Эмбриональная диапауза
| Часть серии о |
| животных Покой |
|---|
 |
Эмбриональная диапауза [а] ( отсроченная имплантация у млекопитающих) — репродуктивная стратегия, используемая рядом видов животных разных биологических классов . У более чем 130 видов млекопитающих , у которых это имеет место, процесс происходит на бластоцисты стадии эмбрионального развития . [1] характеризуется резким снижением или полным прекращением митотической активности чаще всего в G0 , прекращающейся или G1 . и фазе деления [2]
При плацентарно-эмбриональной диапаузе бластоциста не имплантируется сразу в матку после того, как половое размножение привело к образованию зиготы , а скорее остается в этом неделящемся состоянии покоя до тех пор, пока условия не позволят прикрепляться к стенке матки происходить как обычно. [3] В результате нормальный период беременности продлевается на видоспецифическое время. [4] [5]
Диапауза дает потомству преимущество в выживании, поскольку рождение или появление детенышей можно приурочить к наиболее благоприятным условиям, независимо от того, когда происходит спаривание или продолжительность беременности; любое такое увеличение выживаемости потомства дает эволюционное преимущество.
Эволюционное значение
[ редактировать ]Организмы, переживающие эмбриональную диапаузу, способны синхронизировать рождение потомства с наиболее благоприятными условиями для репродуктивного успеха, независимо от того, когда происходило спаривание. [3] Вызвать эмбриональную диапаузу может множество различных факторов, таких как время года, температура, лактация и наличие пищи. [3]
Эмбриональная диапауза — относительно широко распространенное явление за пределами млекопитающих; известно, что она встречается в репродуктивных циклах многих насекомых, нематод, рыб и других позвоночных, не относящихся к млекопитающим. [6] Это наблюдалось примерно у 130 видов млекопитающих. [7] что составляет менее двух процентов всех видов млекопитающих. [8] К ним относятся некоторые грызуны , медведи , броненосцы , куньи (например, ласки и барсуки ) и сумчатые (например, кенгуру ). В некоторых группах есть только один вид, который подвергается эмбриональной диапаузе, например, косуля из отряда парнокопытных . [5]
Была достигнута экспериментальная индукция прерывистого эмбрионального развития у видов, которые в природе не подвергаются спонтанной эмбриональной диапаузе; была успешно продемонстрирована обратимая задержка развития. Это может быть свидетельством эволюционного значения этого явления, поскольку скрытая способность к диапаузе потенциально присутствует у гораздо более широкого сегмента видов, чем та, которая, как известно, встречается в природе. [8] [9]
Общий механизм
[ редактировать ]Все многоклеточные организмы, по их замыслу, начинаются с небольшого количества клеток и растут и развиваются только по мере деления этих клеток. У организмов, способных к эмбриональной диапаузе, в неидеальных репродуктивных условиях происходит прекращение деления клеток, что препятствует росту и созреванию эмбриона, задерживая созревание эмбриона до тех пор, пока условия не станут достаточно идеальными, чтобы способствовать выживанию потомства. , а в некоторых случаях и мать.
Регуляция клеточного цикла, связанная с эмбриональной диапаузой, связана с геном дакапо у плодовой мушки, ответственным за ингибирование образования комплексов циклин E-cdk2, необходимых для синтеза ДНК. Имеются также данные, указывающие на активацию гена 1 транслокации В-клеток (Btg1) у эмбрионов мышей во время диапаузы, другого известного регулятора клеточного цикла, ответственного за ингибирование перехода от G 0 /G 1 . Другие исследования, наоборот, продемонстрировали отсутствие участия более распространенных регуляторов клеточного цикла, таких как р53, в плацентарной модели эмбриональной диапаузы. [2] Хотя большая часть молекулярной регуляции, участвующей в активации спящих бластоцист , охарактеризована, мало широко применимых характеристик доступно в отношении вступления в диапаузу и условий, которые позволяют бластоцисте оставаться в спящем состоянии. Как только эмбрион выходит из состояния диапаузы и возобновляет нормальное развитие, никаких побочных эффектов не наблюдается. [10]
В частности, в плацентарно-эмбриональной диапаузе это прекращение вызвано преднамеренной неспособностью бластоцисты имплантироваться в стенку матки, что является важным компонентом прогрессирования развития у этих видов. [11] Гормоны, связанные с неудачной имплантацией, также способствуют остановке эмбриона. [9]
Типы
[ редактировать ]Существуют две различные формы эмбриональной диапаузы, характеризующиеся разными условиями возникновения. Факультативная диапауза возникает в ответ на определенные факторы окружающей среды или метаболические стрессоры, такие как резкие изменения температуры, кормление или лактация. [10] Облигатная диапауза регулярно возникает в репродуктивном цикле пораженных видов и часто связана с сезонными изменениями и фотопериодом. [10]
Факультативная диапауза
[ редактировать ]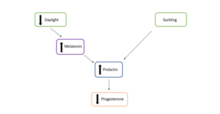
Факультативная диапауза регулируется несколькими факторами, включая материнское окружение и работоспособность яичников, гипофиз, метаболический стресс и лактацию. [2]
Что касается многих других регуляторов этой формы диапаузы, у плацентарных млекопитающих факультативная диапауза чаще всего является результатом оплодотворения вскоре после рождения предыдущего помета. Последующое кормление детенышей грудью во время лактации способствует пролактина высвобождению . Это, в свою очередь, снижает прогестерона секрецию желтым телом у беременной женщины. Желтое тело является временным эндокринным органом , который формируется из остатков клеток фолликула яичника после того , как оно выпустило зрелую яйцеклетку . Основная функция желтого тела — секреция прогестерона во время беременности для поддержания необходимой среды матки . Пролактин, воздействуя на желтое тело, приводит к снижению уровня прогестерона ниже оптимальной концентрации и, следовательно, вызывает эмбриональную факультативную диапаузу.
Каждый вид , подвергающийся факультативной диапаузе, имеет тенденцию иметь определенную стадию развития , генетически детерминированную, на которой инициируется этот процесс. Эта форма диапаузы наиболее хорошо изучена у грызунов и сумчатых. [2] но был идентифицирован у многих других видов, включая немлекопитающих. Неясно, насколько хорошо механизмы, изученные для начала, поддержания и выхода из факультативной диапаузы на модели грызунов, применимы к этим другим видам.
Облигатная диапауза
[ редактировать ]Облигатная (прил.: по необходимости) диапауза (она же сезонная отложенная имплантация) — это механизм, обеспечивающий приурочивание рождения потомства к оптимальным условиям окружающей среды, чтобы обеспечить максимальную выживаемость. [12] [9] Предлагаемый механизм заключается в разделении зачатия и родов (родов), чтобы каждое из них могло произойти в наиболее благоприятное время года. [9]
Облигатная диапауза активируется и деактивируется изменением количества световых часов в течение суток (фотопериод) и, следовательно, происходит в определенные времена года. [10] Хотя облигатная диапауза встречается у многих видов в разных группах, продолжительность диапаузы существенно различается. У западных пятнистых скунсов ( Spilogale gracilis ) диапауза составляет около 200 дней, тогда как у американских норок ( Neogale vison ) диапауза составляет всего около четырнадцати дней. [10]
Подобно факультативной диапаузе, ряд гормональных изменений останавливает развитие бластоцисты до имплантации , предотвращая дальнейший рост эмбриона. Однако при облигатной диапаузе бластоциста в каждом репродуктивном сезоне переходит в состояние покоя. Это означает, что каждая бластоциста, которую производит мать, вступает в период диапаузы. [10]
Тщательная регуляция облигатной диапаузы необходима для выживания матери и потомства. Преждевременная диапауза может привести к утрате возможностей для роста и размножения, а поздняя диапауза может привести к смерти из-за неблагоприятных условий. [13]
Накануне весеннего равноденствия , [б] фотопериод менее 12 часов. Это увеличивает выработку мелатонина в шишковидной железе . Из-за ингибирующей связи между мелатонином и пролактином это увеличение уровня мелатонина снижает секрецию пролактина гипофизом . Снижение уровня пролактина, следовательно, снижает выработку прогестерона в желтом теле , предотвращая развитие бластоцисты. Это вызывает эмбриональную диапаузу. [10]
После весеннего равноденствия фотопериод превышает 12 часов. Это снижает выработку мелатонина в шишковидной железе и, следовательно, увеличивает выработку пролактина и прогестерона в гипофизе и желтом теле соответственно. [10]
Увеличение пролактина индуцирует экспрессию гена Odc ( орнитиндекарбоксилазы ). продуцирует белок ODC, фермент, ограничивающий скорость производства полиамина путресцина в Ген Odc среде матки . Присутствие путресцина может указывать на его роль в индукции выхода эмбриона из облигатной диапаузы. [10]
Эмбриональные стволовые клетки
[ редактировать ]Эмбриональные стволовые клетки (ЭСК) могут позволить глубже понять механизмы, контролирующие эмбриональную диапаузу. [13] Это связано с тем, что ЭСК и диапаузирующие бластоцисты имеют очень схожие профили транскриптома. [13] ЭСК происходят из недифференцированных клеток внутренней массы бластоцист эмбриона и обладают способностью к постоянной пролиферации in vitro. [13] ЭСК в основном получают из моделей мышей, в тот момент, когда ЭСК достигают оптимальной эффективности и способны входить в диапаузу. [3]
И диапаузирующие бластоцисты, и ЭСК имеют сходство профиля транскриптома, включая подавление метаболизма , биосинтеза и экспрессии генов . путей [3] Эти сходства позволяют использовать ESC в качестве клеточной модели для идентификации молекулярных факторов, которые регулируют эмбриональную диапаузу. [13]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Диапауза , из английского конца XIX века: диа- «сквозь» + пауза- «задержка». [ нужна ссылка ]
- ^ Весеннее равноденствие — это мартовское равноденствие в северном полушарии и сентябрьское равноденствие в южном полушарии .
Ссылки
[ редактировать ]- ^ «Обзор: Эмбриональная диапауза у европейской косули – замедлена, но не остановлена – ScienceDirect» .
- ^ Jump up to: а б с д Лопес Ф.Л., Десмаре Х.А., Мерфи Б.Д. (декабрь 2004 г.). «Эмбриональная диапауза и ее регуляция» . Размножение . 128 (6): 669–678. дои : 10.1530/rep.1.00444 . ПМИД 15579584 .
- ^ Jump up to: а б с д и Ренфри М.Б., Фенелон Дж.К. (сентябрь 2017 г.). «Загадка эмбриональной диапаузы» . Разработка . 144 (18): 3199–3210. дои : 10.1242/dev.148213 . ПМИД 28928280 . S2CID 6441064 .
- ^ Демарэ Дж.А., Бординьон В., Лопес Ф.Л., Смит Л.К., Мерфи Б.Д. (март 2004 г.). «Выход зародыша норки из облигатной диапаузы» . Биология размножения . 70 (3): 662–670. doi : 10.1095/biolreprod.103.023572 . ПМИД 14585805 . S2CID 38759201 .
- ^ Jump up to: а б Ренфри М.Б., Шоу Дж. (март 2000 г.). «Диапауза». Ежегодный обзор физиологии . 62 (1): 353–375. дои : 10.1146/annurev.phyol.62.1.353 . ПМИД 10845095 .
- ^ Чалар С, Кливио Дж, Монтань Х, Костабиль А, Лима А, Папа Н.Г., Беруа Н., Аресо М.Дж. (2021). «Задержка эмбрионального развития у однолетнего убийцы Austrolebias charrua: протеомный подход к диапаузе III» . ПЛОС ОДИН . 16 (6): e0251820. дои : 10.1371/journal.pone.0251820 . ПМЦ 8177498 . ПМИД 34086690 .
- ^ Фенелон Дж.К., Банерджи А., Мерфи Б.Д. (2014). «Эмбриональная диапауза: развитие приостановлено» . Международный журнал биологии развития . 58 (2–4): 163–174. дои : 10.1387/ijdb.140074bm . ПМИД 25023682 .
- ^ Jump up to: а б Птак Г.Е., Таккони Э., Черник М., Тоски П., Модлински Дж.А., Лой П. (12 марта 2012 г.). «Эмбриональная диапауза сохраняется у млекопитающих» . ПЛОС ОДИН . 7 (3): e33027. Бибкод : 2012PLoSO...733027P . дои : 10.1371/journal.pone.0033027 . ПМК 3299720 . ПМИД 22427933 .
- ^ Jump up to: а б с д Мерфи Б.Д. (декабрь 2012 г.). «Эмбриональная диапауза: прогресс в понимании загадки сезонной отсроченной имплантации». Размножение домашних животных . 47 (Приложение 6): 121–124. дои : 10.1111/rda.12046 . ПМИД 23279480 .
- ^ Jump up to: а б с д и ж г час я Дэн Л, Ли С, Чен Л, Лю Ю, Хоу Р, Чжоу Икс (ноябрь 2018 г.). «Достижения исследований эмбриональной диапаузы у млекопитающих». Наука о воспроизводстве животных . 198 : 1–10. doi : 10.1016/j.anireprosci.2018.09.009 . ПМИД 30266523 . S2CID 52884156 .
- ^ Аплин Дж. Д., Кимбер С. Дж. (июль 2004 г.). «Взаимодействие трофобласта и матки при имплантации» . Репродуктивная биология и эндокринология . 2 (1): 48. дои : 10.1186/1477-7827-2-48 . ПМК 471567 . ПМИД 15236654 .
- ^ Хусейн А.М., Балачандар Н., Матье Дж., Руохола-Бейкер Х. (сентябрь 2022 г.). «Молекулярные регуляторы эмбриональной диапаузы и ракового диапаузоподобного состояния» . Клетки . 11 (19): 2929. doi : 10.3390/cells11192929 . ПМЦ 9562880 . ПМИД 36230891 .
- ^ Jump up to: а б с д и Сайгин Д., Табиб Т., Биттар Х.Э., Валензи Э., Сембрат Дж., Чан С.Ю. и др. (апрель 2020 г.). «Транскрипционное профилирование популяций клеток легких при идиопатической легочной артериальной гипертензии» . Легочное кровообращение . 10 (1): 167–181. дои : 10.1111/een.12792 . ПМК 7052475 . ПМИД 32166015 . S2CID 202006476 .
Дальнейшее чтение
[ редактировать ]- «Группа по исследованию косули» . Проверено 5 апреля 2008 г.