Скорость мутации
В генетике частота мутаций — это частота новых мутаций в одном гене, нуклеотидной последовательности или организме с течением времени. [2] Скорость мутаций не является постоянной и не ограничивается одним типом мутации; существует много различных типов мутаций. Частота мутаций указана для конкретных классов мутаций. Точечные мутации — это класс мутаций, которые представляют собой изменения одного основания. Миссенс- и нонсенс-мутации представляют собой два подтипа точковых мутаций. Скорость этих типов замен можно дополнительно разделить на спектр мутаций, который описывает влияние генетического контекста на скорость мутаций. [3]
Для каждой из этих скоростей существует несколько естественных единиц времени, при этом темпы характеризуются либо мутациями на пару оснований на деление клетки, на ген на поколение или на геном на поколение. Скорость мутаций организма является эволюционной характеристикой и находится под сильным влиянием генетики каждого организма, а также сильного влияния окружающей среды. Верхний и нижний пределы, до которых может развиваться частота мутаций, являются предметом продолжающихся исследований. Однако частота мутаций варьируется в зависимости от генома . Скорость мутаций в ДНК, РНК или отдельном гене меняется. [ нужна ссылка ]
Когда уровень мутаций у людей увеличивается, могут возникнуть определенные риски для здоровья, например, рак и другие наследственные заболевания. Знание скорости мутаций жизненно важно для понимания будущего рака и многих наследственных заболеваний. [4]
Фон
[ редактировать ]Различные генетические варианты внутри вида называются аллелями, поэтому новая мутация может создать новую аллель. В популяционной генетике каждая аллель характеризуется коэффициентом отбора, который измеряет ожидаемое изменение частоты аллеля с течением времени. Коэффициент отбора может быть либо отрицательным, что соответствует ожидаемому уменьшению, либо положительным, что соответствует ожидаемому увеличению, либо нулевым, что соответствует отсутствию ожидаемых изменений. Распределение эффектов приспособленности новых мутаций является важным параметром популяционной генетики и стало предметом обширных исследований. [5] Хотя измерения этого распределения в прошлом были противоречивыми, сейчас принято считать, что большинство мутаций умеренно вредны, что многие из них мало влияют на приспособленность организма, а некоторые могут быть благоприятными.
В результате естественного отбора неблагоприятные мутации обычно удаляются из популяции, в то время как благоприятные изменения обычно сохраняются для следующего поколения, а нейтральные изменения накапливаются со скоростью, с которой они создаются мутациями. Этот процесс происходит путем размножения. В конкретном поколении «наиболее приспособленные» выживают с большей вероятностью, передавая свои гены потомству. Знак изменения этой вероятности определяет мутации как полезные, нейтральные или вредные для организмов. [6]
Измерение
[ редактировать ]Скорость мутаций в организме можно измерить с помощью ряда методов.
Одним из способов измерения частоты мутаций является тест флуктуации, также известный как эксперимент Лурии-Дельбрюка . Этот эксперимент продемонстрировал, что мутации бактерий происходят при отсутствии отбора, а не при наличии отбора. [7]
Это очень важно для скорости мутаций, поскольку экспериментально доказывает, что мутации могут происходить без участия отбора — на самом деле мутация и отбор представляют собой совершенно разные эволюционные силы . Различные последовательности ДНК могут иметь разную склонность к мутациям (см. ниже) и не могут происходить случайным образом. [8]
Наиболее часто измеряемым классом мутаций являются замены, поскольку их относительно легко измерить с помощью стандартного анализа данных последовательностей ДНК. Однако замены имеют существенно разную скорость мутаций (10 −8 до 10 −9 на поколение для большинства клеточных организмов), чем мутации других классов, которые зачастую намного выше (~10 −3 на поколение для расширения/сокращения сателлитной ДНК [9] ).
Коэффициенты замещения
[ редактировать ]Многие участки генома организма могут допускать мутации с небольшим эффектом приспособленности. Такие сайты обычно называются нейтральными. Теоретически мутации без отбора фиксируются между организмами именно с той скоростью мутации. Фиксированные синонимичные мутации, то есть синонимичные замены , представляют собой изменения последовательности гена, которые не меняют белок, продуцируемый этим геном. Их часто используют для оценки частоты мутаций, несмотря на то, что некоторые синонимичные мутации оказывают влияние на приспособленность. Например, частота мутаций была напрямую выведена из последовательностей всего генома экспериментально полученных репликативных линий Escherichia coli B. [10]
Линии накопления мутаций
[ редактировать ]Особенно трудоемким способом характеристики скорости мутаций является линия накопления мутаций.
Линии накопления мутаций использовались для характеристики скорости мутаций с помощью метода Бейтмана-Мукаи и прямого секвенирования хорошо изученных экспериментальных организмов, начиная от кишечных бактерий ( E. coli ), круглых червей ( C. elegans ), дрожжей ( S. cerevisiae ), фруктов. мухи ( D. melanogaster ) и мелкие эфемерные растения ( A. thaliana ). [11]
Изменение скорости мутаций
[ редактировать ]
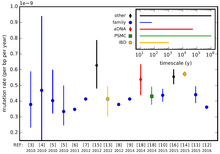
Скорость мутаций различается между видами и даже между разными областями генома одного вида. Эти различные скорости замен нуклеотидов измеряются в заменах ( фиксированных мутациях ) на пару оснований за поколение. Например, мутации в межгенной, или некодирующей, ДНК имеют тенденцию накапливаться быстрее, чем мутации в ДНК, которая активно используется в организме ( экспрессия генов ). Это не обязательно связано с более высокой частотой мутаций, а с более низким уровнем очищающего отбора . Область, которая мутирует с предсказуемой скоростью, является кандидатом на использование в качестве молекулярных часов .
Если скорость нейтральных мутаций в последовательности предполагается постоянной (подобной часам) и если большинство различий между видами нейтральны, а не адаптивны, то количество различий между двумя разными видами можно использовать для оценки того, как давно два вида виды разошлись (см. молекулярные часы ). Фактически, скорость мутаций организма может меняться в ответ на стресс окружающей среды. Например, ультрафиолетовый свет повреждает ДНК, что может привести к ошибкам в попытках клетки выполнить восстановление ДНК .
Частота мутаций у человека выше в мужской зародышевой линии (сперме), чем в женской (яйцеклетке), но оценки точной скорости различаются на порядок или более. Это означает, что в геноме человека накапливается около 64 новых мутаций за поколение, поскольку каждое полное поколение включает в себя ряд клеточных делений для образования гамет. [12] По оценкам, митохондриальная ДНК человека имеет частоту мутаций ~3× или ~2,7×10. −5 за базу за 20-летнюю генерацию (в зависимости от метода оценки); [13] эти показатели считаются значительно выше, чем показатели геномных мутаций человека, примерно в 2,5×10. −8 на базу на поколение. [14] Используя данные, доступные в результате полногеномного секвенирования, частота мутаций в геноме человека оценивается примерно в 1,1×10. −8 на сайт на поколение. [15]
Частота других форм мутаций также сильно отличается от точковых мутаций . Отдельный микросателлитный локус часто имеет частоту мутаций порядка 10. −4 , хотя это может сильно отличаться в зависимости от длины. [16]
Некоторые последовательности ДНК могут быть более восприимчивы к мутациям. Например, участки ДНК в человеческой сперме, в которых отсутствует метилирование, более склонны к мутациям. [17]
В целом частота мутаций у одноклеточных эукариот (и бактерий ) составляет примерно 0,003 мутации на геном на одно поколение клеток . [12] Однако некоторые виды, особенно инфузории рода Paramecium, имеют необычно низкую частоту мутаций. Например, у Paramecium тетраурелия частота мутаций с заменой оснований составляет ~ 2 × 10. −11 на сайт на деление клетки. Это самая низкая частота мутаций, наблюдаемая на данный момент в природе: она примерно в 75 раз ниже, чем у других эукариот с аналогичным размером генома, и даже в 10 раз ниже, чем у большинства прокариот. Низкая частота мутаций у Paramecium объясняется его транскрипционно молчащим зародышевой линии ядром , что согласуется с гипотезой о том, что точность репликации выше при более низких уровнях экспрессии генов . [18]
Самая высокая частота мутаций на пару оснований на поколение наблюдается у вирусов , которые могут иметь геномы РНК или ДНК. ДНК-вирусы имеют частоту мутаций от 10 −6 до 10 −8 мутаций на одно основание на поколение, а РНК-вирусы имеют частоту мутаций от 10 −3 до 10 −5 на базу на поколение. [12]
Спектр мутаций
[ редактировать ]Спектр мутаций — это распределение скоростей или частот мутаций, значимых в определенном контексте, основанное на признании того, что уровни возникновения не все одинаковы. В любом контексте спектр мутаций отражает детали мутагенеза и зависит от таких условий, как наличие химических мутагенов или генетического фона с мутаторными аллелями или поврежденных систем репарации ДНК. Самая фундаментальная и обширная концепция спектра мутаций — это распределение скоростей всех отдельных мутаций, которые могут произойти в геноме (например, [19] ). Например, из этого полного спектра de novo можно рассчитать относительную скорость мутаций в кодирующих и некодирующих регионах. Обычно концепция спектра частот мутаций упрощается, чтобы охватить широкие классы, такие как переходы и трансверсии (рисунок), т.е. различные мутационные преобразования по всему геному объединяются в классы, и для каждого класса существует совокупная частота.
Во многих контекстах спектр мутаций определяется как наблюдаемая частота мутаций, идентифицируемых по некоторому критерию отбора, например, распределение мутаций, клинически связанных с конкретным типом рака, [20] или распределение адаптивных изменений в конкретном контексте, таких как устойчивость к антибиотикам (например, [21] ).В то время как спектр частот мутаций de novo отражает только мутагенез, этот вид спектра может также отражать эффекты систематических ошибок отбора и установления (например, оба вида спектра используются в [22] ).

Эволюция
[ редактировать ]Теория эволюции частоты мутаций определяет три основные задействованные силы: генерацию более вредных мутаций с более высокой мутацией, генерацию более выгодных мутаций с более высокой мутацией, а также метаболические затраты и снижение скорости репликации, необходимые для предотвращения мутаций. Различные выводы делаются в зависимости от относительной важности, приписываемой каждой силе. Оптимальная скорость мутаций организмов может быть определена путем компромисса между затратами на высокую скорость мутаций, [23] такие как вредные мутации и метаболические затраты на поддержание систем для снижения скорости мутаций (например, увеличение экспрессии ферментов репарации ДНК). [24] или, согласно обзору Bernstein et al. [25] с повышенным потреблением энергии для восстановления, кодированием дополнительных генных продуктов и/или с более медленной репликацией). Во-вторых, более высокая частота мутаций увеличивает частоту полезных мутаций, и эволюция может предотвратить снижение частоты мутаций, чтобы поддерживать оптимальные темпы адаптации. [26] Таким образом, гипермутация позволяет некоторым клеткам быстро адаптироваться к изменяющимся условиям, чтобы избежать вымирания всей популяции. [27] Наконец, естественный отбор может не суметь оптимизировать частоту мутаций из-за относительно незначительных преимуществ снижения частоты мутаций, и, таким образом, наблюдаемая частота мутаций является продуктом нейтральных процессов. [28] [29]
Исследования показали, что лечение РНК-вирусов, таких как полиовирус, рибавирином дает результаты, соответствующие идее о том, что вирусы слишком часто мутируют, чтобы поддерживать целостность информации в своих геномах. [30] Это называется катастрофой ошибок .
Высокая частота мутаций ВИЧ (вирус иммунодефицита человека) - 3 x 10. −5 на одно основание и поколение в сочетании с коротким циклом репликации приводит к высокой вариабельности антигена , что позволяет ему уклоняться от иммунной системы. [31]
См. также
[ редактировать ]- Мутация
- Критическая частота мутаций
- Частота мутаций
- Дисгеника
- Частота аллеля
- Скорость эволюции
- Генетика
- Рак
Ссылки
[ редактировать ]- ^ Скалли А (декабрь 2016 г.). «Скорость мутаций в эволюции человека и демографические выводы» . Текущее мнение в области генетики и развития . 41 : 36–43. дои : 10.1016/j.gde.2016.07.008 . ПМИД 27589081 . Архивировано из оригинала 02 января 2021 г. Проверено 8 сентября 2020 г.
- ^ Кроу Дж. Ф. (август 1997 г.). «Высокий уровень спонтанных мутаций: риск для здоровья?» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (16): 8380–6. Бибкод : 1997PNAS...94.8380C . дои : 10.1073/pnas.94.16.8380 . ПМК 33757 . ПМИД 9237985 .
- ^ Поуп К.Ф., О'Салливан Д.М., МакХью Т.Д., Гиллеспи С.Х. (апрель 2008 г.). «Практическое руководство по измерению уровня мутаций при устойчивости к антибиотикам» . Антимикробные средства и химиотерапия . 52 (4): 1209–14. дои : 10.1128/AAC.01152-07 . ПМК 2292516 . ПМИД 18250188 .
- ^ Томлинсон И.П., Новелли М.Р., Бодмер В.Ф. (декабрь 1996 г.). «Скорость мутаций и рак» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (25): 14800–3. Бибкод : 1996PNAS...9314800T . дои : 10.1073/pnas.93.25.14800 . ПМК 26216 . ПМИД 8962135 .
- ^ Эйр-Уокер А., Кейтли П.Д. (август 2007 г.). «Распределение фитнес-эффектов новых мутаций». Обзоры природы. Генетика . 8 (8): 610–8. дои : 10.1038/nrg2146 . ПМИД 17637733 . S2CID 10868777 .
- ^ Скалли А., Дурбин Р. (октябрь 2012 г.). «Пересмотр скорости мутаций человека: значение для понимания эволюции человека». Обзоры природы. Генетика . 13 (10): 745–53. дои : 10.1038/nrg3295 . ПМИД 22965354 . S2CID 18944814 .
- ^ "Эксперимент Лурии-Дельбрюка" . Arc.Ask3.Ru . 25 апреля 2017 г.
- ^ Монро, Дж. Г., Шрикант, Т., Карбонелл-Беджерано, П. и др. Смещение мутаций отражает естественный отбор Arabidopsis thaliana. Природа 602, 101–105 (2022). https://doi.org/10.1038/s41586-021-04269-6. Архивировано 25 июня 2022 г. в Wayback Machine.
- ^ Флинн, Жюльен М.; Калдас, Ян; Кристеску, Мелания Э.; Кларк, Эндрю Г. (2017). «Отбор ограничивает высокие уровни тандемных повторяющихся мутаций ДНК у Daphnia pulex» . Генетика . 207 (2): 697–710. дои : 10.1534/genetics.117.300146 . ПМЦ 5629333 . ПМИД 28811387 . Архивировано из оригинала 21 марта 2020 г. Проверено 21 марта 2020 г.
- ^ Вилгосс С., Баррик Дж.Э., Тенайлон О., Крювейлер С., Чейн-Вун-Минг Б., Медиг С., Ленски Р.Э., Шнайдер Д. (август 2011 г.). «Скорость мутаций, выведенная на основе синонимических замен в долгосрочном эксперименте по эволюции Escherichia coli» . Г3 . 1 (3): 183–186. дои : 10.1534/g3.111.000406 . ПМК 3246271 . ПМИД 22207905 .
- ^ Оссовский С., Шнебергер К., Лукас-Лледо Дж.И., Вартманн Н., Кларк Р.М., Шоу Р.Г., Вайгель Д., Линч М. (январь 2010 г.). «Скорость и молекулярный спектр спонтанных мутаций Arabidopsis thaliana» . Наука . 327 (5961): 92–4. Бибкод : 2010Sci...327...92O . дои : 10.1126/science.1180677 . ПМЦ 3878865 . ПМИД 20044577 .
- ^ Jump up to: а б с Дрейк Дж.В., Чарльзуорт Б., Чарльзуорт Д., Кроу Дж.Ф. (апрель 1998 г.). «Темпы спонтанных мутаций» . Генетика . 148 (4): 1667–86. дои : 10.1093/генетика/148.4.1667 . ПМК 1460098 . ПМИД 9560386 . Архивировано из оригинала 21 августа 2010 г. Проверено 27 сентября 2007 г.
- ^ Шнайдер С., Экскофье Л. (июль 1999 г.). «Оценка прошлых демографических параметров по распределению парных различий, когда скорость мутаций варьируется в зависимости от сайта: применение к митохондриальной ДНК человека» . Генетика . 152 (3): 1079–89. дои : 10.1093/генетика/152.3.1079 . ПМК 1460660 . ПМИД 10388826 . Архивировано из оригинала 8 сентября 2008 г. Проверено 25 февраля 2008 г.
- ^ Нахман М.В., Кроуэлл С.Л. (сентябрь 2000 г.). «Оценка частоты мутаций на нуклеотид у человека» . Генетика . 156 (1): 297–304. дои : 10.1093/генетика/156.1.297 . ПМЦ 1461236 . ПМИД 10978293 . Архивировано из оригинала 8 апреля 2011 г. Проверено 19 октября 2007 г.
- ^ Роуч Дж.С., Глусман Г., Смит А.Ф., Хафф CD, Хабли Р., Шеннон П.Т., Роуэн Л., Пант К.П., Гудман Н., Бамшад М., Шендюр Дж., Дрманак Р., Джорд Л.Б., Худ Л., Галас DJ (апрель 2010 г.). «Анализ генетического наследования в семейном квартете методом полногеномного секвенирования» . Наука . 328 (5978): 636–9. Бибкод : 2010Sci...328..636R . дои : 10.1126/science.1186802 . ПМК 3037280 . ПМИД 20220176 .
- ^ Уиттакер Дж.К., Харборд Р.М., Боксалл Н., Маккей И., Доусон Дж., Сибли Р.М. (июнь 2003 г.). «Оценка частоты микросателлитных мутаций на основе вероятности» . Генетика . 164 (2): 781–7. дои : 10.1093/генетика/164.2.781 . ПМЦ 1462577 . ПМИД 12807796 . Архивировано из оригинала 28 ноября 2011 г. Проверено 3 мая 2011 г.
- ^ Гравтиз, Лорен (28 июня 2012 г.). «Отсутствие модификации ДНК создает горячие точки для мутаций» . Инициатива по исследованию аутизма Фонда Саймонса. Архивировано из оригинала 5 марта 2014 года . Проверено 20 февраля 2014 г.
- ^ Сунг В., Такер А.Е., Доак Т.Г., Чой Э., Томас В.К., Линч М. (ноябрь 2012 г.). «Чрезвычайная стабильность генома инфузории Paramecium Tetraurelia» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (47): 19339–44. Бибкод : 2012PNAS..10919339S . дои : 10.1073/pnas.1210663109 . ПМЦ 3511141 . ПМИД 23129619 .
- ^ Э. Лопес-Кортегано, Р. Дж. Крейг, Дж. Чебиб, Э. Дж. Балогун и П. Д. Кейтли (2022). «Скорость и спектры структурных мутаций de novo у Chlamydomonas Reinhardtii» . Геном Рез . 33 (1): 45–60. дои : 10.1101/гр.276957.122 . ПМЦ 9977147 . ПМИД 36617667 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ В. Цао, С. Ван и Дж. К. Ли (2013). «Наследственный рак молочной железы у ханьцев» . Дж Эпидемиол . 23 (2): 75–84. дои : 10.2188/jea.je20120043 . ПМК 3700245 . ПМИД 23318652 .
- ^ Д.Д. Леви, Б. Шарма и Т.А. Себула (2004). «Спектры мутаций однонуклеотидного полиморфизма и устойчивость к хинолонам у серовара Enteritidis Salmonella enterica с мутаторным фенотипом» . Противомикробные средства Химиотер . 48 (7): 2355–63. doi : 10.1128/AAC.48.7.2355-2363.2004 . ПМК 434170 . ПМИД 15215081 .
- ^ А.В. Кано, Х. Рожонова, А. Штольцфус, Д.М. МакКэндлиш и Дж.Л. Пейн (2022). «Смещение мутаций формирует спектр адаптивных замен» . Proc Natl Acad Sci США . 119 (7). Бибкод : 2022PNAS..11919720C . дои : 10.1073/pnas.2119720119 . ПМЦ 8851560 . ПМИД 35145034 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Альтенберг Л. (июнь 2011 г.). «Принцип эволюционного снижения скорости мутаций в нескольких локусах». Бюллетень математической биологии . 73 (6): 1227–70. arXiv : 0909.2454 . дои : 10.1007/s11538-010-9557-9 . ПМИД 20737227 . S2CID 15027684 .
- ^ Сниговский П.Д., Джерриш П.Дж., Джонсон Т., Шейвер А. (декабрь 2000 г.). «Эволюция скорости мутаций: отделение причин от последствий». Биоэссе . 22 (12): 1057–66. doi : 10.1002/1521-1878(200012)22:12<1057::AID-BIES3>3.0.CO;2-W . ПМИД 11084621 . S2CID 36771934 .
- ^ Бернштейн Х., Хопф Ф.А., Мишод Р.Э. (1987). «Молекулярные основы эволюции пола». Молекулярная генетика развития . Достижения генетики. Том. 24. С. 323–70. дои : 10.1016/s0065-2660(08)60012-7 . ISBN 9780120176243 . ПМИД 3324702 .
- ^ Орр Х.А. (июнь 2000 г.). «Скорость адаптации у асексуалов» . Генетика . 155 (2): 961–8. дои : 10.1093/генетика/155.2.961 . ПМК 1461099 . ПМИД 10835413 . Архивировано из оригинала 25 июня 2022 г. Проверено 1 ноября 2014 г.
- ^ Качели, Шоу; Ван ден Берг, Брэм; Вуйтс, Сандер; Ойен, Элин; Вордекерс, Карин; Верстрепен, Кевин Дж; Фоварт, Мартен; Верстратен, Натали; Михилс, Ян (2 мая 2017 г.). «Адаптивная настройка скорости мутаций позволяет быстро реагировать на летальный стресс у Escherichia coli» . электронная жизнь . 6 . doi : 10.7554/eLife.22939 . ISSN 2050-084X . ПМК 5429094 . ПМИД 28460660 .
- ^ Линч М. (август 2010 г.). «Эволюция скорости мутаций» . Тенденции в генетике . 26 (8): 345–52. дои : 10.1016/j.tig.2010.05.003 . ПМЦ 2910838 . ПМИД 20594608 .
- ^ Сунг В., Акерман М.С., Миллер С.Ф., Доак Т.Г., Линч М. (ноябрь 2012 г.). «Гипотеза дрейфового барьера и эволюция скорости мутаций» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (45): 18488–92. Бибкод : 2012PNAS..10918488S . дои : 10.1073/pnas.1216223109 . ПМЦ 3494944 . ПМИД 23077252 .
- ^ Кротти С., Кэмерон С.Э., Андино Р. (июнь 2001 г.). «Катастрофа ошибки РНК-вируса: прямой молекулярный тест с использованием рибавирина» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (12): 6895–900. Бибкод : 2001PNAS...98.6895C . дои : 10.1073/pnas.111085598 . ПМЦ 34449 . ПМИД 11371613 .
- ^ Рамбо А., Посада Д., Крэндалл К.А., Холмс Э.К. (январь 2004 г.). «Причины и последствия эволюции ВИЧ» . Обзоры природы Генетика . 5 (52–61): 52–61. дои : 10.1038/nrg1246 . ПМИД 14708016 . S2CID 5790569 . Архивировано из оригинала 09.11.2019 . Проверено 28 мая 2019 г.
Внешние ссылки
[ редактировать ] СМИ, связанные со скоростью мутаций, на Викискладе?
СМИ, связанные со скоростью мутаций, на Викискладе?