Лепидодендралы
| Лепидодендралы Временной диапазон:
| |
|---|---|

| |
| Раздвоение в Лепидофлоиосе | |
| Научная классификация | |
| Королевство: | Растения |
| Клэйд : | Трахеофиты |
| Клэйд : | Ликофиты |
| Сорт: | Ликоподиопсида |
| Заказ: | † Лепидодендралы |
| Роды | |
| |
Lepidodendrales (от греческого «чешуйчатое дерево») или древесные плауны — вымерший отряд примитивных, сосудистых, разноспоровых , древесных ( древесных ) растений, принадлежащих к Lycopodiopsida . Из ископаемых ликопсид лучше всего изучены представители Lepidodendrales из-за огромного разнообразия образцов Lepidodendrales и разнообразия, в котором они сохранились; Широкое распространение экземпляров Lepidodendrales, а также их хорошая сохранность дают палеоботаникам исключительно подробные знания о репродуктивной биологии угольных болотных гигантов, вегетативном развитии и роли в их палеоэкосистеме. Определяющими характеристиками Lepidodendrales являются их вторичная ксилема , обширное развитие перидермы , трехзонная кора , корнеподобные придатки, известные как стигматические корешки, расположенные по спирали, и мегаспорангий, каждый из которых содержит одну функциональную мегаспору, которая прорастает внутри спорангия . Многим из этих различных органов растений были присвоены как общие, так и конкретные названия, поскольку относительно немногие из них были обнаружены органически связанными друг с другом. Было обнаружено несколько экземпляров, высота которых достигает 40. [ 1 ] и даже 50 метров [ 2 ] и диаметром более 2 метров у основания. Массивные стволы некоторых видов обильно разветвлялись, образуя большие кроны из облиственных ветвей; хотя некоторые листья были длиной до 1 метра, большинство из них были намного короче, и когда листья падали с ветвей, их заметные основания листьев оставались на поверхности ветвей. Стробилы можно найти на кончиках дистальных ветвей или в верхней части основного ствола. Подземные органы Lepidodendrales обычно состояли из дихотомических осей, несущих спирально расположенные боковые придатки, выполняющие функцию, эквивалентную корням. Иногда называемые «гигантскими плаунами», они, как полагают, более тесно связаны с современными иглами по ксилеме, хотя ископаемые экземпляры вымерших Selaginellales из позднего карбона также имели вторичную ксилему. [ 3 ] [ 4 ]
Морфология
[ редактировать ]Lepidodendrales имели высокие, толстые стволы, редко ветвящиеся и увенчанные кроной из раздвоенных ветвей с пучками листьев . Эти листья были длинными и узкими, похожими на большие травинки, и располагались спирально. Сосудистая система прямостоячего ствола необычна тем, что по мере роста растения меняет свое морфологическое развитие. Молодой ствол начинался как протостела , в которой сначала созревала внешняя ксилема (экзарх), но более поздняя и более высокая часть ствола развивалась как эктофлоическая сифоностела , в которой ксилема была окружена тканью флоэмы как с внутренней, так и с внешней стороны. [ 5 ]
Структура стебля
[ редактировать ]
Наиболее распространенными ископаемыми экземплярами Lepidodendrales, а также наиболее узнаваемыми являются сжатые поверхности стеблей, отмеченные постоянными, хотя и частично асимметричными, ромбовидными листовыми подушками. Эти окаменелости очень похожи на следы шин или кожу аллигатора , отсюда и греческое название «Lepidodendrales», что означает «чешуйчатые деревья». Эти листовые подушки на самом деле представляют собой расширенное основание листа, которое осталось после того, как листья опали, поскольку опадение листа не происходило вровень с поверхностью стебля. Ромбовидная форма возникает из-за острого угла верхней и нижней частей подушек, известных как листовая валик, и закругленного угла боковых сторон. Фактический рубец от листа находится немного выше середины подушки и имеет примерно эллиптическую форму. На рубце листа иногда можно обнаружить три небольших ямчатых отпечатка. Центральная и всегда присутствующая ямка возникает в результате сосудистого пучка, простирающегося в лист от стебля, известного как «парихнос» — системы аэрирующих тканей. Два других канала parichnos можно найти на Поверхности стебля лепидодендрона , хотя они не встречаются у Diaphorodendraceae. Над листовым рубцом имеется след от бывшего язычка . [ 6 ] покрывала Восковая кутикула поверхность стебля, включая листовые подушки, но не включала рубцы на стебле. [ 7 ] В простом эпидермисе отсутствуют специализированные клетки, такие как трихомы или эпидермальные железы. Устьица частые, затонувшие в неглубоких углублениях. [ 8 ]
Стебли Lepidodendrales могут быть протостелическими , как у Diaphorodendron , иметь смешанную сердцевину или быть сифоностелическими , как у Diaphorodendron. [ нужны разъяснения ] и Лепидодендрон . В стеблях со смешанной сердцевиной клетки паренхимы разбросаны, а трахеиды расположены посередине, хотя трахеиды имеют короткую, приземистую паренхиматозную форму; это приводится в качестве доказательства того, что сердцевина Lepidodendrales возникла из незрелых паренхиматозных клеток, которые не смогли должным образом дифференцироваться в трахеиды. Вокруг первичной ксилемы Lepidodendrales может располагаться вторичная ксилема, толщина которой может достигать нескольких сантиметров. В отличие от современных древесных деревьев, вторичная ксилема Lepidodendrales составляет лишь небольшую часть диаметра стебля, поскольку за большие стволы отвечает хорошо развитая перидерма. Первичные и вторичные трахеиды ксилемы имеют лестничную форму и имеют полоски Вильямсона или фимбриллы между этими лестничными линиями. Фимбриллы характеризуют древесину древесных ликопсид, хотя аналогичные структуры встречаются у современных буланых и колючих мхов, и эти фимбриллы представляют собой общую структуру для всех ликопсид. [ 9 ] С внешней стороны вторичной ксилемы граничит участок клеток с тонкими стенками, представляющий сосудистый камбий . Хотя современные семенные растения имеют двусторонний камбий, у лепидодендридов камбий односторонний, образующий вторичную ксилему только на внутренней стороне. [ 10 ] Зона флоэмы . отделена от этой вторичной ксилемы участком тонкостенных клеток, известным как «паренхимная оболочка» Имеющиеся данные свидетельствуют о том, что у древесных ликопсид не было вторичной флоэмы. Кора лепидодендридов обычно состояла из внутренней , средней и внешней коры, различающихся типами клеток. Внутренняя кора самая узкая и состоит из мелких клеток паренхимы; секреторные клетки, лакуны и различные склеротические клетки В этом разделе также можно найти . Средняя кора крупнее и, в свою очередь, состоит из более крупных клеток паренхимы. Для этого отдела характерны радиально простирающиеся лакуны в молодых стеблях, тогда как в более старых стеблях средняя кора обычно не сохраняется, за исключением нескольких клеток паренхимы. Внешняя кора не имеет определенного устройства, но ее клетки имеют несколько более толстые стенки. Перидерма образуется во внешней коре головного мозга. Перидерма у Diaphorodendron двузонная, внутренняя зона состоит из чередующихся толсто- и тонкостенных клеток, а наружная зона содержит темные, «смолистые» клетки. У лепидодендрона однородная или бизонатная перидерма массивная. [ 6 ] Рыхлая конструкция коры и большое количество тонкостенной перидермы способствовали отслаиванию слоев ткани в процессе окаменения. Это привело к появлению множества декортированных окаменелостей, которые часто предположительно имели внешние особенности стебля и ствола, но не имели листовых подушек и других особенностей. Декортикированным экземплярам были даны различные родовые названия, в том числе Кноррия — название стеблей, у которых почти все ткани за пределами ксилемы отсутствуют. [ 11 ]
Характер роста стебля Lepidodendrales можно реконструировать, анализируя закономерности роста их коры. У незрелых растений кора обширная, а внешняя поверхность стебля покрыта множеством рядов оснований листьев. По мере того как дерево продолжает расти, вторичная ксилема и перидерма происходят из сосудистого камбия и феллогена . Это увеличение ткани стебля и диаметра стебля приводит к отслаиванию внешних тканей, включая основания листьев; следовательно, на более старых участках растения внешняя поверхность ствола защищена перидермой. На многих старых рисунках лепидодендрона неправильно изображены основания листьев старых деревьев, доходящие до земли. На более высоких, более молодых уровнях дерева на ветвях меньше рядов более мелких листьев. В этих участках образуется меньше вторичной ксилемы и перидермы. Это уменьшение размера стелы и образования вторичных тканей продолжает сужаться к наиболее дистальным ветвям, где существует только крошечная протостела, нет вторичных тканей и мало рядов листьев; эта дистальная стадия развития известна как «апоксогенез». Эти маленькие дистальные веточки не могут со временем превратиться в более крупные ветви. Такой тип роста известен как детерминантный рост; это контрастирует с современным неопределенный характер роста большинства современных древесных растений. [ 12 ]
Структура листа
[ редактировать ]
Листья растений Lepidodendrales линейные, длиной около 1–2 м (3 фута 3 дюйма – 6 футов 7 дюймов). Стебли с наибольшим диаметром имеют самые длинные листья, что коррелирует с определенным ростом растений. [ 13 ] Многие органные таксоны, установленные для отдельных листьев Lepidodendrales, вероятно, произошли от одного и того же вида растений и различаются по морфологии только из-за своего положения на растении. Родовое название Lepidophyllum — это оригинальное название сохранившихся листьев Lepidodendrid, но, поскольку это название уже использовалось для отдельного цветкового растения, название Lepidophylloides . сегодня вместо него используется [ 14 ] Вдоль всей пластинки Lepidophylloides . единый проводящий пучок окаймлен неглубокими бороздками на абаксиальной поверхности Устьица погружены в ямки, выстроенные рядами, параллельными этим бороздкам. Сосудистый пучок листа окружает гиподермальная зона волокон. [ 6 ]
Подземные органы
[ редактировать ]
Подземным органам Lepidodendrales присвоено родовое название Stigmaria . Эти структуры являются одними из наиболее распространенных окаменелостей ликопсидов и основным органом, обнаруженным в слое глины под большинством угольных отложений каменноугольного периода; этот слой глины представляет собой слой почвы, в котором укоренились растения. Несмотря на существование множества видов стигмарий , наше понимание подземных органов основано прежде всего на широко распространенном виде Stigmaria ficoides . [ 15 ] Стигмарические органы берут начало от основания ствола в виде четырех основных осей, идущих горизонтально, что приводит к относительно неглубокой корневой системе. Боковые придатки прикреплены к каждой оси по спирали. Эти придатки отпадали по мере роста растения, в результате чего образовывались характерные круглые внешние шрамы, характерные для ископаемых образцов стигмарий . Хотя эти придатки часто называют «стигматическими корешками», их спиральное расположение и опадение роста на самом деле более характерны для листьев, чем для современных боковых корней. Четыре основные оси Стигмарии часто дихотомируются, образуя обширную подземную систему, радиус которой может достигать 15 м (49 футов). Корешки различаются по размеру: до 40 см (16 дюймов) в длину и 0,5–1 см (0,20–0,39 дюйма) в ширину, обычно сужаются дистально и не образуют дихотомии. В каждом корешке имеется небольшая монархическая сосудистая жилка, окруженная компактной внутренней корой. За пределами этой внутренней коры находится полая средняя кора и тонкая внешняя кора; иногда связь распространяется от внутренней коры к внешней коре. [ 16 ] Первичная ксилема стигмарий эндархична . и расположена в виде полос, окруженных сосудистым камбием Вторичные трахеиды ксилемы расположены радиальными линиями и содержат лестничные утолщения стенок с фибриллами, идентичными таковым в надземных ветвях. не было обнаружено вторичной флоэмы В ископаемых образцах стигмарий , а сосудистый камбий был односторонним, а первичная флоэма обеспечивала транслокацию. Радиально ориентированные трахеиды в большинстве осей стигмарий образованы утолщающейся меристемой, а не сосудистым камбием. [ 17 ]
Развитие подземных органов Lepidodendrales, вероятно, было сходно с развитием надземных стеблей. Однако некоторые особенности функционирования этих органов еще предстоит выявить, а некоторые современные особенности корней у стигмарий отсутствуют . Спиральное расположение придатков корешков отличается от неправильного расположения современных корней. Корневых волосков не обнаружено, хотя грибы в некоторых клетках кортикальной паренхимы могли функционировать как микоризы. Сосудистый пучок монарха в корешках двусторонне-симметричен, но современные корни имеют радиально-симметричную сосудистую ткань, хотя сосудистые пучки в листьях двусторонне симметричны. Кроме того, корешки регулярно отпадали от оси по мере того, как растение росло аналогично процессу опадения листьев. Однако у современных растений опадение корней неизвестно. Эти особенности корешков позволяют предположить, что они гомологичны воздушным листьям Lepidodendrales, но модифицированы для выполнения закрепляющих и поглощающих функций. Это означает, что подземные органы растений возникли как эволюционная модификация надземных органов. [ 18 ]

Несмотря на то, что некоторые растения Lepidodendrales достигают 40-метровой высоты (130 футов), их рыльцевая система обычно была неглубокой, и поэтому сомнительно, как подземные органы могли поддерживать огромные деревья, тем более что многие растения росли в перенасыщенной, водянистой почве, которая в основном была нестабильный. Возникли разные предположения, объясняющие их рост и корневую систему: возможно, обширный горизонтальный рост корневых осей обеспечивал достаточную поддержку или что кроны соседних деревьев могли запутываться и обеспечивать взаимную поддержку. Характер древесины и плотность кроны современных деревьев могут иметь большое влияние на выкорчевывание деревьев, и, поскольку у древесных ликопсидов было мало вторичной ксилемы и густые кроны, они, возможно, лучше подходили для стояния в вертикальном положении. [ 19 ]
Репродуктивные органы
[ редактировать ]
Репродуктивные органы лепидодендридов состояли из шишек или шишек на дистальных ветвях кроны. У Synchysidendron шишки располагаются на поздно сформировавшихся ветвях кроны, а у Diaphorodendron - на лиственных боковых ветвях. [ 20 ] Шишки могут вырасти до значительно больших размеров, так как экземпляры Lepidostrobus goldernbergii имеют длину более 50 см (20 дюймов). Шишки состоят из центральной оси со спорофиллами , расположенными спирально; спорангии расположены на адаксиальной поверхности спорофиллов и перевернуты дистально, перекрывая расположенные выше спорофиллы. Часть спорофилла обычно простирается вниз, образуя пятку или другое дистальное расширение. Язычок можно найти в небольшой ямке дистальнее спорангия. Хотя Lepidostrobus - наиболее распространенное название шишек Lepidodendrales, это название использовалось для образцов любой формы сохранности, а также для моноспорангиатных и биспорангиатных форм, поэтому часто возникают таксономические проблемы. [ 6 ] Были предприняты попытки разубедить эту таксономическую путаницу. Некоторые предположили, что название Lepidostrobus должно обозначать только моноспорангиатные шишки, а название Flemingites описывает биспорангиатные шишки, в то время как другие использовали морфологию шишек, чтобы попытаться лучше дифференцировать виды внутри Lepidostrobus . [ 21 ] [ 22 ]
Образцы эмбрионов были обнаружены в шишке Bothrodendrostrobus . [ 23 ] Эмбрион начинается как неваскуляризированная глобулярная структура, обнаруженная в ткани мегагаметофита , а у более зрелых экземпляров через трехдольный шов проходят два васкуляризированных придатка, представляющие первый побег и первый корень. Генерация гаметофитов Lepidodendrales плохо изучена и основана на небольшом количестве экземпляров, но шишки Flemingites schopfii демонстрируют хорошо сохранившиеся признаки микро- и мегагаметофитной фазы. [ 24 ] По сравнению с гаметофитами современных ликопсид F. schopfii имеет микрогаметофиты, наиболее сходные с современными Selaginella , тогда как мегагаметофиты более сходны с Isoetes . Другие хорошо сохранившиеся гаметофиты лепидодендрид были обнаружены в спорах Lepidodendron rhodumnense, окаменевших в кремне позднего Визе . [ 25 ]
Рост
[ редактировать ]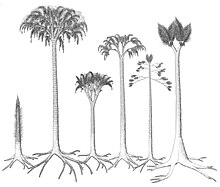
На ранних стадиях роста древесные ликофиты росли в виде неразветвленных стволов, листья которых росли непосредственно из подушек/оснований листьев. На более позднем этапе цикла роста, в зависимости от вида, ствол образовывал либо серию латерально (перпендикулярно стволу) растущих ветвей с дихотомическим характером роста, либо крону из дихотомических ветвей. Предполагается, что некоторые виды Sigillaria вообще не разветвляются. На более поздних стадиях роста пластинки листьев в нижних частях ствола опадали, хотя скорость опадания не была быстрой, поскольку были обнаружены большие стебли с оставшимися прикрепленными листьями. Основания листьев оставались на стволе до тех пор, пока самые крупные стебли не отслоились, обнажив перидерму. Скорость роста древесных ликофитов оспаривается; некоторые авторы утверждают, что они имели быстрый жизненный цикл: вырастали до максимальных размеров, размножались, а затем умирали всего за 10–15 лет, в то время как другие авторы утверждают, что эти темпы роста завышены. [ 26 ] Было высказано предположение, что древесные ликофиты имели такой же механизм фиксации углерода, как и современные иголки , где углерод поглощался из окружающих отложений, а повышенные концентрации углекислого газа во внутренних газовых пространствах позволяли увеличить поглощение углерода. [ 27 ] Большинство частей растения, включая листья, стебли и части укореняющихся структур ризофора, вероятно, были фотосинтезирующими. [ 26 ]
Экология
[ редактировать ]Предполагается, что древесные ликофиты отдавали предпочтение нарушенным местообитаниям. [ 26 ] Большие количества биомассы , которые были ответственны за формирование широко распространенных во всем мире угольных пластов каменноугольного периода, производились преимущественно древесными ликофитами. Предполагается, что Lepidodendrales ответственны за почти 70% растительного материала в Вестфальских угольно-болотных лесах Америки. [ 28 ] хотя в конце Вестфальского периода представители Lepidodendrales пришли в упадок и стали составлять только 5% угольной биомассы. [ 29 ] К концу каменноугольного периода древовидные ликопсиды в значительной степени вымерли в Северной Америке и Европе, поскольку древовидные папоротники начали приобретать все большую популярность, хотя древесные ликопсиды сохранялись в Китае до средней перми . [ 6 ] Некоторые ученые предположили, что сокращение численности лепидодендридов в этот период было результатом тектонической активности Варискана , создавшей нестабильные условия за счет уменьшения размеров экосистем угольных болот. [ 29 ] в то время как другие предполагают, что их снижение произошло из-за изменения климата; некоторые ученые предполагают комбинацию этих теорий о том, что тектоническая активность вызвала изменения в цветочном составе, которые спровоцировали изменение климата, что, в свою очередь, привело к этому упадку. [ 30 ]
Таксономия
[ редактировать ]Среди Lycopodiopsida Lepidodendrales считаются более близкими родственниками Isoetales (включая современные игольчатые ), чем плаунами или колючими мхами . [ 31 ] Некоторые авторы не используют Lepidodendrales, а вместо этого включают древесные ликофиты в состав Isoetales. [ 32 ]
Разновидность
[ редактировать ]Различные экземпляры Lepidodendrales исторически были отнесены к семейству Lepidodendron , роду, определяемому морфологией листовых подушек. ДиМишель основал Diaphorodendron, чтобы избежать двусмысленности в отношении этих широко распространенных экземпляров, включая некоторые структурно сохранившиеся экземпляры, которые ранее были членами Lepidodendron . Позже диафородендрон был разделен на два рода Diaphorodendron и Synchysidendron , и эти роды были помещены в новое семейство Diaphorodendraceae. [ 20 ] Синапоморфиями этого нового семейства являются сердцевидная протостела и уплощенный в дорсивентральном направлении мегаспорангий. Синапоморфии семейства Lepidodendraceae представляют собой уплощенный с двух сторон мегаспорангий и подлистной парихнос, простирающийся ниже рубца листа. [ 33 ] Родовые названия Лепидодендрон и Диафородендрон сегодня описывают как сохранившиеся в клетках сегменты стебля, так и целые растения, включая их лиственные органы, подземные органы и репродуктивные органы. В частности, родовое название Лепидодендрон обычно используется для описания компрессионных образцов , которые имеют определенный тип морфологии листовой подушки. [ 6 ]
Кроме того, к Lepidodendrales было идентифицировано множество «таксонов органов»: корни ( Stigmaria ), листья и шишки ( Lepidostrobus ) первоначально получили другое название рода и вида, прежде чем можно было доказать, что они принадлежали одному и тому же организму.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ A. V. Lopatin (2012). Палеонтологический музей имени Ю.А. Орлова (The Orlov Museum of Paleontology) . Moscow: PIN RAN. p. 56. ISBN 978-5-903825-14-1 . Проверено 5 октября 2020 г.
- ^ V. V. Alekhin (1961). Geografiia rastenii s osnovani botaniki (Geography of plants and basics of botany) . Gos. nauchno-pedagog. izd-vo. p. 167 . Retrieved 2020-10-05 .
- ^ Викелл, Дэвид; Куо, Ли-Янг; Ян, Сяо-Пей; Дхабалия Ашок, Амра; Ирисарри, Икер; Дадрас, Армин; Де Врис, Софи; Де Врис, Ян; Хуан, Яо-Моан; Ли, Чжэн; Баркер, Майкл С.; Хартвик, Нолан Т.; Майкл, Тодд П.; Ли, Фэй-Вэй (2021). «Подводный фотосинтез CAM, объясненный геномом Isoetes» . Природные коммуникации . 12 (1): 6348. Бибкод : 2021NatCo..12.6348W . дои : 10.1038/s41467-021-26644-7 . ПМЦ 8566536 . ПМИД 34732722 .
- ^ «Селагинеллалес» . НаукаДирект . 2009 . Проверено 17 января 2022 г.
- ^ Смелый, Гарольд К.; Си Джей Алексопулос; Т. Делеворяс (1987). Морфология растений и грибов (5-е изд.). Нью-Йорк: Харпер-Коллинз. стр. 496–503. ISBN 0-06-040839-1 .
- ^ Jump up to: а б с д и ж Тейлор, Эдит Л.; Тейлор, Томас Н.; Крингс, Майкл (2009). Палеоботаника: биология и эволюция ископаемых растений . Академическая пресса.
- ^ Томас, бакалавр (1966). «Кутикула стебля лепидодендроида» . Новый фитолог . 65 (3). Интернет-библиотека Wiley: 296–303. дои : 10.1111/j.1469-8137.1966.tb06365.x .
- ^ Томас, бакалавр (1974). «Лепидодендроидная стома».
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Уайлдер, Джордж Дж. (1970). «Строение трахеид трех видов Lycopodium». Американский журнал ботаники . 57 (9). Интернет-библиотека Уайли: 1093–1107. дои : 10.1002/j.1537-2197.1970.tb09913.x .
- ^ Эггерт, Д.А.; Канемото, Нью-Йорк (1977). «Стеблевая флоэма лепидодендрона среднего Пенсильвании». Ботанический вестник . 138 (1). Издательство Чикагского университета: 102–111. дои : 10.1086/336903 . S2CID 85391641 .
- ^ Хирмер, Макс; Пиа, Юлиус; Тролль, Вильгельм (1927). «Справочник по палоботанике: Thallophyta-Bryophyta-Pteridophyta». 1 . Р. Ольденбург.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Эггерт, Дональд Алан (1961). «Онтогенез каменноугольных древесных Lycopsida» . Палеонтографический отдел Б. Schweizerbart'sche Verlagsbuchhandlung: 43–92.
- ^ Чалонер, В.Г.; Мейер-Берто, Б. (1983). «Рост листьев и стеблей Lepidodendrales» . Ботанический журнал Линнеевского общества . 86 (1–2). Издательство Оксфордского университета: 135–148. дои : 10.1111/j.1095-8339.1983.tb00721.x .
- ^ Рыхновский, Войтех (2013). «Ревизия видов Lepidodendron lycopodioides и L. selaginoides из каменноугольного периода Богемии» . Карлов университет, факультет естественных наук.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Уильямсон, Уильям Кроуфорд (1887). Монография по морфологии и гистологии Stigmaria ficoides . Том. 40. Палеонтографическое общество.
- ^ Вайс, Фредерик Эрнест (1902). «Сосудистые ветви корешков стигмарий». Анналы ботаники . 16 (63). JSTOR: 559–573. JSTOR 43235190 .
- ^ Ротвелл, Гар В.; Прайор, Джанель С. (1991). «Динамика развития древесных ликофитов - апикальный и боковой рост Stigmaria ficoides». Американский журнал ботаники . 78 (12). Интернет-библиотека Айли: 1740–1745. дои : 10.1002/j.1537-2197.1991.tb14538.x .
- ^ Франкенберг, Джулиан М.; Эггерт, Дональд А. (1969). «Окаменелые стигмарии из Северной Америки: Часть I. Stigmaria ficoides, подземные части Lepidodendraaceae». Палеонтографический отдел Б. Schweizerbart'sche Verlagsbuchhandlung: 1–47.
- ^ Никлас, Карл Дж. (1992). Биомеханика растений: инженерный подход к форме и функциям растений . Пресса Чикагского университета. ISBN 9780226586311 .
- ^ Jump up to: а б ДиМишель, Уильям А.; Бейтман, Ричард М. (1992). «Diaphorodendraceae, семейство nov. (Lycopsida: Carboniferous): систематика и эволюционные взаимоотношения Diaphorodendraceae и Synchysidendron, gen. nov». Американский журнал ботаники . 79 (6). Интернет-библиотека Wiley: 605–617. дои : 10.1002/j.1537-2197.1992.tb14602.x .
- ^ Брэк-Хейнс, Шейла Д.; Томас, Барри А. (1983). «Повторное исследование Лепидостробуса Броньярта» . Ботанический журнал Линнеевского общества . 86 (1–2). Издательство Оксфордского университета: 125–133. дои : 10.1111/j.1095-8339.1983.tb00720.x .
- ^ Бек, Иржи; Оплюштил, Станислав (2006). «Шесть редких видов Lepidostrobus из Пенсильвании Чешской Республики и их влияние на классификацию ликоспор» . Обзор палеоботаники и палинологии . 139 (1–4). Эльзевир: 211–226. Бибкод : 2006RPaPa.139..211B . дои : 10.1016/j.revpalbo.2006.01.003 .
- ^ Стабблфилд, Сара П.; Ротвелл, Гар В. (1981). «Эмбриогенез и репродуктивная биология Bothrodendrostrobus mundus (Lycopsida)». Американский журнал ботаники . 68 (5). Интернет-библиотека Wiley: 625–634. дои : 10.1002/j.1537-2197.1981.tb12394.x .
- ^ Брэк-Ханес, Шейла Д. (1978). «О мегагаметофитах двух лепидодендрацеевых шишек». Ботанический вестник . 139 (1). Издательство Чикагского университета: 140–146. дои : 10.1086/336979 . S2CID 84420384 .
- ^ Галтье, Дж. (1964). «ПАЛЕБОТАНИКА-НА ЖЕНСКИХ ГАМЕТОФИТАХ ЛЕПИДОДЕНДРАКОВ». Еженедельные отчеты сессий Академии наук . 258 (9). GOTHIE-VILLARS/EDITIONS ELSEVIER 23 RUE LINOIS, 75015 ПАРИЖ, ФРАНЦИЯ: 2625.
- ^ Jump up to: а б с Томас, Барри А.; Клил, Кристофер Дж. (май 2018 г.). «Рост древесных ликофитов в угольных болотах позднего карбона» . Новый фитолог . 218 (3): 885–890. дои : 10.1111/nph.14903 . ПМИД 29282734 .
- ^ Грин, Вашингтон (07 августа 2010 г.). «Функция аэренхимы у древовидных ликопсидов: свидетельства незнакомой метаболической стратегии» . Труды Королевского общества B: Биологические науки . 277 (1692): 2257–2267. дои : 10.1098/rspb.2010.0224 . ISSN 0962-8452 . ПМЦ 2894907 . ПМИД 20356894 .
- ^ ДиМишель, Уильям А. (1985). «Диафородендрон, генерация ноября, отдельный от лепидодендрона (пенсильванский возраст)». Систематическая ботаника . 10 (4). JSTOR: 453–458. дои : 10.2307/2419138 . JSTOR 2419138 .
- ^ Jump up to: а б Керп, Ганс (2000). «Модернизация ландшафтов в период позднего палеозоя-раннего мезозоя» . Документы Палеонтологического общества . 6 . Издательство Кембриджского университета: 79–114. дои : 10.1017/S1089332600000723 .
- ^ ДиМишель, Уильям А.; Филлипс, Том Л. (1996). «Изменение климата, исчезновение растений и восстановление растительности во время переходного периода среднего и позднего Пенсильванского периода: пример тропической среды, образующей торф в Северной Америке». Геологическое общество, Лондон, специальные публикации . 102 (1). Лондонское геологическое общество: 201–221. дои : 10.1111/j.1472-4669.2005.00043.x . S2CID 129219852 .
- ^ Спенсер, Виктория; Немец Венза, Зоя; Харрисон, Сесили Джилл (май 2021 г.). «Чему ликофиты могут научить нас об эволюции и развитии растений? Современные взгляды на древнюю линию» . Эволюция и развитие . 23 (3): 174–196. дои : 10.1111/ede.12350 . ISSN 1520-541X . ПМИД 32906211 . S2CID 221622444 .
- ^ Бейтман, Ричард М.; ДиМишель, Уильям А. (октябрь 2021 г.). «Побег из добровольных ограничений таксономии «колеи шин»» . Таксон . 70 (5): 1062–1077. дои : 10.1002/tax.12540 . ISSN 0040-0262 . S2CID 238818487 .
- ^ ДиМишель, Уильям А.; Бейтман, Ричард М. (1996). «Ризоморфные ликопсиды: пример палеоботанической классификации». Систематическая ботаника . 21 (4). JSTOR: 535–552. дои : 10.2307/2419613 . JSTOR 2419613 .
Дальнейшее чтение
[ редактировать ]- Дэвис, Пол и Кенрик, Пол. Ископаемые растения . Смитсоновские книги, Вашингтон, округ Колумбия (2004).
- Морран, Робин, К.; Естественная история папоротников . Тимбер Пресс (2004). ISBN 0-88192-667-1