Дуррин
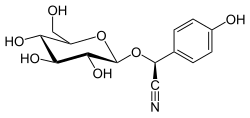
| |
| Имена | |
|---|---|
| Название ИЮПАК
( S )-(β- D -глюкопиранозилокси)(4-гидроксифенил)ацетонитрил
| |
| Систематическое название ИЮПАК
( S )-(4-Гидроксифенил){[(2 R ,3 R ,4 S ,5 S ,6 R )-3,4,5-тригидрокси-6-(гидроксиметил)оксан-2-ил]окси}ацетонитрил | |
| Другие имена
( S )-4-гидроксиалмонднитрил-β- D -глюкопиранозид
| |
| Идентификаторы | |
3D model ( JSmol )
|
|
| ХимическийПаук | |
| Информационная карта ECHA | 100.007.163 |
ПабХим CID
|
|
| НЕКОТОРЫЙ | |
Панель управления CompTox ( EPA )
|
|
| Характеристики | |
| С 14 Ч 17 НЕТ 7 | |
| Молярная масса | 311.29 g/mol |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа).
| |
Дхуррин — цианогенный гликозид, вырабатываемый многими растениями . Обнаруженный в 1906 году во многих сортах сорго как виновник отравления скота цианистым водородом , дуррин чаще всего ассоциируется с сорго двухцветным . [ 1 ] организм, используемый для картирования биосинтеза дуррина из тирозина . Имя Дуррин происходит от арабского слова, обозначающего сорго.
Биосинтез
[ редактировать ]Регулирование у сорго двухцветного
[ редактировать ]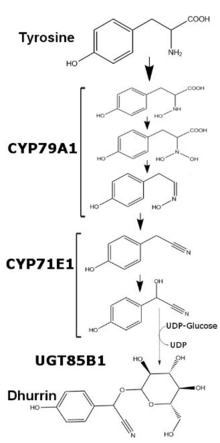
У Sorghum bicolor выработка дуррина регулируется на уровне транскрипции и варьируется в зависимости от возраста растения и доступных питательных веществ. Содержание дуррина в S. bicolor можно коррелировать с количеством мРНК и транслируемого белка ферментов CYP79A1 и CYP71E1, двух мембраносвязанных членов цитохрома P450 суперсемейства . Хотя транскрипция и трансляция этих двух ферментов относительно выше в первые несколько дней роста, транскрипция значительно снижается после одной недели роста. После пяти недель роста транскрипция и трансляция обоих ферментов в листьях становится необнаружимой, в то время как стебли указанных растений сохраняют минимальное производство обоих ферментов. При добавлении избытка нитратов транскрипция обоих ферментов увеличивается, хотя и не до уровней, наблюдаемых на ранних стадиях развития. [ 2 ] Последний фермент синтеза дуррина, UGT85B1, представляет собой растворимый фермент, который обменивает глюкозу из УДФ-глюкозы на агликон дуррина и образует гликозидную связь .
Трансгенный синтез
[ редактировать ]добавления CYP79A1 и CYP71E1 в геномы Arabidopsis thaliana и Nicotiana tabacum достаточно для образования дуррина. Было показано, что [ 3 ] Оба этих фермента достаточны и необходимы для производства дуррина, поскольку удаление гена CYP79A1 из генома сорго двухцветного приводит к тому, что в растениях отсутствует содержание дуррина. Этот штамм теоретически можно использовать как более безопасную культуру на корм в засушливых регионах, где сорго является единственным доступным зерном. Биосинтез дуррина in vitro был построен как в микросомах , выделенных из сорго двухцветного, проростков так и в мицеллах . [ 4 ]
Токсичность
[ редактировать ]Млекопитающие
[ редактировать ]Кишечник млекопитающих содержит множество глюкозидаз , которые эффективно гидролизуют гликозидные связи. При гидролизе гликозидной связи агликон дуррина быстро разлагается с образованием цианистого водорода, который затем всасывается в кровоток. Смертельная доза дуррина для людей и других млекопитающих теоретически высока, поскольку на молекулу дуррина образуется одна молекула цианистого водорода. Массовое содержание дуррина в сорго относительно невелико по отношению к общему растительному веществу. Таким образом, человеку потребуется съесть значительно большое количество сырого сорго, прежде чем он почувствует побочные эффекты. В засушливых условиях сорго является лучшим вариантом зерна и кормов для зерновых культур, поскольку оно может противостоять экстремальным засушливым условиям. [ 5 ] Животные, потребляющие сырое сорго в качестве корма, с гораздо большей вероятностью съедят количество, содержащее смертельную дозу дуррина для их соответствующего вида, что может привести к гибели животных из-за отравления цианистым водородом.
В качестве средства от насекомых
[ редактировать ]В ответ на внешнее повреждение стебля сорта сорго могут выделять в месте повреждения дуррин. Было показано, что этот ответ отпугивает насекомых, поскольку трансгенное сорго, неспособное производить дуррин, пользовалось большим успехом у травоядных насекомых по сравнению с сортами сорго дикого типа . [ 6 ]
Ссылки
[ редактировать ]- ^ Блит, Александр Винтер (13 мая 2013 г.). Яды: их действие и обнаружение. Руководство для химиков-аналитиков и экспертов . США: Чарльз Гриффин и компания. п. 204.
- ^ Буск, Питер Камп (июль 2002 г.). «Синтез дуррина в сорго регулируется на транскрипционном уровне и индуцируется азотным оплодотворением у старых растений» . Физиология растений . 129 (3): 1222–1231. дои : 10.1104/стр.000687 . ПМК 166516 . ПМИД 12114576 .
- ^ Бак, Сорен (август 2000 г.). «Трансгенные растения табака и арабидопсиса, экспрессирующие два многофункциональных фермента цитохрома P450 сорго, CYP79A1 и CYP71E1, являются цианогенными и накапливают метаболиты, полученные из промежуточных продуктов биосинтеза дуррина» . Физиология растений . 123 (4): 1437–1448. дои : 10.1104/стр.123.4.1437 . ПМК 59100 . ПМИД 10938360 .
- ^ Кан, РА (декабрь 1997 г.). «Выделение и восстановление цитохрома P450ox и восстановление in vitro всего пути биосинтеза цианогенного глюкозида дуррина из сорго» . Физиология растений . 115 (4): 1661–1670. дои : 10.1104/стр.115.4.1661 . ПМК 158632 . ПМИД 9414567 .
- ^ Боррелл, Эндрю К. (2014). «Адаптация сорго к засухе связана с развитием кроны, анатомией листьев, ростом корней и поглощением воды» . Журнал экспериментальной ботаники . 65 (21): 6251–6263. дои : 10.1093/jxb/eru232 . ПМК 4223986 . ПМИД 25381433 .
- ^ Кротапалли, Картикея (октябрь 2013 г.). «Передовая генетика с помощью секвенирования генома показывает, что быстрое высвобождение цианида сдерживает травоядных насекомых сорго двухцветного» . Генетика . 195 (2): 309–318. дои : 10.1534/genetics.113.149567 . ПМК 3781961 . ПМИД 23893483 .