воды и метаболизм пруда, в котором преобладают харофиты». Пресноводная биология . 58 (7): 1353–1365. дои : 10.1111/fwb.12132 . ISSN 1365-2427 .
- ^ Конт, Жером; Лангенхедер, Силке; Берга, Мерсе; Линдстрем, Ева С. (2017). «Вклад различных источников распространения в метаболическую реакцию бактериопланктона озера после изменения солености». Экологическая микробиология . 19 (1): 251–260. дои : 10.1111/1462-2920.13593 . ISSN 1462-2920 . ПМИД 27871136 . S2CID 13011521 .
- ^ Цзян, Хунчэнь, Хайлян; Лю, Синьци; Ли, Цзи, Шаньшань; Чжан, Чуанлунь Л. (2007). . микробиология : 9 : 2603–2621 doi Экологическая 10.1111 . ISSN 1462-2920 . / j.1462-2920.2007.01377.x (10 )
- ^ Топфер, Конрад; Бартон, Майкл (1 сентября 1992 г.). «Влияние солености на скорость потребления кислорода у двух видов пресноводных рыб: Phoxinus erythrogaster (семейство Cyprinidae) и Fundulus catenatus (семейство Fundulidae)». Гидробиология . 242 (3): 149–154. дои : 10.1007/BF00019963 . ISSN 1573-5117 . S2CID 21293418 .
- ^ Альфонсо, Мария Вифлеем; Брендель, Андреа Соледад; Витале, Алехандро Хосе; Зейтц, Карина; Пикколо, Мария Синтия; Перилло, Херардо Мигель Эдуардо (2018). «Драйверы экосистемного метаболизма в двух управляемых мелких озерах с разной соленостью и трофическими условиями: озера Соус-Гранде и Ла-Салада (Аргентина)» . Вода . 10 (9): 1136. дои : 10.3390/w10091136 . hdl : 11336/90492 .
- ^ Морейра-Турк, Патрисия Ф. (1 июня 2000 г.). «Влияние года с низкой соленостью на метаболизм гиперсоленой прибрежной лагуны (Бразилия)». Гидробиология . 429 (1): 133–140. дои : 10.1023/A:1004037624787 . ISSN 1573-5117 . S2CID 20968144 .
- ^ Дуган, Хилари А.; Бартлетт, Сара Л.; Берк, Саманта М.; Дубек, Джонатан П.; Кривак-Тетли, Флора Э.; Скафф, Николас К.; Саммерс, Джейми С.; Фаррелл, Кейтлин Дж.; Маккалоу, Ян М.; Моралес-Уильямс, Ана М.; Робертс, Дерек К. (25 апреля 2017 г.). «Соление наших пресноводных озер» . Труды Национальной академии наук . 114 (17): 4453–4458. Бибкод : 2017PNAS..114.4453D . дои : 10.1073/pnas.1620211114 . ISSN 0027-8424 . ПМК 5410852 . ПМИД 28396392 .
- ^ Корси, Стивен Р.; Грачик, Дэвид Дж.; Гейс, Стивен В.; Бут, Натаниэль Л.; Ричардс, Кевин Д. (1 октября 2010 г.). «Свежий взгляд на дорожную соль: водная токсичность и влияние на качество воды в местном, региональном и национальном масштабах» . Экологические науки и технологии . 44 (19): 7376–7382. Бибкод : 2010EnST...44.7376C . дои : 10.1021/es101333u . ISSN 0013-936X . ПМЦ 2947309 . ПМИД 20806974 .
- ^ Перейти обратно: а б Колозо, Джеймс Дж; Коул, Джонатан Дж; Хэнсон, Пол С; Пейс, Майкл Л. (2008). «Интегрированные по глубине, непрерывные оценки метаболизма в озере с чистой водой». Канадский журнал рыболовства и водных наук . 65 (4): 712–722. дои : 10.1139/f08-006 . ISSN 0706-652X .
- ^ Кавальканти, Дж. Рафаэль; да Мотта-Маркес, Дэвид; Фрагосо, Карлос Руберто (2016). «Процессуальное моделирование метаболизма мелководных озер: пространственно-временная изменчивость и относительная важность отдельных процессов» . Экологическое моделирование . 323 : 28–40. doi : 10.1016/j.ecolmodel.2015.11.010 . ISSN 0304-3800 .
- ^ Джонс, Стюарт Э.; Цварт, Джейкоб А.; Келли, Патрик Т.; Соломон, Кристофер Т. (2018). «Гидрологическая обстановка ограничивает гетеротрофию озер и судьбу наземного углерода» . Письма по лимнологии и океанографии . 3 (3): 256–264. дои : 10.1002/lol2.10054 . ISSN 2378-2242 .
- ^ Перейти обратно: а б с Уильямсон, Таннер Дж.; Ванни, Майкл Дж.; Ренвик, Уильям Х. (2020). «Пространственная и временная изменчивость динамики питательных веществ и метаболизма экосистемы в гиперэвтрофном водоеме различается в влажный и засушливый год». Экосистемы . 24 : 68–88. дои : 10.1007/s10021-020-00505-8 . ISSN 1435-0629 . S2CID 216559711 .
- ^ Ивон-Дюроше, Габриэль; Кэффри, Джейн М.; Ческатти, Алессандро; Доссена, Маттео; Джорджио, Поль дель; Газоль, Хосеп М.; Монтойя, Хосе М.; Пумпанен, Юкка; Штер, Питер А.; Триммер, Марк; Вудворд, Гай (2012). «Согласование температурной зависимости дыхания в разных временных масштабах и типах экосистем» . Природа . 487 (7408): 472–476. Бибкод : 2012Natur.487..472Y . дои : 10.1038/nature11205 . ISSN 1476-4687 . ПМИД 22722862 . S2CID 4422427 .
- ^ Роуз, Кевин С.; Уинслоу, Люк А.; Прочтите, Джордан С.; Прочтите, Эмили К.; Соломон, Кристофер Т.; Адриан, Рита; Хэнсон, Пол К. (2014). «Повышение точности оценок метаболизма экосистемы озера путем выявления предикторов неопределенности модели» . Лимнология и океанография: Методы . 12 (5): 303–312. дои : 10.4319/лом.2014.12.303 . ISSN 1541-5856 . S2CID 96712639 .
- ^ Ламперт, Винфрид; Флекнер, Уолтер; Рай, Хакумат; Тейлор, Барбара Э. (1986). «Контроль фитопланктона путем выпаса зоопланктона: исследование весенней фазы прозрачной воды1» . Лимнология и океанография . 31 (3): 478–490. Бибкод : 1986LimOc..31..478L . дои : 10.4319/lo.1986.31.3.0478 . ISSN 1939-5590 .
- ^ Перейти обратно: а б Обертеггер, Ульрике; Обрадор, Биль; Флаим, Джованна (2017). «Динамика растворенного кислорода подо льдом: три зимы высокочастотных данных из озера Товель, Италия». Исследования водных ресурсов . 53 (8): 7234–7246. Бибкод : 2017WRR....53.7234O . дои : 10.1002/2017WR020599 . hdl : 2445/123119 . ISSN 1944-7973 . S2CID 55378645 .
- ^ Перейти обратно: а б с Сун, Чанъю; Чжао, Тянь, Вэйдун; Цао, Сяовэй; Ван, Хуотари, Юсси (2019). Подледный " . мелком в и климате в холодном метаболизм засушливом озере
- ^ Блок, Бенджамин Д.; Денфельд, Блейз А.; Стоквелл, Джейсон Д.; Флайм, Джованна; Гроссарт, Ханс-Петер Ф.; Нолл, Лесли Б.; Майер, Доминик Б.; Норт, Ребекка Л.; Раутио, Милла; Русак, Джеймс А.; Садро, Стив (2019). «Уникальные методологические проблемы зимней лимнологии» . Лимнология и океанография: Методы . 17 (1): 42–57. дои : 10.1002/lom3.10295 . ISSN 1541-5856 .
- ^ Бидданда, Бопайя А.; Котнер, Джеймс Б. (2002). «Ручки любви в водных экосистемах: роль снижения растворенного органического углерода, ресуспендированных отложений и терригенных примесей в углеродном балансе озера Мичиган». Экосистемы . 5 (5): 431–445. дои : 10.1007/s10021-002-0163-z . ISSN 1432-9840 . S2CID 25449277 .
- ^ Перейти обратно: а б Штер, Питер А.; Санд-Йенсен, Кай (январь 2007 г.). «Временная динамика и регуляция озерного метаболизма» . Лимнология и океанография . 52 (1): 108–120. Бибкод : 2007LimOc..52..108S . дои : 10.4319/lo.2007.52.1.0108 . ISSN 1939-5590 .
- ^ Перейти обратно: а б Бригенти, Людмила Силва; Штер, Питер Антон; Гальярди, Лаура Мартинс; БРАНДИО, Лусиана Пенья Мелло; Элиас, Элиан Корреа; де Меллу, Нельсон Азеведу Сантос Тейшейра; БАРБОСА, Франсиско Антониу Родригес; Безерра-Нето, Хосе Фернандес (июнь 2015 г.). «Временные изменения скорости метаболизма двух тропических озер в атлантическом лесу Бразилии» . Экосистемы . 18 (4): 589–604. дои : 10.1007/s10021-015-9851-3 . ISSN 1435-0629 . S2CID 16135518 .
- ^ Бригенти, Людмила С.; Штер, Питер А.; Брандао, Лусиана, премьер-министр; Барбоса, Франциско, Арканзас; Безерра-Нето, Хосе Фернандес (2018). «Важность питательных веществ, органических веществ и доступности света для скорости эпилимнетического метаболизма в мезотрофном тропическом озере». Пресноводная биология . 63 (9): 1143–1160. дои : 10.1111/fwb.13123 . ISSN 1365-2427 .
- ^ Гальярди, Лаура Мартинс; Бригенти, Людмила Силва; Штер, Питер Антон; Барбоза, Франсиско Антониу Родригес; Безерра-Нето, Хосе Фернандес (2019). «Уменьшение количества осадков увеличивает скорость метаболизма в верхних смешанных слоях тропических озер». Экосистемы . 22 (6): 1406–1423. дои : 10.1007/s10021-019-00346-0 . ISSN 1435-0629 . S2CID 71146859 .
- ^ Штер, Питер А.; Санд-Йенсен, Кай; Раун, Ане Л.; Нильссон, Бертель; Кидмос, Джейкоб (2010). «Драйверы метаболизма и чистая гетеротрофия в контрастных озерах» . Лимнология и океанография . 55 (2): 817–830. Бибкод : 2010LimOc..55..817S . дои : 10.4319/lo.2010.55.2.0817 . ISSN 1939-5590 .
- ^ Тонетта, Дениз; Штер, Питер Антон; Шмитт, Рафаэль; Петручио, Маурисио Мелло (2016). «Физические условия, обуславливающие пространственную и временную изменчивость водного метаболизма субтропического прибрежного озера» . Лимнологика . 58 : 30–40. дои : 10.1016/j.limno.2016.01.006 . ISSN 0075-9511 .
- ^ Джонс, Стюарт Э.; Крац, Тимоти К.; Чиу, Чи-Ю; МакМАХОН, Кэтрин Д. (2009). «Влияние тайфунов на годовой поток CO 2 из субтропического гуминового озера». Биология глобальных изменений . 15 (1): 243–254. Бибкод : 2009GCBio..15..243J . дои : 10.1111/j.1365-2486.2008.01723.x . ISSN 1365-2486 . S2CID 20671659 .
- ^ Цай, Дженг-Вэй; Крац, Тимоти К.; Хэнсон, Пол С.; Кимура, Нобуаки; Лю, Вэнь-Чэн; Линь, Фанг-Пан; Чжоу, Сю-Мэй; Ву, Цзюнь-Цонг; Чиу, Чи-Ю (2011). «Метаболические изменения, устойчивость и устойчивость субтропического гетеротрофного озера к возмущениям тайфунов». Канадский журнал рыболовства и водных наук . 68 (5): 768–780. дои : 10.1139/f2011-024 . ISSN 0706-652X .
- ^ Ричардсон, Дэвид С.; Кэри, Кайелан К.; Брюсевиц, Дениз А.; Уэзерс, Кэтлин С. (2017). «Внутри- и межгодовая изменчивость метаболизма в олиготрофном озере» . Водные науки . 79 (2): 319–333. дои : 10.1007/s00027-016-0499-7 . hdl : 10919/94382 . ISSN 1420-9055 .