Кортикальная гранула
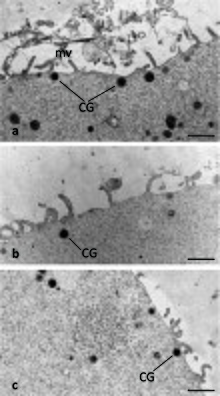
Кортикальные гранулы представляют собой регуляторные секреторные органеллы (диаметром от 0,2 до 0,6 мкм), обнаруженные внутри ооцитов и наиболее связанные с предотвращением полиспермии после оплодотворения. [ 1 ] Кортикальные гранулы встречаются у всех млекопитающих , многих позвоночных и некоторых беспозвоночных . [ 2 ] Внутри ооцита кортикальные гранулы расположены вдоль коры, области, наиболее удаленной от центра клетки. После оплодотворения сигнальный путь заставляет кортикальные гранулы сливаться с клеточной мембраной ооцита и высвобождать их содержимое во внеклеточный матрикс ооцита. Этот экзоцитоз кортикальных гранул известен как корковая реакция . У млекопитающих внеклеточный матрикс ооцита включает окружающий слой перивителлинового пространства , пеллюцидную зону и, наконец, кумулюсные клетки . Экспериментальные данные показали, что высвободившееся содержимое кортикальных гранул модифицирует внеклеточный матрикс ооцита, особенно пеллюцидную зону. Это изменение компонентов зоны пеллюцида известно как реакция зоны. Корковая реакция возникает не у всех млекопитающих, что указывает на вероятность других функциональных целей кортикальных гранул. [ 1 ] Помимо модификации внеклеточного матрикса ооцита и создания блока полиспермии, экзоцитоз кортикальных гранул может также способствовать защите и поддержке развивающегося эмбриона во время предимплантации. [ 3 ] После того как кортикальные гранулы завершают свои функции, ооцит их не восполняет. [ 1 ]
Формирование
[ редактировать ]Формирование кортикальных гранул происходит на ранних стадиях роста ооцита. Более конкретно, у человека, обезьяны, хомяка и кролика кортикальные гранулы образуются, когда фолликул яичника становится многослойным. У крыс и мышей кортикальные гранулы наблюдались раньше при развитии фолликула, когда фолликул яичника однослойный. На ранних стадиях роста ооцитов комплекс Гольджи увеличивается в размерах, пролиферирует и производит небольшие пузырьки, которые мигрируют в подкорковую область клетки. Эти маленькие пузырьки сливаются друг с другом, образуя зрелые кортикальные гранулы, которые, таким образом, становятся отдельными от Гольджи образованиями. [ 1 ] У некоторых организмов, например у хомяков, секретируемая везикула из Гольджи может сливаться с секретируемой везикулой из шероховатой эндоплазматической сети, образуя в конечном итоге кортикальную гранулу. [ 4 ] У млекопитающих ооцит непрерывно производит и перемещает кортикальные гранулы в кору до тех пор, пока не произойдет овуляция . На моделях млекопитающих и немлекопитающих было показано, что миграция кортикальных гранул зависит от процессов цитоскелета , особенно от активности микрофиламентов . Для млекопитающих миграция кортикальных гранул считается показателем зрелости ооцита и организации органелл. [ 1 ]
Распределение
[ редактировать ]В результате транслокации кортикальные гранулы равномерно распределяются по коре ооцита. Однако у грызунов наблюдалось, что некоторые кортикальные гранулы перестраиваются, оставляя пространство среди оставшихся кортикальных гранул. свободным от кортикальных гранул (CGFD), и оно наблюдается в обеих областях мейотического веретена клетки во время метафазы I и метафазы II мейоза Это пространство называется доменом , . CGFD не наблюдались в ооцитах кошек, лошадей, крупного рогатого скота, свиней и человека. Исследования ооцитов грызунов показывают, что определенные кортикальные гранулы подвергаются перераспределению и/или экзоцитозу на протяжении мейотического цикла, тем самым создавая CGFD. Более конкретно, доказательства включают увеличение количества кортикальных гранул, окружающих CGFD, и уменьшение общего количества кортикальных гранул в клетке во время мейотического цикла. Кроме того, некоторые экзоцитозные события кортикальных гранул перед оплодотворением происходят в борозде расщепления клетки одновременно с полярным тельцем. формирование. [ 1 ]
Существует множество гипотез относительно биологической функции CGFD и экзоцитоза кортикальных гранул перед оплодотворением. Напр., образование CGFDs может быть механизмом ооцита, позволяющим сохранять больше кортикальных гранул для будущего использования, а не терять их из-за полярных телец, когда полярные тельца выдавливаются из клетки. Поскольку некоторые высвобождаемые кортикальные гранулы происходят из области вблизи мейотических веретен, исследователи также предположили, что высвободившиеся кортикальные гранулы могут модифицировать внеклеточный матрикс ооцита так, что сперматозоиды не могут связываться в этой области. Если бы сперматозоид связывался в этой области, отцовская ДНК по мере ее деконденсации могла бы нарушить целостность материнской ДНК из-за ее близости. Такая блокировка сперматозоидов в определенном месте называется локальной блокировкой. Учитывая, что ооциты грызунов имеют примерно на 75% меньшую площадь поверхности, чем ооциты более крупных видов млекопитающих, связывание сперматозоидов в этой области более вероятно, что, возможно, вызывает необходимость локального блокирования. Исследователи также предполагают, что ооцит высвобождает некоторые кортикальные гранулы перед оплодотворением, чтобы внести незначительные изменения во внеклеточный матрикс ооцита, так что связывание ограничивается только сперматозоидами, способными связываться, несмотря на эти незначительные модификации. [ 1 ]
Регулирование
[ редактировать ]После оплодотворения и до высвобождения кальция инозитолфосфата инициируется сигнальный каскад вторичных молекул-мессенджеров, таких как инозитол-1,4,5-трифосфат (IP3) и диацилглицерин (PIP2). В ходе этого процесса концентрация (DAG), увеличивается. Было показано, что инозитол-1,4,5-трифосфат взаимодействует с эндоплазматическим ретикулумом, вызывая высвобождение запасов кальция из эндоплазматического ретикулума. Еще раз подчеркивая взаимосвязь между эндоплазматическим ретикулумом и кортикальными гранулами, во время созревания ооцита наблюдалось, что эндоплазматический ретикулум либо растет, либо мигрирует ближе к области кортикальных гранул. Помимо кальция, диацилглицерин, по-видимому, инициирует корковую реакцию. [ 5 ] Диациглицерин также активирует протеинкиназу C (PKC), которая также способствует корковой реакции. сперматозоидов Было показано, что после оплодотворения протеинкиназа C способствует акросомальному экзоцитозу , процессу, гомологическому экзоцитозу кортикальных гранул ооцита. Подобно протеинкиназы С, кальмодулин активируется кальцием, еще больше стимулируя корковую реакцию. [ 6 ]
Это увеличение кальция происходит в виде одной волны у иглокожих и в виде множественных волн у млекопитающих. Было показано, что экзоцитоз кортикальных гранул происходит непосредственно после волны кальция. Например, в оплодотворенной яйце морского ежа было показано, что экзоцитоз кортикальных гранул сразу же следует за увеличением кальция примерно через 6 секунд. У млекопитающих первая волна кальция возникает в течение 1–4 минут после оплодотворения, а экзоцитоз кортикальных гранул происходит в течение 5–30 минут после оплодотворения. Более того, когда волны кальция подавлялись экспериментально, экзоцитоза кортикальных гранул и/или изменений во внеклеточном матриксе не происходило. Как показано на неоплодотворенных ооцитах позвоночных, экзоцитоз кортикальных гранул индуцируется при искусственном повышении содержания кальция. [ 7 ]
Считается также, что повышенное содержание кальция активирует белки, деполимеризующие актин , такие как гельзолин и сциндерин. У млекопитающих эти деполимеризующие актин белки служат для разборки кортикального актина, тем самым обеспечивая пространство для транслокации кортикальных гранул к плазматической мембране. [ 7 ]
Ооцит приобретает способность завершать экзоцитоз кортикальных гранул к моменту достижения поздней зрелости. Более конкретно, например, у мышей способность подвергаться экзоцитозу кортикальных гранул возникает через некоторое время между метафазой I и метафазой II мейоза, что также происходит за 5 часов до наступления овуляции. Было показано, что ооцит достигает максимальной способности высвобождать кальций на этой же клеточной стадии, между метафазой I и метафазой II, что еще больше подчеркивает кальций-зависимость экзоцитоза кортикальных гранул. [ 7 ]
Состав органелл
[ редактировать ]Хотя весь состав кортикальных гранул еще не идентифицирован, следующие молекулы связаны с содержимым кортикальных гранул млекопитающих:
Гликозилированные компоненты: Было показано, что кортикальные гранулы млекопитающих содержат высокий уровень углеводов. Кроме того, многие из этих углеводов являются компонентами гликозилированных молекул, таких как маннозилированные белки, α-D-ацетилгалактозамин, N-ацетилглюкозамин , N-ацетиллактозамин , N-ацетилнейраминовая кислота , DN-ацетилгалактозамин, N-ацетилгалактозамин и N-гликолилнейраминовая кислота . Например, считается, что некоторые маннозилированные белки вносят вклад в структуру оболочки кортикальных гранул. [ 1 ]
Протеиназы : Протеиназы, присутствующие в гранулах коры млекопитающих, в первую очередь служат для модификации прозрачной зоны во время реакции зоны. Некоторыми связанными протеиназами кортикальных гранул являются трипсин -подобная протеиназа, протеиназа ZP2 и активатор плазминогена тканевого типа (tPA). И протеиназа ZP2, и трипсиноподобная протеиназа способствуют предотвращению полиспермии. Как следует из названия, протеиназа ZP2 протеолизирует ZP2 во время реакции зоны. тканевого типа Активатор плазминогена (tPA) представляет собой сериновую протеиназу, которая превращает плазминоген в его активированную форму, плазмин . Присутствие тканевого плазминогена зафиксировано при кортикальной реакции. Однако, несмотря на эту связь с кортикальной реакцией, еще предстоит найти доказательства, подтверждающие, что активатор плазминогена тканевого типа является компонентом кортикальных гранул. Более того, мРНК, кодирующая тканевой активатор плазминогена, не транслируется до тех пор, пока внутри ооцита не сформируется большая часть кортикальных гранул. [ 1 ]
Овопероксидаза: белок овопероксидаза, скорее всего, действует как катализатор, который сшивает остатки тирозина, обнаруженные в зоне пеллюцида . Это сшивание способствует отверждению зоны пеллюцида. [ 1 ]
Кальретикулин : данные указывают на присутствие белка кальретикулина в кортикальных гранулах. Исследователи предположили, что кальретикулин служит белком-шапероном для других компонентов кортикальных гранул, способствуя предотвращению полиспермии. Помимо предотвращения полиспермии, кальретикулин может также ингибировать определенные гликопротеины, которые способствуют взаимодействию между ооцитом и спермой. С другой стороны, различные исследования показали, что кальретикулин может высвобождаться из везикул, отличных от кортикальных гранул. Более того, при экзоцитозе этот кальретикулин взаимодействует с цитоскелетом ооцита, тем самым обеспечивая передачу трансмембранных сигналов для продолжения клеточного цикла. [ 1 ]
N-ацетилглюкозаминидаза : экспериментально обнаруженная в кортикальных гранулах мышей, N-ацетилглюкозаминидаза представляет собой гликозидазу , которая гидролизует остатки N-ацетилглюкозамина, расположенные в прозрачной зоне. N-ацетилглюкозамины в зоне пеллюцида обычно способствуют связыванию сперматозоидов. [ 1 ] Таким образом, N-ацетилглюкозаминидаза способствует предотвращению полиспермии.
p32 : Название p32 относится к молекулярной массе белка, 32 кДа. После высвобождения из кортикальных гранул р32, по-видимому, либо кратковременно функционирует, либо подвергается модификации вскоре после оплодотворения, поскольку на эмбрионе присутствуют лишь небольшие количества р32. Исследования также показывают, что р32 не способствует предотвращению полиспермии. [ 1 ]
Пептидиларгининдеиминаза (PAD/ABL2 антиген/p75): Пептидиларгининдеиминаза представляет собой секреторный белок в кортикальных гранулах, который способствует регуляции развития эмбриона до момента имплантации. Альтернативное название пептидиларгининдеиминазы, p75, относится к ее молекулярной массе 75 кДа. [ 1 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н Лю, Мин (17 ноября 2011 г.). «Биология и динамика корковых гранул млекопитающих» . Репродуктивная биология и эндокринология . 9 (1): 149. дои : 10.1186/1477-7827-9-149 . ПМК 3228701 . ПМИД 22088197 .
- ^ Вессель, Гэри М.; Брукс, Жаклин М.; Грин, Эмма; Хейли, Шейла; Воронина Екатерина; Вонг, Джулиан; Зайдфудим, Виктор; Коннер, Шон (2001). Биология кортикальных гранул . Том. 209. С. 117–206. дои : 10.1016/s0074-7696(01)09012-x . ISBN 9780123646132 . ПМИД 11580200 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Худбхой, Таня; Талбот, П. (декабрь 1994 г.). «Кортикальные гранулы млекопитающих: содержание, судьба и функции» (PDF) . Молекулярное воспроизводство и развитие . 39 (4): 439–448. дои : 10.1002/mrd.1080390413 . ПМИД 7893493 . S2CID 640323 . Проверено 13 мая 2015 г.
- ^ Гуляс, Б.Дж. (1980). Кортикальные гранулы яиц млекопитающих . Том. 63. С. 357–392. дои : 10.1016/S0074-7696(08)61762-3 . ISBN 9780123644633 . ПМИД 395132 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Дучибелла, Том (1996). «Корковая реакция и развитие активационной компетентности в ооцитах млекопитающих» . Обновление репродукции человека . 2 (1): 29–42. дои : 10.1093/humupd/2.1.29 . ПМИД 9079401 .
- ^ Сунь, Цин-Юань (1 июля 2003 г.). «Клеточные и молекулярные механизмы, приводящие к кортикальной реакции и блоку полиспермии в яйцах млекопитающих» . Микроскопические исследования и техника . 61 (4): 342–348. дои : 10.1002/jemt.10347 . ПМИД 12811739 . S2CID 22884757 .
- ^ Перейти обратно: а б с Эбботт, Алабама; Дучибелла, Т. (1 июля 2001 г.). «Кальций и контроль экзоцитоза кортикальных гранул млекопитающих» . Границы бионауки . 6 (1): d792–806. doi : 10.2741/Эбботт . ПМИД 11438440 .