Темноглазый Юнко
| Темноглазый Юнко | |
|---|---|

| |
| Продолжительность: 21 секунда. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Птицы |
| Заказ: | Воробьинообразные |
| Семья: | Воробьиные |
| Род: | Торопиться |
| Разновидность: | Дж. зима
|
| Биномиальное имя | |
| Зимний тростник | |
| |
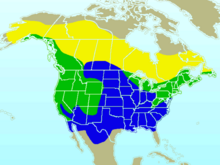
| Приблизительный диапазон в Северной Америке Ареал разведения Круглогодичный ассортимент Зимовка
| |
| Синонимы | |
(но см. текст ) | |

Темноглазый юнко ( Junco hyemalis ) — вид юнко Света , группы мелких сероватых воробьев Нового . Этот вид распространен на большей части территории Северной Америки с умеренным климатом , а летом проникает далеко в Арктику . Это изменчивый вид , во многом похожий на родственного ему лисьего воробья ( Passerella iliaca ), и его систематика до сих пор полностью не выяснена.
Таксономия
[ редактировать ]Темноглазый юнко был официально описан шведским натуралистом Карлом Линнеем в его знаковом 10-м издании Systema Naturae 1758 года как Fringilla hyemalis . Описание состояло лишь из лаконичной реплики « F[ringilla] nigra, ventre albo. («Черный зяблик » с белым брюхом») и заявления о том, что оно прибыло из Америки. [ 2 ] Линней основал свое описание на «Снежной птице», которую Марк Кейтсби описал и проиллюстрировал в своей « Естественной истории Каролины, Флориды и Багамских островов» 1731 года . [ 3 ]
Клюв этой птицы белый: Грудь и брюхо белые. Все остальное тело черное; но местами темный, склоняющийся к свинцовому цвету. В Вирджинии и Каролине они появляются только зимой, а в Сноу их больше всего. Летом их не видно. Уходят ли они на пенсию и размножаются на Севере (что наиболее вероятно) или куда они направляются, покинув эти страны весной, мне неизвестно. [курсив в оригинале] [ 3 ]

Типовой ареал был ограничен Южной Каролиной Союзом американских орнитологов в 1931 году. [ 4 ] [ 5 ] Темноглазый юнко теперь отнесен к роду Юнко , который был введен в 1831 году немецким натуралистом Иоганном Георгом Ваглером . [ 6 ] [ 7 ] Название рода Junco по-испански означает « раш» , от латинского слова «juncus» . [ 8 ] Его современное научное название означает «зимнее юнко» от латинского слова hyemalis «зима». [ 9 ]
Подвиды
[ редактировать ]14 или 15 подвидов . Признано [ 7 ] Эти подвиды группируются в две-три большие или политипные группы и три-четыре мелкие или монотипные группы, все в зависимости от авторитета. Эти группы раньше считались отдельными видами, но они широко скрещиваются в местах соприкосновения. Птицам , пытающимся определить подвиды, рекомендуется обратиться к подробным справочным материалам по идентификации. [ 10 ] [ 11 ]
Группа шиферного цвета
[ редактировать ]
- юнко шиферно-темноглазый ( J. h. hyemalis )
- Каролинское черноглазое юнко ( J. h. carolinensis )
- Юнко кассиаровый темноглазый ( J. h. cismontanus темноглазого юнко грифельного цвета ( J. h. hyemalis ) x джунко орегонский темноглазый ( J. h. oreganus ) ; возможно, гибрид )
У этих двух или трех подвидов темно-серые голова, грудь и верхняя часть тела. Самки буровато-серые, иногда с красновато-коричневыми боками. [ 10 ] Северной Америки Они размножаются в бореальных лесах от Аляски до Ньюфаундленда и на юг до Аппалачей , зимуя на большей части территории Соединенных Штатов. Они относительно распространены во всем своем ареале.
Белокрылая группа
[ редактировать ]
- белокрылый темноглазый юнко ( J.h.aikeni )
У этого подвида голова, грудь и верхняя часть тела средне-серого цвета с белыми полосами на крыльях. Самки окрашены в коричневатый цвет. У него больше белого в хвосте, чем у других 14 подвидов. Это обычный эндемик, гнездящийся в Блэк-Хиллз Южной Дакоты , Вайоминга , Небраски и Монтаны , и зимует на юге до северо-востока Нью-Мексико . [ 12 ] [ 10 ]
Орегон или группа с коричневой спиной
[ редактировать ]
- Юнко монтанский темноглазый ( J.h.montanus )
- Невадский темноглазый юнко ( J. h. mutabilis )
- Юнко орегонский темноглазый ( J. h. oreganus )
- Юнко пойнт Пинос темноглазый ( J. h. pinosus )
- Лагуна Хэнсон темноглазая юнко ( J. h. pontilis )
- Черноглазый юнко Шуфельдта ( J.h.shufeldti )
- Темноглазый юнко Тербера ( J.h.thurberi )
- Темноглазый юнко Таунсенда ( Jh Townsendi )
У этих восьми подвидов черновато-серые головы и груди с коричневой спиной и крыльями, а также красноватыми боками, с тенденцией к более тусклому и бледному оперению во внутренних и южных частях ареала. [ 11 ] Орегонские темноглазые юнко также менее известны как темноглазые юнко с коричневой спиной. Это наиболее распространенная группа подвидов на Западе, гнездящаяся в хребтах Тихоокеанского побережья от юго-восточной Аляски до крайней северной Нижней Калифорнии и зимующая на Великих равнинах и в северной части Соноры . Существует нерешенный спор о том, является ли эта большая и обособленная группа подвидов на самом деле отдельным видом с восемью (или девятью, см. ниже) собственными подвидами. [ нужна ссылка ]

Розовобокая группа
[ редактировать ]- розовобокий темноглазый юнко ( J.h.mearnsi )
Иногда его считают девятым подвидом в группе орегонских/коричнево-спинных особей. Этот подвид имеет более светлую серую голову и грудь, чем восемь темноглазых юнко орегонских/коричнево-спинных особей, с контрастирующими темными окрасами . Спина и крылья коричневые. Он имеет розовато-коричный цвет, который более насыщенный и покрывает большую часть боков и груди, чем у восьми темноглазых юнко Орегона с коричневой спиной. Он гнездится в северных Скалистых горах от южной Альберты до восточного Айдахо и западного Вайоминга и зимует в центральном Айдахо и близлежащей Монтане, а также от юго-западной Южной Дакоты, южного Вайоминга и северной Юты до северной Соноры и Чиуауа . [ 11 ]

Группа седых
[ редактировать ]- Юнко седоголовый темноглазый ( J. h. caniceps )
Этот подвид по существу довольно светло-серый сверху с ржавой спинкой. Он гнездится в южных районах Скалистых гор от Колорадо до центральной Аризоны и Нью-Мексико, а зимует в северной Мексике. [ 12 ] [ 10 ]
Группа красных
[ редактировать ]- юнко красноспинный темноглазый ( J. h. dorsalis )

Юнко красноспинный темноглазый ( J. h. dorsalis )

Иногда включаемый в состав собственно седоголового темноглазого юнко в составе седой группы, этот подвид отличается от него более серебристым клювом. [ 11 ] с темной верхней челюстью и светлой нижней челюстью, [ 12 ] [ 10 ] переменное количество ржавчины на крыльях и бледная нижняя часть тела. Это роднит его с желтоглазым юнко ( Junco phaeonotus ), за исключением темных глаз. Встречается в южных горах Аризоны и Нью-Мексико. [ 10 ] По своему гнездовому ареалу он не пересекается с желтоглазым юнко.
Родственные виды
[ редактировать ]Чрезвычайно редкий Guadalupe junco ( Junco insularis ) ранее считался подвидом этого вида (либо включенным в группу седых головок, либо помещенным в седьмую собственную группу, группу Guadalupe ), но теперь рассматривается как отдельный вид. самостоятельный вид – возможно, довольно молодой, но, безусловно, эта популяция развивалась быстрее , чем 14 или 15 подвидов темноглазого юнко на материке, из-за небольшой численности популяции и эффекта основателя . [ 7 ]
Описание
[ редактировать ]Взрослые темноглазые юнко обычно имеют серые головы, шеи и груди, серые или коричневые спины и крылья, а также белый живот, но демонстрируют сбивающее с толку разнообразие деталей оперения. Белые внешние рулевые перья отчетливо сверкают в полете и во время прыжков по земле. Клюв обычно бледно-розоватый. [ 12 ]
Самцы, как правило, имеют более темные и заметные отметины, чем самки. Темноглазый юнко имеет длину от 13 до 17,5 см (от 5,1 до 6,9 дюйма) и размах крыльев от 18 до 25 см (от 7,1 до 9,8 дюйма). [ 12 ] [ 13 ] Масса тела может варьироваться от 18 до 30 г (от 0,63 до 1,06 унции). [ 12 ] Среди стандартных размеров хорда крыла составляет от 6,6 до 9,3 см (от 2,6 до 3,7 дюйма), хвост - от 6,1 до 7,3 см (от 2,4 до 2,9 дюйма), клюв - от 0,9 до 1,3 см (от 0,35 до 0,51 дюйма), а предплюсна - От 1,9 до 2,3 см (от 0,75 до 0,91 дюйма). [ 14 ] Молодые особи часто имеют бледные пестринки на нижней части тела, и их даже можно принять за вечерних воробьев ( Pooecetes gramineus ), пока в два-три месяца они не приобретут взрослое оперение, но головы темноглазых птенцов юнко, как правило, уже довольно однородны по цвету, и первоначально их клювы все еще имеют заметные желтоватые края на зеве - остатки мясистых сережек , которыми родители кормят птенцов.
Песня представляет собой трель, сходную с песней воробья-щебня ( Spizella passerina ), за исключением того, что песня красноспинного темноглазого юнко (см. выше ) более сложная, похожа на песню желтоглазого юнко ( Junco phaeonotus ). Крик также напоминает крик чернозобой синей славки ( Setophaga caerulescens ), представителя семейства камышевок Нового Света . [ 15 ] Звонки включают тикающие звуки и очень высокие звенящие чипсы . [ 10 ] Среди любителей пения птиц он известен как отличная птица для изучения «птичьего языка».

Распространение и среда обитания
[ редактировать ]Ареалом размножения темноглазого юнко являются участки хвойных или смешанных лесов по всей Северной Америке. В других оптимальных условиях он также использует другие среды обитания, но на южной окраине своего ареала он может сохраняться только в своей любимой среде обитания. [ 16 ] Северные птицы мигрируют дальше на юг, прибывают на зимовку в период с середины сентября по ноябрь и отправляются на размножение с середины марта, причем почти все они улетают примерно к концу апреля. [ 16 ] [ 17 ] Многие популяции являются постоянными жителями или высотными мигрантами, а в холодные годы они могут предпочесть остаться на зимнем ареале и размножаться там. [ 16 ] Например, в горах Сьерра-Невада в восточной Калифорнии популяции J. hyemalis мигрируют в зимние ареалы на 5 000–7 000 футов (1 500–2 100 м) ниже, чем их летний ареал. Сезонно-симпатичные самки демонстрируют разницу в миграции и сроках размножения, которые зависят от гормональной регуляции и регуляции яичников. [ 18 ] У самок-мигрантов J. hyemails рост гонад задерживается, что дает время для сезонной миграции. Затем они мигрируют на северо-восток США, где постоянным подвидом является каролинский темноглазый юнко ( J. h. carolinensis ). Самки темноглазых юнко Каролины имеют большие яичники и, следовательно, не испытывают задержки роста гонад, поскольку они проживают в этом районе. Зимой темноглазые юнко распространены в городах и их окрестностях, а во многих местах они являются наиболее распространенными птицами на кормушках. [ 12 ] Сланцевый темноглазый юнко ( J. h. hyemalis ) — редкий залет в Западную Европу и может успешно зимовать в Великобритании , обычно в домашних садах.
Поведение и экология
[ редактировать ]Эти птицы кормятся на земле. Зимой они часто кормятся стаями, которые могут содержать несколько разных подвидов. В основном они питаются семенами, а иногда и насекомыми. Известно, что стаю темноглазых юнко называют метелью. [ нужна ссылка ]
Разведение
[ редактировать ]Гнездятся темноглазые юнко обычно в чашеобразном углублении на земле, хорошо скрытом растительностью или другим материалом, хотя иногда гнезда встречаются и в нижних ветвях кустарника или дерева. Гнезда имеют внешний диаметр около 10 см (3,9 дюйма) и выстланы мелкой травой и волосами. Обычно две кладки в период размножения откладываются по четыре яйца. Слегка блестящие яйца сероватые или бледно-голубовато-белые, с сильными пятнами (иногда с пятнами) различных оттенков коричневого, фиолетового или серого. Пятна сосредоточены на большом конце яйца. Яйца насиживает самка в течение 12–13 дней. Птенцы покидают гнездо через 11–14 дней после вылупления. [ 19 ]
-
 Гнездо темноглазого юнко с яйцами
Гнездо темноглазого юнко с яйцами -
 Молодой розовобокий темноглазый юнко ( J. h. mearnsi ) примерно через месяц после вылупления, Йеллоустонский национальный парк.
Молодой розовобокий темноглазый юнко ( J. h. mearnsi ) примерно через месяц после вылупления, Йеллоустонский национальный парк.


Диета
[ редактировать ]Темноглазые юнко в основном питаются насекомыми и семенами , а также ягодами . [ 20 ]
Эволюция
[ редактировать ]Постледниковая теория и диверсификация
[ редактировать ]Темноглазые юнко широко исследовались как модель быстрого видообразования . Это происходит из-за исключительно высокого фенотипического разнообразия, о чем свидетельствует большое количество цветовых узоров, за, казалось бы, очень короткий промежуток времени. [ 21 ] Текущие оценки эволюции темноглазых юнко ( J. hyemalis ) относят диверсификацию от желтоглазых юнко ( J. phaeonotus ) 1800 лет назад, основываясь на мтДНК. [ 22 ] Одной из теорий, объясняющих причину этого расширения, является послеледниковая теория. Эта теория утверждает, что предковые популяции юнко распространились дальше на север по всей Северной Америке по мере таяния ледников. Отступающие ледники откроют множество новых мест обитания с новым селективным давлением . В этих условиях естественный отбор может очень сильно повлиять на популяции, поскольку существует множество открытых ниш . Таким образом, даже короткие периоды изоляции могут привести к дивергенции популяций. Постледниковая теория подтверждается тем, что желтоглазые и темноглазые юнко имеют общий доминантный гаплотип в митохондриальной ДНК, что указывает на недавний всплеск численности популяции. Желтоглазые юнко относительно репродуктивно изолированы . Таким образом, признак с большей вероятностью будет обнаружен у обоих видов из-за происхождения, чем из-за потока генов , поскольку вероятность скрещивания, происходящего достаточно часто, чтобы признак был настолько распространенным, гораздо менее вероятна, чем вероятность того, что он унаследован от одного предкового события. Ареал красноспинных ( J. h. dorsalis ) и седых юнко ( Дж. Х. caniceps ) на юге Северной Америки также предоставляет доказательства, поскольку эти два вида, по-видимому, представляют собой последовательные шаги в развитии темноглазых форм. Красноспинный юнко внешне очень похож на желтоглазого юнко. Здесь также находится самый южный ареал темноглазых видов юнко. Согласно постледниковой теории, эта популяция должна была быть старше остальных, поскольку популяция распространилась на север. Седоголовый юнко, встречающийся севернее, имеет такой же светлый клюв, как и остальные представители комплекса юнко. Вместе они показывают, что количество темноглазых юнко-подобных черт увеличивается по мере их продвижения на север. Если применить постледниковую теорию, северные подвиды юнко диверсифицировались бы позже. Это делает их широкую гамму окраски более заметной, поскольку она должна возникнуть еще быстрее.
Диверсификация группы орегонских юнко, вероятно, является результатом как генетического дрейфа , так и отбора. [ 21 ] Географически изолированные подвиды, такие как J. Townsendi и J. pontilis , имели низкую генетическую связь с окружающей средой. Это означало, что большая часть различий между ними не приносила измеримого улучшения физической формы . Вместо этого большинство различий, вероятно, было связано с изоляцией и небольшим размером популяции. [ 21 ] Это усилит влияние генетического дрейфа, увеличивая различия между подвидами. Барьеры изоляции, созданные неподходящей средой обитания в пустыне, вероятно, привели к отсутствию потока генов. Другие подвиды, такие как J. pinosis и J. thurberi , были гораздо менее географически изолированы. Их ареалы часто перекрываются, образуя зоны интерградации , где оба подвида существуют и скрещиваются. Вместо географической изоляции различия между этими подвидами, вероятно, обусловлены адаптацией . Это подтверждается тем, что популяции J. pinosis коррелируют с изотермией или с тем, насколько температура меняется от дня к ночи и от сезона к сезону, и отрицательно с высотой над уровнем моря . Это указывает на то, что они, вероятно, адаптировались к этим конкретным условиям. Репродуктивные барьеры могут возникнуть из-за этих адаптаций и различий в предпочтительной среде обитания, а не из-за отсутствия взаимодействия. Также наблюдался заметный поток генов между J. thurberi , J. shufeldti , J. montanus и J. oreganus . Эти виды образовали широтный градиент, где подвиды перетекают друг в друга и уменьшаются или увеличиваются в количестве в зависимости от широты. Дивергенция, вероятно, была вызвана местной адаптацией на пути на север. В целом, вполне вероятно, что не существует единого механизма, управляющего исторической диверсификацией темноглазых юнко. Разные подвиды с большей или меньшей частотой ощущают разные эволюционные силы. Однако совокупность множества сил приводит к расхождению всех или большинства подвидов. Вполне вероятно, что в случае с темноглазыми юнко эти многочисленные факторы способствовали ускорению перемен. Половой отбор и географическая изоляция укрепляют и усиливают небольшие изменения, возникшие в результате генетического дрейфа и естественного отбора. Точные механизмы быстрой диверсификации юнко остаются предметом дальнейших исследований.
Урбанизация
[ редактировать ]Популяции темноглазых юнко также представляют интерес для ученых из-за того, как они реагируют на урбанизацию . Во многом это моделируется на примере популяции темноглазых юнко, живущих в кампусе Калифорнийского университета в Сан-Диего . Впервые население было изучено в 1980-х годах, и оно стало предметом интереса из-за того, насколько оно отличается от менее городского населения соседних регионов. [ 23 ] Одним из наиболее заметных отличий популяции UCSD является то, что они не мигрируют, как другие популяции юнко штата Орегон. Вместо этого они остаются в кампусе круглый год. Это отличает их от других популяций юнко, которые мигрируют в кампус Калифорнийского университета в Сан-Франциско только зимой. На отсутствие миграции, вероятно, повлиял мягкий средиземноморский климат UCSD. Этот климат также приводит к более длительным сезонам размножения, чем в более высоких широтах. Это позволяет юнко UCSD иметь до 4 выводков в год, а не 1 или 2 в близлежащих популяциях. Большая численность выводка, в свою очередь, могла действовать как селективное давление в пользу участия родителей. Поскольку птицы размножаются больше в течение сезона, первоначальное спаривание менее важно, и самцы, участвующие в уходе, с большей вероятностью снова размножатся в том же сезоне. [ 24 ] Птицы, проживающие в UCSD, также чаще собираются парами, чем зимующие птицы, что, возможно, является результатом того же селективного давления.
Несмотря на то, что поблизости существуют другие популяции юнко, их популяции различаются гораздо сильнее, чем ожидалось. Степень разницы между объединенными компаниями UCSD и другими местными объединенными компаниями была ближе к той, которую можно было бы ожидать в условиях географической изоляции. Поскольку ближайшие популяции (расположенные в горах) относятся к подвиду J. thurberi , предполагалось, что птицы UCSD произошли от предковой популяции J. thurberi . Однако анализ генома показывает, что популяция, вероятно, возникла из прибрежного подвида J. pinosis 20-30 поколений назад, и это условия, которые делают эффект основателя весьма вероятным. [ 24 ] Этот вывод дополнительно подтверждается фенотипическим сходством: птицы UCSD и J. pinosis имеют одинаковую степень белого цвета хвоста и немигрирующего поведения. Это осознание подрывает идею о том, что урбанизация приводит к отсутствию миграции, но не о том, что она вызвала многие другие изменения. Дополнительные доказательства получены от птиц UCSD, время от времени скрещивающихся с зимующими особями J. pinosis , но не с J. thurberi . И это несмотря на то, что J. thurberi juncos гораздо более распространен в этом районе. Если бы популяция UCSD скрещивалась с обоими, это, скорее всего, уже наблюдалось бы. Поскольку популяция-основательница была довольно маленькой, а ближайшая популяция J. pinosis находилась далеко, генетический дрейф, вероятно, оказал большое влияние на развитие популяции. Однако была обнаружена сильная связь между переменными среды обитания и функциональными генами. Эта ассоциация является признаком естественного отбора и адаптивных особенностей, а не просто дрейфа. Это указывает на то, что генетический дрейф, вероятно, не единственная действующая эволюционная сила. Отбор, вероятно, усиливает изменения, первоначально вызванные дрейфом, что позволяет популяции UCSD очень быстро расходиться со своими соседями.
Дополнительные доказательства наличия адаптивных генов в популяции UCSD можно найти на уровне генов. Два наиболее дифференцированных гена между предковой популяцией J. pinosis и птицами UCSD были связаны с полезными чертами для городской среды. [ 23 ] В частности, ABCB6 позволил птицам лучше переносить тяжелые металлы. Поскольку отравление тяжелыми металлами является документально подтвержденной проблемой для городских птиц, этот ген, вероятно, повышает приспособленность к жизни в городских условиях. KCNQ4 связан с высокочастотной эхолокацией у летучих мышей и, по-видимому, коррелирует с более высокими звуками. Это полезно в городских условиях, поскольку низкочастотный городской шум часто заглушает крики птиц. Благодаря более высоким звукам их голоса с меньшей вероятностью затеряются в этом шуме. Учитывая теоретические преимущества выживания, распространенность этих черт, вероятно, обусловлена естественным отбором. Еще предстоит провести исследование того, какие гены в популяции UCSD выполняют адаптивную функцию, а какие, вероятно, являются лишь следствием дрейфа. Многие ранее существовавшие исследования могут оказаться менее эффективными, особенно после открытия того, что они более тесно связаны с J. pinosis . Тем не менее, они по-прежнему являются очень многообещающей модельной системой того, как птицы адаптируются к городским экосистемам.
Ссылки
[ редактировать ]- ^ БердЛайф Интернэшнл. (2021). « Юнко гиемалис » . Красный список исчезающих видов МСОП . 2021 : e.T22721097A138466281. doi : 10.2305/IUCN.UK.2021-3.RLTS.T22721097A138466281.en . Проверено 3 июля 2024 г.
- ^ Линней, Карл (1758). Система природы по трем царствам природы по классам, отрядам, родам, видам, с признаками, различиями, синонимами, местами (на латыни). Том. 1 (10-е изд.). Холмии (Стокгольм): Лоуренс Сальвиус. п. 183.
- ^ Перейти обратно: а б Кэтсби, Марк (1729–1732). Естественная история Каролины, Флориды и Багамских островов (на английском и французском языках). Том. 1. Лондон: У. Иннис и Р. Мэнби. п. 36, Табличка 36.
- ^ Комитет по классификации и номенклатуре (1931 г.). Контрольный список птиц Северной Америки (4-е изд.). Ланкастер, Пенсильвания: Союз американских орнитологов. п. 345.
- ^ Пейнтер, Раймонд А. младший, изд. (1970). Контрольный список птиц мира . Том. 13. Кембридж, Массачусетс: Музей сравнительной зоологии. п. 63.
- ^ Ваглер, Иоганн Георг (1831). «Некоторые заметки о животных Мексики» . Исида Окенская (на немецком и латыни). 1831 год . Кол. 510–535 [526].
- ^ Перейти обратно: а б с Джилл, Фрэнк ; Донскер, Дэвид; Расмуссен, Памела , ред. (январь 2022 г.). «Воробьи Нового Света, Буш-Танагеры» . Всемирный список птиц МОК, версия 12.1 . Международный союз орнитологов . Проверено 4 февраля 2022 г.
- ^ «Юнко» . Оксфордский словарь английского языка (онлайн-изд.). Издательство Оксфордского университета . (Требуется подписка или членство участвующей организации .)
- ^ Джоблинг, Джеймс А. (2010). Словарь научных названий птиц Хелма . Лондон, Великобритания: Кристофер Хелм. стр. 197 , 212. ISBN. 978-1-4081-2501-4 .
- ^ Перейти обратно: а б с д и ж г Сибли, Дэвид Аллен (2000): Путеводитель Сибли по птицам . Альфред А. Кнопф, Нью-Йорк, стр. 500–502, ISBN 0-679-45122-6
- ^ Перейти обратно: а б с д Данн, Джон Л. (2002). «Идентификация розовобоких юнко с предостережениями относительно вариаций оперения и гибридизации». Птичье наблюдение . 34 (5): 432–443.
- ^ Перейти обратно: а б с д и ж г Корнельская лаборатория орнитологии (2002): Путеводитель по птицам – Темноглазый юнко . Проверено 20 января 2007 г.
- ^ Rising, JD (2010) Руководство по идентификации и естественной истории воробьев в Соединенных Штатах и Канаде . Издательство Christopher Helm, Лондон, ISBN 1-4081-3460-8 .
- ^ Воробьи и овсянки: Путеводитель по воробьям и овсянкам Северной Америки и мира Клайва Байерса и Урбана Олссона. Хоутон Миффлин (1995). ISBN 978-0-395-73873-3 .
- ^ «Черногорлая славка ( Dendroica caerulescens )» . Птицы в лесных ландшафтах . Корнеллская лаборатория орнитологии.
- ^ Перейти обратно: а б с Орнитологическое общество Огайо (2004 г.): Аннотированный контрольный список штата Огайо. Архивировано 18 июля 2004 г., в Wayback Machine.
- ^ Хеннингер, ВФ (1906). «Предварительный список птиц округа Сенека, штат Огайо» (PDF) . Бюллетень Уилсона . 18 (2): 47–60.
- ^ Киммитт, Эбигейл А.; Хардман, Джек В.; Стрикер, Крейг А.; Кеттерсон, Эллен Д. (сентябрь 2019 г.). Сокман, Кейт (ред.). «Миграционная стратегия объясняет различия во времени репродуктивного развития самок у сезонно симпатичных певчих птиц» . Функциональная экология . 33 (9): 1651–1662. Бибкод : 2019FuEco..33.1651K . дои : 10.1111/1365-2435.13386 . ISSN 0269-8463 .
- ^ «Жизнеописание черноглазого юнко» . Все о птицах . Корнельская лаборатория орнитологии. 2019 . Проверено 21 октября 2021 г.
- ^ «Темноглазый Юнко | Полевой гид Одюбона» . www.audubon.org . Проверено 20 апреля 2024 г.
- ^ Перейти обратно: а б с Фриис, Гильермо; Фандос, Гильермо; Зеллмер, Аманда Дж.; МакКормак, Джон Э.; Фэрклот, Брант К.; Мила, Борха (декабрь 2018 г.). «Общегеномные сигналы дрейфа и местной адаптации во время быстрого расхождения линий у певчих птиц». Молекулярная экология . 27 (24): 5137–5153. Бибкод : 2018MolEc..27.5137F . дои : 10.1111/mec.14946 . ISSN 1365-294X . ПМИД 30451354 .
- ^ Мила, Борха; МакКормак, Джон Э; Кастаньеда, Габриэла; Уэйн, Роберт К.; Смит, Томас Б. (07 ноября 2007 г.). «Недавнее расширение ареала послеледникового периода привело к быстрой диверсификации линии певчих птиц рода Junco» . Труды Королевского общества B: Биологические науки . 274 (1626): 2653–2660. дои : 10.1098/rspb.2007.0852 . ISSN 0962-8452 . ПМК 2279216 . ПМИД 17725978 .
- ^ Перейти обратно: а б Раснер, Калифорния; Ага, П.; Эггерт, Л.С.; Хант, Кентукки; Вудрафф, Д.С.; Прайс, ТД (6 февраля 2004 г.). «Генетическая и морфологическая эволюция после события-основателя темноглазого юнко, Junco hyemalis thurberi» . Молекулярная экология . 13 (3): 671–681. Бибкод : 2004MolEc..13..671R . дои : 10.1046/j.1365-294X.2004.02104.x . ПМИД 14871370 .
- ^ Перейти обратно: а б Фриис, Гильермо; Этвелл, Джонатан В.; Фудикар, Адам М.; Грейвс, Тимоти Дж.; Да, Памела Дж.; Прайс, Тревор Д.; Кеттерсон, Эллен Д.; Мила, Борха (май 2022 г.). «Быстрая эволюционная дивергенция популяции певчих птиц после недавней колонизации городской территории» . Молекулярная экология . 31 (9): 2625–2643. Бибкод : 2022MolEc..31.2625F . дои : 10.1111/mec.16422 . ISSN 0962-1083 . ПМИД 35253305 .
Внешние ссылки
[ редактировать ]- Идентификация темноглазого юнко, включая звук и видео , в Корнельской орнитологической лаборатории.
- Темноглазый юнко — Junco hyemalis. Архивировано 17 апреля 2011 г. в Wayback Machine — Информационный центр по идентификации птиц USGS Patuxent.
- Юнкос: Что мы знаем? - Экспертное обсуждение атипичных особей, тонкостей подвидовой идентификации и правильного понимания популяции цисмонтана из списка рассылки ID-Frontiers (январь 2004 г.), дополненное фотографиями и рисунками.
- Изучите виды: темноглазый Юнко на eBird (Корнельская лаборатория орнитологии)
- Фотогалерея черноглазых юнко в VIREO (Университет Дрекселя)
- Виды, находящиеся в Красном списке МСОП, вызывающие наименьшее беспокойство
- Птицы, описанные в 1758 году.
- Птицы Аппалачей (США)
- Птицы Канады
- Птицы Сьерра-Невады (США)
- Фауна калифорнийского чапараля и лесных массивов
- Фауна района залива Сан-Франциско
- Торопиться
- Местные птицы Аляски
- Местные птицы северо-востока США.
- Местные птицы Скалистых гор.
- Местные птицы запада США
- Таксоны, названные Карлом Линнеем