Доминирование (генетика)
Эта статья нуждается в дополнительных цитатах для проверки . ( февраль 2018 г. ) |
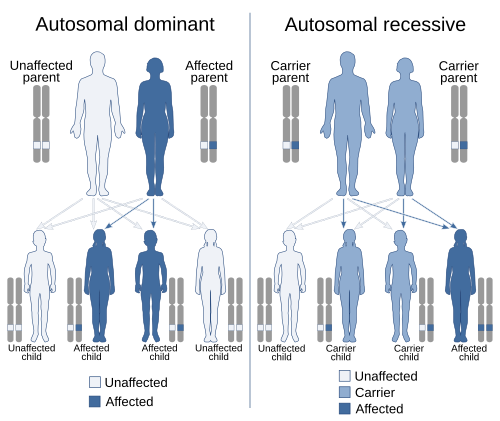
В генетике аллель доминирование — это явление, когда один вариант ( ) гена на хромосоме маскирует или подавляет действие другого варианта того же гена на другую копию хромосомы . [ 1 ] [ 2 ] Первый вариант называется доминантным , а второй — рецессивным . Это состояние наличия двух разных вариантов одного и того же гена на каждой хромосоме изначально вызвано мутацией одного из генов, либо новой ( de novo ), либо унаследованной . Термины аутосомно-доминантный или аутосомно-рецессивный используются для описания вариантов генов на неполовых хромосомах ( аутосомах ) и связанных с ними признаков, тогда как варианты генов на половых хромосомах (аллосомах) называются Х-сцепленными доминантными , Х-сцепленными рецессивными или Y-сцепленными ; у них есть образец наследования и проявления, который зависит от пола как родителя, так и ребенка (см. Связывание полов ). Поскольку существует только одна копия Y-хромосомы , Y-сцепленные признаки не могут быть доминантными или рецессивными. [ 3 ] Кроме того, существуют и другие формы доминирования, такие как неполное доминирование , при котором вариант гена оказывает частичный эффект по сравнению с тем, когда он присутствует на обеих хромосомах, и кодоминирование , при котором разные варианты на каждой хромосоме проявляют связанные с ними признаки. .
Доминирование является ключевым понятием в менделевском наследовании и классической генетике . Буквы и квадраты Пеннета используются для демонстрации принципов доминирования в обучении, заглавные буквы используются для обозначения доминантных аллелей, а строчные — для рецессивных аллелей. Часто цитируемый пример доминирования — наследование формы семян у гороха . Горох может быть круглым, связанным с аллелем R , или морщинистым, связанным с аллелем r . При этом возможны три комбинации аллелей (генотипов): RR , Rr и rr . У RR ( гомозиготных ) особей горошины имеют круглую форму, а у RR (гомозиготных) — горошины морщинистые. У Rr ( гетерозиготных ) особей аллель R маскирует наличие аллеля r , поэтому у этих особей также имеется круглый горошек. Таким образом, аллель R доминантен над аллелем r , а аллель r рецессивен по отношению к R. аллелю [ 4 ]
Доминирование не присуще аллелю или его признаку ( фенотипу ). Это строго относительный эффект между двумя аллелями данного гена любой функции; одна аллель может быть доминантной по отношению ко второй аллели того же гена, рецессивной по отношению к третьей и кодоминантной по отношению к четвертой. Кроме того, один аллель может быть доминантным по одному признаку, но не по другим. [ 5 ] Доминирование отличается от эпистаза , явления, когда аллель одного гена маскирует действие аллелей другого гена . [ 6 ]
Фон
[ редактировать ]
Грегор Иоганн Мендель , «Отец генетики», провозгласил идею доминирования в 1860-х годах. Однако он не был широко известен до начала двадцатого века. Мендель заметил, что для множества признаков садового гороха, связанных с внешним видом семян, семенных коробочек и растений, существовало два отдельных фенотипа, такие как круглые и морщинистые семена, желтые и зеленые семена, красные и белые цветки или высокие и низкие растения. При раздельном разведении растения всегда давали одни и те же фенотипы, поколение за поколением. Однако при скрещивании (интербред) линий с разными фенотипами в потомстве проявлялся один и только один из родительских фенотипов (зеленый, круглый, красный или высокий). Однако при скрещивании этих гибридных растений потомки показали два исходных фенотипа в характерном соотношении 3:1, причем более распространенным фенотипом был фенотип родительских гибридных растений. Мендель рассуждал, что каждый родитель в первом скрещивании был гомозиготным по разным аллелям (один родительский АА, другой родительский аа), что каждый из них передал потомству один аллель, в результате чего все эти гибриды были гетерозиготными (Аа), и что один из двух аллелей в гибридном скрещивании доминировал над выражением другого: A замаскированного a. Окончательное скрещивание двух гетерозигот (Аа X Аа) даст потомство АА, Аа и аа с соотношением генотипов 1:2:1, при этом первые два класса будут показывать фенотип (А), а последний — фенотип (а). , тем самым создавая соотношение фенотипов 3:1.
Мендель не использовал термины ген, аллель, фенотип, генотип, гомозигота и гетерозигота, которые были введены позже. Он действительно ввел обозначения заглавных и строчных букв для доминантных и рецессивных аллелей соответственно, которые используются до сих пор.
В 1928 году британский популяционный генетик Рональд Фишер предположил, что доминирование основано на естественном отборе посредством вклада генов-модификаторов . В 1929 году американский генетик Сьюэлл Райт ответил, что доминирование — это просто физиологическое следствие метаболических путей и относительной необходимости задействованного гена. [ 7 ] [ 8 ] [ 9 ] [ 5 ]
Виды доминирования
[ редактировать ]Полное доминирование (менделевское)
[ редактировать ]При полном доминировании действие одного аллеля в гетерозиготном генотипе полностью маскирует действие другого. Аллель, который маскируется, считается доминантным по отношению к другому аллелю, а замаскированный аллель считается рецессивным . [ 10 ]
Когда мы рассматриваем только один признак, определяемый одной парой генов, мы называем это моногибридным наследованием . Если скрещивание осуществляется между родителями (P-поколение, F0-поколение), которые являются гомозиготно-доминантными и гомозиготно-рецессивными, потомство (F1-поколение) всегда будет иметь гетерозиготный генотип и всегда иметь фенотип, связанный с доминантным геном.
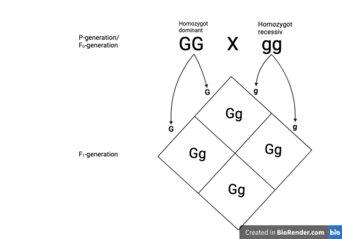

Однако, если поколение F1 в дальнейшем скрестить с поколением F1 (гетерозигота, скрещенная с гетерозиготой), потомство (поколение F2) будет представлять фенотип, связанный с доминантным геном, ¾ раза. Хотя гетерозиготное моногибридное скрещивание может привести к появлению двух вариантов фенотипа, оно может привести к появлению трех вариантов генотипа - гомозиготного доминантного, гетерозиготного и гомозиготного рецессивного соответственно. [ 11 ]
При дигибридном наследовании мы рассматриваем одновременное наследование двух пар генов. Предполагая здесь, что две пары генов расположены в негомологичных хромосомах, так что они не являются спаренными генами (см. Генетическое сцепление ), а наследуются независимо. Рассмотрим теперь скрещивание родителей (Р-поколение) гомозиготных доминантных и рецессивных генотипов соответственно. Потомство (поколение F1) всегда будет гетерозиготным и будет иметь фенотип, связанный с вариантом доминантного аллеля.


Однако при скрещивании поколения F1 возможны четыре фенотипические возможности, и фенотипическое соотношение для поколения F2 всегда будет 9:3:3:1. [ 12 ]
Неполное доминирование (неменделевское)
[ редактировать ]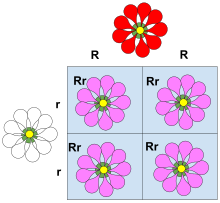
Неполное доминирование (также называемое частичным доминированием , полудоминированием , промежуточным наследованием или иногда ошибочным кодоминированием в генетике рептилий). [ 13 ] ) возникает, когда фенотип гетерозиготного генотипа отличается от фенотипов гомозиготных генотипов и часто является промежуточным по отношению к ним. Фенотипический результат часто проявляется как смешанная форма признаков в гетерозиготном состоянии. Например, цвет цветка львиного зева гомозиготен либо по красному, либо по белому цвету. Когда красный гомозиготный цветок соединяется с белым гомозиготным цветком, в результате получается розовый цветок львиного зева. Розовый львиный зев – результат неполного доминирования. Подобный тип неполного доминирования обнаружен у четырехчасового растения , у которого розовый цвет получается при скрещивании чистопородных родителей с белыми и красными цветками. В количественной генетике , где фенотипы измеряются и обрабатываются численно, если фенотип гетерозиготы находится точно между (численно) фенотипом двух гомозигот, говорят, что фенотип вообще не демонстрирует доминирования , т.е. доминирование существует только тогда, когда мера фенотипа гетерозиготы лежит ближе к одной гомозиготе, чем к другой.
растений поколения F 1 При самоопылении фенотипическое и генотипическое соотношение поколения F 2 будет составлять 1:2:1 (Красный:Розовый:Белый). [ 14 ]
Кодоминирование (неменделевское)
[ редактировать ]


Кодоминирование происходит, когда вклад обоих аллелей виден в фенотипе и ни один аллель не маскирует другой.
Например, в системе групп крови АВО химические модификации гликопротеина ( антигена H) на поверхности клеток крови контролируются тремя аллелями, два из которых кодоминантны друг другу ( I А , я Б ) и доминирует над рецессивным i в локусе АВО . Я А и я Б аллели производят различные модификации. Фермент, кодируемый I А добавляет N-ацетилгалактозамин к мембраносвязанному антигену H. Я Б фермент добавляет галактозу. Аллель i не вызывает модификаций. Таким образом, я А и я Б каждый из аллелей доминирует над i ( I А я А и я А У меня обоих есть группа крови А, и я Б я Б и я Б у обоих людей группа крови B), но у меня А я Б люди имеют обе модификации клеток крови и, следовательно, имеют кровь группы AB, поэтому I А и я Б Говорят, что аллели кодоминантны. [ 14 ]
Другой пример происходит в локусе бета-глобинового компонента гемоглобина , где три молекулярных фенотипа Hb А /Гб А , Хб А /Гб С и Hb С /Гб С все они различимы с помощью электрофореза белков . (Заболевание, вызванное гетерозиготным генотипом, называется серповидно-клеточным признаком и представляет собой более легкое состояние, отличающееся от серповидно-клеточной анемии , поэтому аллели демонстрируют неполное доминирование в отношении анемии, см. выше). Для большинства генных локусов на молекулярном уровне оба аллеля экспрессируются кодоминантно, поскольку оба транскрибируются в РНК . [ 14 ]
Кодоминирование, при котором аллельные продукты сосуществуют в фенотипе, отличается от неполного доминирования, при котором количественное взаимодействие аллельных продуктов приводит к промежуточному фенотипу. Например, при кодоминировании красный гомозиготный цветок и белый гомозиготный цветок дадут потомство с красными и белыми пятнами. При самоопылении растений поколения F1 фенотипическое и генотипическое соотношение поколения F2 будет 1:2:1 (Красный:Пятнистый:Белый). Эти соотношения такие же, как и при неполном доминировании. Опять же, эта классическая терминология неуместна – на самом деле о таких случаях вообще нельзя говорить, что они демонстрируют доминирование. [ 14 ]
Связь с другими генетическими концепциями
[ редактировать ]На доминирование могут влиять различные генетические взаимодействия, и важно оценивать их при определении фенотипических результатов. Множественные аллели , эпистаз и плейотропные гены являются факторами, которые могут влиять на фенотипический результат. [ 15 ]
Множественные аллели
[ редактировать ]Хотя любая особь диплоидного организма имеет не более двух разных аллелей в данном локусе, большинство генов существуют в большом количестве аллельных версий в популяции в целом. Это называется полиморфизмом и вызвано мутациями. Полиморфизм может влиять на отношения доминирования и фенотип, которые наблюдаются в системе групп крови АВО . Ген, отвечающий за группу крови человека, имеет три аллеля; A, B и O, и их взаимодействие приводит к образованию разных групп крови в зависимости от уровня доминирования аллелей по отношению друг к другу. [ 15 ] [ 16 ]
Плейотропные гены
[ редактировать ]Плейотропные гены — это гены, в которых один единственный ген влияет на два или более признаков (фенотип). Это означает, что ген может оказывать доминантное влияние на один признак, но более рецессивное влияние на другой признак. [ 17 ]
Эпистаз
[ редактировать ]Эпистаз – это взаимодействие между несколькими аллелями в разных локусах. Проще говоря, несколько генов для одного фенотипа. Отношения доминирования между аллелями, участвующими в эпистатических взаимодействиях, могут влиять на наблюдаемые фенотипические соотношения у потомства. [ 18 ]
См. также
[ редактировать ]- Двунаправленное доминирование
- Список менделевских черт у человека
- Митохондриальная ДНК
- Паннетт-сквер
- Пенетрантность
- Теоремы суммирования (биохимия)
- Химеризм
Ссылки
[ редактировать ]- ^ «доминирование» . Оксфордские словари онлайн . Издательство Оксфордского университета. Архивировано из оригинала 18 июля 2012 года . Проверено 14 мая 2014 г.
- ^ "выражать" . Оксфордские словари онлайн . Издательство Оксфордского университета. Архивировано из оригинала 18 июля 2012 года . Проверено 14 мая 2014 г.
- ^ Эггерс, Стефани; Синклер, Эндрю (2012). «Определение пола млекопитающих - опыт людей и мышей». Хромосома Res . 20 (1). Дордрехт: Springer-Verlag: 215–238. дои : 10.1007/s10577-012-9274-3 . hdl : 11343/270255 . ISSN 0967-3849 . ПМИД 22290220 .
- ^ Бейтсон, Уильям; Мендель, Грегор (2009). Принципы наследственности Менделя: защита с переводом оригинальных статей Менделя о гибридизации . Издательство Кембриджского университета. дои : 10.1017/CBO9780511694462 . ISBN 978-1108006132 .
- ^ Jump up to: а б Бильярд, Сильвен; Кастрик, Винсент; Ллоуренс, Виолен (2021). «Интегративная биология генетического доминирования» . Biol Rev Camb Philos Soc . 96 (6). Оксфорд, Великобритания: Оксфорд, Великобритания: Blackwell Publishing Ltd: 2925–2942. дои : 10.1111/brv.12786 . ПМЦ 9292577 . ПМИД 34382317 .
- ^ Гриффитс AJF; Гелбарт В.М.; Миллер Дж. Х.; и др. (1999). «Взаимодействие генов приводит к изменению дигибридных соотношений» . Современный генетический анализ . Нью-Йорк: WH Freeman & Company. ISBN 978-0-7167-3118-4 .
- ^ Мэйо О. и Бюргер Р. 1997. Эволюция доминирования: теория, время которой прошло? Архивировано 4 марта 2016 г. в Wayback Machine «Биологические обзоры», том 72, выпуск 1, стр. 97–110.
- ^ Бурге, Д. 1999. Эволюция доминирования. Архивировано 29 августа 2016 г. в Wayback Machine Heredity , том 83, номер 1, стр. 1–4.
- ^ Багери, ХК 2006. Неразрешенные границы эволюционной теории и вопрос о том, как развиваются системы наследования: 75 лет дебатов об эволюции доминирования. Архивировано 2 июля 2019 г. в Wayback Machine «Журнал экспериментальной зоологии, часть B: Молекулярная и эволюционная». Эволюция», том 306B, выпуск 4, стр. 329–359.
- ^ Родригес-Бельтран, Херонимо; Сёрум, Видар; Толл-Риера, Макарена; де ла Вега, Кармен; Пенья-Миллер, Рафаэль; Сан-Мильян, Альваро (2020). «Генетическое доминирование управляет эволюцией и распространением мобильных генетических элементов у бактерий» . Proc Natl Acad Sci США . 117 (27). США: США: Национальная академия наук: 15755–15762. Бибкод : 2020PNAS..11715755R . дои : 10.1073/pnas.2001240117 . ISSN 0027-8424 . ПМК 7355013 . ПМИД 32571917 .
- ^ Труди, ФК Маккей; Роберт, Р. Х. Анхольт (2022). «Наследие Грегора Менделя в количественной генетике» . ПЛОС Биология . 20 (7). Публичная научная библиотека (PLoS): e3001692. дои : 10.1371/journal.pbio.3001692 . ISSN 1544-9173 . ПМЦ 9295954 . ПМИД 35852997 .
- ^ Альбертс, Брюс; Хилд, Ребекка; Хопкин, Карен; Джонсон, Александр; Морган, Дэвид; Робертс, Кейт; Уолтер, Питер (2023). Эссенциальная клеточная биология (Шестое издание; Изд. международных студентов). WW Нортон и компания. ISBN 9781324033394 .
- ^ Булински, Стивен (5 января 2016 г.). «Ускоренный курс генетики рептилий» . Рептилии . Живые мировые СМИ. Архивировано из оригинала 04 февраля 2020 г. Проверено 3 февраля 2023 г.
Термин кодоминантный часто используется как синоним неполной доминантности, но эти два термина имеют разные значения.
- ^ Jump up to: а б с д Браун, Т.А. (2018). Геномы 4 (4-е изд.). Милтон: Милтон: Гарленд Наука. дои : 10.1201/9781315226828 . ISBN 9780815345084 . S2CID 239528980 .
- ^ Jump up to: а б Ингельман-Сундберг, М. (2005). «Генетические полиморфизмы цитохрома P450 2D6 (CYP2D6): клинические последствия, эволюционные аспекты и функциональное разнообразие». Фармакогеномика Дж . 5 (1). США: США: Издательская группа Nature: 6–13. дои : 10.1038/sj.tpj.6500285 . ISSN 1470-269X . ПМИД 15492763 . S2CID 10695794 .
- ^ Ямамото, Ф; Клаузен, Х; Белый, Т; Маркен, Дж; Хакомори, С (1990). «Молекулярно-генетические основы гистогрупповой системы АВО». Природа . 345 (6272): 229–233. Бибкод : 1990Natur.345..229Y . дои : 10.1038/345229a0 . ПМИД 2333095 . S2CID 4237562 .
- ^ Ду, Цинчжан; Тянь, Цзясин; Ян, Сяохуэй; Пан, Вэй; Сюй, Баохуа; Ли, Байлянь; Ингварссон, Пяр К.; Чжан, Дэцян (2015). «Идентификация аддитивных, доминантных и эпистатических вариаций, обусловленных ключевыми генами пути биосинтеза целлюлозы у Populus tomentosa» . ДНК Рез . 22 (1). Англия: Англия: Издательство Оксфордского университета: 53–67. дои : 10.1093/dnares/dsu040 . ISSN 1340-2838 . ПМК 4379978 . ПМИД 25428896 .
- ^ Филлипс, Патрик С (2008). «Эпистаз – существенная роль взаимодействий генов в строении и эволюции генетических систем» . Нат преподобный Жене . 9 (11). Лондон: Лондон: Издательская группа Nature: 855–867. дои : 10.1038/nrg2452 . ISSN 1471-0056 . ПМК 2689140 . ПМИД 18852697 .
- «Онлайн-заметки по биологии 2250 - Принципы генетики» . Мемориальный университет Ньюфаундленда.
- Онлайн-менделевское наследование у человека (OMIM): гемоглобин — бета-локус; HBB - 141900 — Серповидноклеточная анемия
- Онлайн-менделевское наследование у человека (OMIM): гликозилтрансфераза ABO - 110300 - группы крови ABO