R-петля
R -петля представляет собой трехцепочечную структуру нуклеиновой кислоты, состоящую из гибрида ДНК: РНК и связанной с ней нематрицной одноцепочечной ДНК . R-петли могут образовываться при различных обстоятельствах и могут переноситься или очищаться клеточными компонентами. Термин «R-петля» был дан, чтобы отразить сходство этих структур с D-петлями ; «R» в данном случае означает участие фрагмента РНК .
В лаборатории R-петли могут быть созданы путем транскрипции последовательностей ДНК (например, тех, которые имеют высокое содержание GC), которые способствуют отжигу РНК позади прогрессирующей РНК-полимеразы. [ 1 ] Для формирования стабильной структуры R-петли требуется не менее 100 пар оснований гибрида ДНК:РНК. R-петли также могут быть созданы путем гибридизации зрелой мРНК с двухцепочечной ДНК в условиях, благоприятствующих образованию гибрида ДНК-РНК; в этом случае области интрона (которые были вырезаны из мРНК) образуют одноцепочечные петли ДНК, поскольку они не могут гибридизоваться с комплементарной последовательностью мРНК. [ 2 ]
История
[ редактировать ]
R-петля была впервые описана в 1976 году. [ 3 ] Независимые исследования R-петли, проведенные в лабораториях Ричарда Дж. Робертса и Филиппа А. Шарпа, показали, что белки, кодирующие аденовируса, гены содержат последовательности ДНК, которых нет в зрелой мРНК. [ 4 ] [ 5 ] Робертс и Шарп были удостоены Нобелевской премии в 1993 году за независимое открытие интронов. После открытия в аденовирусе интроны были обнаружены в ряде эукариотических генов, таких как эукариотический ген овальбумина (сначала лабораторией О'Мэлли, затем подтвержденный другими группами), [ 6 ] [ 7 ] гексонная ДНК, [ 4 ] и внехромосомные рРНК гены Tetrahymena thermophila . [ 8 ]
В середине 1980-х годов разработка антитела , которое специфически связывается со структурой R-петли, открыла двери для иммунофлуоресцентных исследований, а также полногеномной характеристики образования R-петли с помощью DRIP-seq . [ 9 ]
Отображение R-петли
[ редактировать ]Картирование R-петли — это лабораторный метод, используемый для отличия интронов от экзонов в двухцепочечной ДНК. [ 10 ] Эти R-петли визуализируются с помощью электронной микроскопии и выявляют интронные области ДНК, создавая несвязанные петли в этих областях. [ 11 ]
R-петли in vivo
[ редактировать ]Возможность использования R-петлей в качестве праймеров репликации была продемонстрирована в 1980 году. [ 12 ] В 1994 году было продемонстрировано присутствие R-петлей in vivo посредством анализа плазмид, выделенных из мутантов E. coli , несущих мутации топоизомеразы . [ 13 ] Это открытие эндогенных R-петлей в сочетании с быстрым развитием технологий генетического секвенирования вдохновило на расцвет исследований R-петли в начале 2000-х годов, который продолжается и по сей день. [ 14 ]
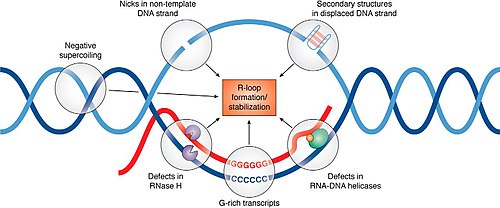
Регуляция формирования и разрешения R-петли
[ редактировать ]Более 50 белков, по-видимому, влияют на накопление R-петли, и хотя считается, что многие из них способствуют секвестрации или обработке вновь транскрибируемой РНК для предотвращения повторного отжига с матрицей, механизмы взаимодействия R-петли для многих из этих белков остаются предстоит определить. [ 15 ]
Существует три основных класса ферментов, которые могут удалять РНК, попавшую в дуплекс внутри R-петли. Ферменты RNaseH являются основными белками, ответственными за растворение R-петлей, разрушая фрагмент РНК, позволяя двум комплементарным нитям ДНК отжигаться. [ 16 ] Альтернативно, хеликазы раскручивают дуплекс РНК:ДНК и высвобождают РНК. Сенатаксин является одной из геликаз, которая может перемещаться по оцРНК и, по-видимому, необходима для предотвращения образования R-петли в сайтах паузы транскрипции. [ 17 ] Третий класс ферментов, способный удалять R-петли, представляет собой транслоказы точки ветвления, такие как FANCM , SMARCAL1 и ZRANB3 у людей или RecG у бактерий. [ 1 ] Транслоказы точки ветвления действуют на двухцепочечную ДНК, прилегающую к гибриду ДНК:РНК. Нажимая на точку ветвления, они «застегивают» ДНК и удаляют застрявшую РНК. Это делает транслоказы точки ветвления эффективными при удалении как РНК, так и белков, связанных со структурой R-петли. Транслоказы точек ветвления могут работать вместе с РНКазой H и хеликазами на некоторых типах R-петлей, которые встречаются в сложных структурах.
Роль R-петлей в генетической регуляции
[ редактировать ]Формирование R-петли является ключевым этапом переключения класса иммуноглобулина , процесса, который позволяет активированным В-клеткам модулировать выработку антител . [ 18 ] Они также, по-видимому, играют роль в защите некоторых активных промоторов от метилирования . [ 19 ] Наличие R-петлей также может ингибировать транскрипцию. [ 20 ] Кроме того, образование R-петли, по-видимому, связано с «открытым» хроматином , характерным для активно транскрибируемых областей. [ 21 ] [ 22 ]
R-петли как генетическое повреждение
[ редактировать ]Когда образуются незапланированные R-петли, они могут вызвать повреждение по ряду различных механизмов. [ 23 ] Открытая одноцепочечная ДНК может подвергаться атаке эндогенных мутагенов, включая ферменты, модифицирующие ДНК, такие как цитидиндезаминаза, индуцируемая активацией , и может блокировать репликационные вилки, вызывая коллапс вилки и последующие двухцепочечные разрывы. [ 24 ] Кроме того, R-петли могут вызывать незапланированную репликацию, действуя как праймер . [ 12 ] [ 22 ]
Накопление R-петли связано с рядом заболеваний, включая боковой амиотрофический склероз 4-го типа (АЛС4) , атаксию глазодвигательной апраксии 2-го типа (АОА2) , синдром Айкарди-Гутьера , синдром Ангельмана , синдром Прадера-Вилли и рак. [ 14 ] Гены, связанные с анемией Фанкони, также, по-видимому, важны для поддержания стабильности генома в условиях накопления R-петлей. [ 25 ]
R-петли, интроны и повреждение ДНК
[ редактировать ]Интроны — это некодирующие области внутри генов , которые транскрибируются вместе с кодирующими областями генов, но впоследствии удаляются из первичного транскрипта РНК путем сплайсинга . Активно транскрибируемые участки ДНК часто образуют R-петли, уязвимые для повреждения ДНК . Интроны уменьшают образование R-петли и повреждение ДНК в высокоэкспрессируемых генах дрожжей. [ 26 ] Полногеномный анализ показал, что интронсодержащие гены демонстрируют пониженные уровни R-петли и меньшее повреждение ДНК по сравнению с безинтронными генами с аналогичной экспрессией как у дрожжей, так и у людей. [ 26 ] Вставка интрона в ген, склонный к R-петле, также может подавлять образование и рекомбинацию R-петли . Боннет и др. (2017) [ 26 ] предположили, что функция интронов по поддержанию генетической стабильности может объяснить их эволюционное сохранение в определенных местах, особенно в генах с высокой экспрессией.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Ходсон, Шарлотта; ван Твест, Сильви; Дылевска, Малгожата; О'Рурк, Жюльен Дж.; Тан, Винни; Мерфи, Винсент Дж.; Валия, Манну; Абуш, Лара; Неминущи, Ядвига; Данн, Элиз; Байтелл-Дуглас, Рохан; Хейерхорст, Джордж; Недзведз, Войцех; Динс, Эндрю Дж. (2022). «Транслокация точки ветвления ремоделировщиками вилок как общий механизм удаления R-петли» . о ячейках Отчеты 41 10):111749.doi : ( 10.1016/j.cellrep.2022.111749 .
- ^ Ван, Хунхун; Ли, Чжинанг; Ли, Сян, Ин; 02-19 ) . Достижения науки .7 / (8). : 2021SciA .... doi : 10.1126 sciadv.abe3516 . ISSN 2375-2548 . PMC 7888926 . 7.3516W . Бибкод
- ^ Томас М., Уайт Р.Л., Дэвис Р.В. (июль 1976 г.). «Гибридизация РНК с двухцепочечной ДНК: образование R-петли» . Труды Национальной академии наук Соединенных Штатов Америки . 73 (7): 2294–8. Бибкод : 1976PNAS...73.2294T . дои : 10.1073/pnas.73.7.2294 . ПМК 430535 . ПМИД 781674 .
- ^ Jump up to: а б Бергет С.М., Мур С., Шарп П.А. (август 1977 г.). «Сплайсированные сегменты на 5'-конце поздней мРНК аденовируса 2» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (8): 3171–5. Бибкод : 1977PNAS...74.3171B . дои : 10.1073/pnas.74.8.3171 . ПМЦ 431482 . ПМИД 269380 .
- ^ Чоу Л.Т., Гелинас Р.Э., Брокер Т.Р., Робертс Р.Дж. (сентябрь 1977 г.). «Удивительное расположение последовательностей на 5'-концах информационной РНК аденовируса 2». Клетка . 12 (1): 1–8. дои : 10.1016/0092-8674(77)90180-5 . ПМИД 902310 . S2CID 2099968 .
- ^ Лай Э.К., Ву С.Л., Дугайчик А., Каттералл Дж.Ф., О'Мэлли Б.В. (май 1978 г.). «Ген овальбумина: структурные последовательности в нативной куриной ДНК не являются смежными» . Труды Национальной академии наук Соединенных Штатов Америки . 75 (5): 2205–9. Бибкод : 1978PNAS...75.2205L . дои : 10.1073/pnas.75.5.2205 . ПМК 392520 . ПМИД 276861 .
- ^ О'Хара К., Бренак Р., Бенуа К., Шамбон П. (сентябрь 1979 г.). «Не более семи прерываний в гене овальбумина: сравнение геномной и двухцепочечной последовательностей кДНК» . Исследования нуклеиновых кислот . 7 (2): 321–34. дои : 10.1093/нар/7.2.321 . ПМК 328020 . ПМИД 493147 .
- ^ Чехия, Рио, округ Колумбия (октябрь 1979 г.). «Локализация транскрибируемых областей на генах внехромосомной рибосомальной РНК Tetrahymena thermophila путем картирования R-петли» . Труды Национальной академии наук Соединенных Штатов Америки . 76 (10): 5051–5. Бибкод : 1979PNAS...76.5051C . дои : 10.1073/pnas.76.10.5051 . ПМК 413077 . ПМИД 291921 .
- ^ Богуславски С.Дж., Смит Д.Е., Михалак М.А., Микельсон К.Е., Йеле К.О., Паттерсон В.Л., Каррико Р.Дж. (май 1986 г.). «Характеристика моноклональных антител к ДНК.РНК и их применение для иммунодетекции гибридов». Журнал иммунологических методов . 89 (1): 123–30. дои : 10.1016/0022-1759(86)90040-2 . ПМИД 2422282 .
- ^ Вулфорд Дж. Л., Росбаш М. (июнь 1979 г.). «Использование R-петли для структурной идентификации генов и очистки мРНК» . Исследования нуклеиновых кислот . 6 (7): 2483–97. дои : 10.1093/нар/6.7.2483 . ПМК 327867 . ПМИД 379820 .
- ^ Кинг RC, Стэнсфилд WD, Маллиган ПК (2007). Генетический словарь . Издательство Оксфордского университета 7.
- ^ Jump up to: а б Ито Т., Томизава Дж. (май 1980 г.). «Формирование РНК-праймера для инициации репликации ДНК ColE1 рибонуклеазой H» . Труды Национальной академии наук Соединенных Штатов Америки . 77 (5): 2450–4. Бибкод : 1980PNAS...77.2450I . дои : 10.1073/pnas.77.5.2450 . ПМЦ 349417 . ПМИД 6156450 .
- ^ Дроле М, Би Х, Лю Л.Ф. (январь 1994 г.). «Гипернегативная суперспирализация матрицы ДНК во время элонгации транскрипции in vitro» . Журнал биологической химии . 269 (3): 2068–74. дои : 10.1016/S0021-9258(17)42136-3 . ПМИД 8294458 .
- ^ Jump up to: а б Гро М., Громак Н. (сентябрь 2014 г.). «Разбалансировка: R-петли в болезнях человека» . ПЛОС Генетика . 10 (9): e1004630. дои : 10.1371/journal.pgen.1004630 . ПМК 4169248 . ПМИД 25233079 .
- ^ Чан Ю.А., Аристисабал М.Дж., Лу П.Ю., Луо З., Хамза А., Кобор М.С., Стирлинг ПК, Хитер П. (апрель 2014 г.). «Полногеномное профилирование участков, склонных к гибридизации ДНК:РНК дрожжей, с помощью DRIP-чипа» . ПЛОС Генетика . 10 (4): e1004288. дои : 10.1371/journal.pgen.1004288 . ПМЦ 3990523 . ПМИД 24743342 .
- ^ Серрителли С.М., Крауч Р.Дж. (март 2009 г.). «Рибонуклеаза H: ферменты эукариот» . Журнал ФЭБС . 276 (6): 1494–505. дои : 10.1111/j.1742-4658.2009.06908.x . ПМК 2746905 . ПМИД 19228196 .
- ^ Скурти-Статаки, Константина; Праудфут, Николас Дж.; Громак, Наталья (2011). «Человеческий сенатаксин растворяет гибриды РНК/ДНК, образующиеся в местах транскрипционной паузы, чтобы способствовать Xrn2-зависимому терминации» . Молекулярная клетка . 42 (6): 794–805. doi : 10.1016/j.molcel.2011.04.026 . ПМК 3145960 . ПМИД 21700224 .
- ^ Рой Д., Ю.К., Либер М.Р. (январь 2008 г.). «Механизм образования R-петли в последовательностях переключения класса иммуноглобулина» . Молекулярная и клеточная биология . 28 (1): 50–60. дои : 10.1128/mcb.01251-07 . ПМК 2223306 . ПМИД 17954560 .
- ^ Джинно П.А., Лотт П.Л., Кристенсен Х.К., Корф И., Шедин Ф. (март 2012 г.). «Формирование R-петли является отличительной характеристикой неметилированных промоторов CpG-островков человека» . Молекулярная клетка . 45 (6): 814–25. doi : 10.1016/j.molcel.2012.01.017 . ПМК 3319272 . ПМИД 22387027 .
- ^ Д'Суза А.Д., Белоцерковский Б.П., Ханавальт П.С. (февраль 2018 г.). «Новый способ ингибирования транскрипции, опосредованный PNA-индуцированными R-петлями, с помощью модельной системы in vitro» . Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1861 (2): 158–166. дои : 10.1016/j.bbagrm.2017.12.008 . ПМК 5820110 . ПМИД 29357316 .
- ^ Кастеллано-Посо М., Сантос-Перейра Х.М., Рондон А.Г., Баррозу С., Андухар Э., Перес-Алегри М., Гарсиа-Мусе Т., Агилера А. (ноябрь 2013 г.). «Петли R связаны с фосфорилированием гистона H3 S10 и конденсацией хроматина» . Молекулярная клетка . 52 (4): 583–90. дои : 10.1016/j.molcel.2013.10.006 . ПМИД 24211264 .
- ^ Jump up to: а б Константино Л., Кошланд Д. (июнь 2015 г.). «Инь и Ян биологии R-петли» . Современное мнение в области клеточной биологии . 34 : 39–45. дои : 10.1016/j.ceb.2015.04.008 . ПМЦ 4522345 . ПМИД 25938907 .
- ^ Белоцерковский Б.П., Торналетти С., Д'Суза А.Д., Ханавальт ПК (ноябрь 2018 г.). «Генерация R-петли во время транскрипции: формирование, обработка и клеточные результаты» . Восстановление ДНК . 71 : 69–81. дои : 10.1016/j.dnarep.2018.08.009 . ПМК 6340742 . ПМИД 30190235 .
- ^ Сольер Дж., Цимприч К.А. (сентябрь 2015 г.). «Во все тяжкие: R-петли и целостность генома» . Тенденции в клеточной биологии . 25 (9): 514–22. дои : 10.1016/j.tcb.2015.05.003 . ПМК 4554970 . ПМИД 26045257 .
- ^ Шваб, Ребекка А.; Неминущи, Ядвига; Шах, Фенил; Лэнгтон, Джейми; Лопес Мартинес, Дэвид; Лян, Чи-Чао; Кон, Мартин А.; Гиббонс, Ричард Дж.; Динс, Эндрю Дж.; Недзведз, Войцех (2015). «Путь анемии Фанкони поддерживает стабильность генома путем координации репликации и транскрипции» . Молекулярная клетка . 60 (3): 351–361. doi : 10.1016/j.molcel.2015.09.012 . ПМЦ 4644232 . ПМИД 26593718 .
- ^ Jump up to: а б с Бонне А, Гроссо АР, Элькаутари А, Колено Е, Пресле А, Шридхара СК, Джанбон Г, Гели В, де Алмейда СФ, Паланкаде Б (август 2017 г.). «Интроны защищают геномы эукариот от генетической нестабильности, связанной с транскрипцией» . Молекулярная клетка . 67 (4): 608–621.e6. doi : 10.1016/j.molcel.2017.07.002 . ПМИД 28757210 .