Лист



- Апекс
- Средняя вена (первичная вена)
- Вторичная вена.
- Ламина.
- Край листа
- Черешок
- Бутон
- Корень

Внизу: скунсовая капуста, Symplocarpus foetidus (простой лист).
- Апекс
- Первичная вена
- Вторичная вена
- пластинка
- Край листа
- Рачис
Лист Leaves ( мн.ч .: ) — главный придаток стебля сосудистого растения . [1] обычно располагаются сбоку над землей и специализируются на фотосинтезе . Листья вместе называются листвой , например «осенняя листва». [2] [3] в то время как листья, стебель, цветы и плоды вместе образуют систему побегов . [4] В большинстве листьев первичной фотосинтетической тканью является палисадный мезофилл , расположенный на верхней стороне пластинки или пластинки листа. [1] но у некоторых видов, включая зрелую листву эвкалипта , [5] палисадный мезофилл присутствует с обеих сторон, и листья называются изобилатеральными. Большинство листьев уплощены и имеют отчетливые верхнюю ( адаксиальную ) и нижнюю ( абаксиальную ) поверхности, различающиеся по цвету, опушению, количеству устьиц (пор, поглощающих и выводящих газы), количеству и строению эпикутикулярного воска и другим признакам. Листья в основном имеют зеленый цвет из-за присутствия соединения под названием хлорофилл , которое необходимо для фотосинтеза, поскольку оно поглощает световую энергию солнца . Лист с более светлыми или белыми пятнами или краями называется пестрым листом .
Листья могут иметь самую разную форму, размер, текстуру и цвет. Широкие плоские листья со сложным жилкованием цветковых растений известны как мегафиллы , а виды, несущие их, большинство - как широколистные или мегафилловые растения, к которым также относятся акрогимноспермовые и папоротники . У плаунов разного эволюционного происхождения листья простые (с одной жилкой) и известны как микрофиллы . [6] Некоторые листья, например чешуи луковицы , не находятся над землей. У многих водных видов листья погружены в воду. Суккулентные растения часто имеют толстые сочные листья, но некоторые листья лишены основной фотосинтетической функции и могут погибнуть по мере созревания, как у некоторых катафиллов и колючек . Более того, некоторые виды листообразных структур, обнаруженных у сосудистых растений, не полностью им гомологичны. Примеры включают уплощенные стебли растений, называемые филлокладами и кладодами , а также уплощенные стебли листьев, называемые филлодиями , которые отличаются от листьев как по своей структуре, так и по происхождению. [3] [7] Некоторые структуры несосудистых растений выглядят и функционируют так же, как листья. включают филлиды мхов . и печеночников Примеры
Общие характеристики
[ редактировать ]Листья — важнейшие органы большинства сосудистых растений. [8] Зеленые растения автотрофны , то есть они не получают пищу от других живых существ, а вместо этого создают себе пищу посредством фотосинтеза . Они улавливают энергию солнечного света и используют ее для производства простых сахаров , таких как глюкоза и сахароза , из углекислого газа и воды. Затем сахара сохраняются в виде крахмала , который далее перерабатывается путем химического синтеза в более сложные органические молекулы, такие как белки или целлюлоза , основной структурный материал в стенках растительных клеток, или метаболизируется посредством клеточного дыхания , чтобы обеспечить химическую энергию для запуска клеточных процессов. Листья всасывают воду из земли в транспирационном потоке через сосудистую проводящую систему, известную как ксилема , и получают углекислый газ из атмосферы путем диффузии через отверстия, называемые устьицами, во внешнем покровном слое листа ( эпидермисе ), при этом листья ориентированы так, чтобы максимизировать их воздействие солнечного света. После синтеза сахара его необходимо транспортировать в места активного роста, например, в побеги растений. и корни . Сосудистые растения транспортируют сахарозу в специальной ткани, называемой флоэмой . Флоэма и ксилема параллельны друг другу, но транспорт веществ обычно происходит в противоположных направлениях. Внутри листа эти сосудистые системы разветвляются (разветвляются), образуя вены, которые снабжают как можно большую часть листа, гарантируя, что клетки, осуществляющие фотосинтез, находятся близко к транспортной системе. [9]
Обычно листья широкие, плоские и тонкие (сплющенные в дорзивентральном направлении), что увеличивает площадь поверхности, непосредственно подвергающейся воздействию света, и позволяет свету проникать в ткани и достигать хлоропластов , способствуя тем самым фотосинтезу. Они расположены на растении так, чтобы максимально эффективно освещать свои поверхности, не затеняя друг друга, но есть много исключений и сложностей. Например, растения, приспособленные к ветреным условиям, могут иметь свисающие листья, как, например, у многих ив и эвкалиптов . Плоская или ламинарная форма также максимизирует тепловой контакт с окружающим воздухом , способствуя охлаждению. Функционально, помимо осуществления фотосинтеза, лист является основным местом транспирации , обеспечивая энергию, необходимую для вытягивания потока транспирации вверх от корней и гуттации .
Многие хвойные деревья имеют тонкие игольчатые или чешуйчатые листья, что может быть полезно в холодном климате с частым снегом и морозами. [10] Они интерпретируются как производные от крупнолистных листьев их девонских предков. [6] Некоторые формы листьев приспособлены модулировать количество поглощаемого ими света, чтобы избежать или смягчить чрезмерное тепло, повреждение ультрафиолетом или высыхание, или пожертвовать эффективностью поглощения света в пользу защиты от травоядных. Для ксерофитов основным ограничением является не световой поток или интенсивность , а засуха. [11] Некоторые оконные растения, такие как виды Fenestraria и некоторые виды Haworthia , такие как Haworthia tesselata и Haworthia truncata, являются примерами ксерофитов. [12] и Bulbine mesembryanthemoides . [13]
Листья также служат для хранения химической энергии и воды (особенно у суккулентов ) и могут стать специализированными органами, выполняющими другие функции, например, усики гороха и других бобовых, защитные колючки кактусов и ловушки для насекомых у плотоядных растений, таких как непентес и саррацения . [14] Листья являются основными структурными единицами, из которых построены шишки у голосеменных растений (каждая чешуя шишки представляет собой модифицированный лист мегафилла, известный как спорофилл). [6] : 408 и из чего построены цветы цветковых растений . [6] : 445

Внутренняя организация большинства видов листьев эволюционировала таким образом, чтобы максимизировать воздействие на фотосинтетические органеллы , хлоропласты , света и увеличить поглощение углекислого газа , одновременно контролируя потерю воды. Их поверхности водонепроницаемы кутикулой растения , а газообмен между клетками мезофилла и атмосферой контролируется мельчайшими (длина и ширина измеряются десятками мкм) отверстиями, называемыми устьицами , которые открываются или закрываются, регулируя скорость обмена углекислого газа (CO 2 ) . ), кислорода (О 2 ) и водяного пара во внутреннее межклеточное пространство и из него. Открытие устьица контролируется тургорным давлением в паре замыкающих клеток , окружающих устьичное отверстие. В любом квадратном сантиметре листа растения может быть от 1000 до 100 000 устьиц. [15]


Форма и структура листьев значительно различаются от вида к виду растений, во многом в зависимости от их адаптации к климату и доступному свету, а также от других факторов, таких как выпас животных (например, олени), наличие питательных веществ и экологическая конкуренция со стороны других растений. . Значительные изменения в типе листьев происходят и внутри видов, например, по мере взросления растения; Например, виды эвкалипта обычно имеют изобилатеральные свисающие листья, когда созревают и доминируют над своими соседями; однако такие деревья, как правило, имеют прямостоячие или горизонтальные дорзивентральные листья в качестве сеянцев, когда их рост ограничен доступным светом. [16] Другие факторы включают необходимость сбалансировать потерю воды при высокой температуре и низкой влажности с необходимостью поглощать углекислый газ из атмосферы. У большинства растений листья также являются основным органом, ответственным за транспирацию и гуттацию (капельки жидкости, образующиеся по краям листьев).
Листья также могут хранить пищу и воду и соответствующим образом модифицируются для выполнения этих функций, например, в листьях суккулентных растений и в чешуях луковиц . Концентрация фотосинтетических структур в листьях требует, чтобы они были богаче белками , минералами и сахарами, чем, скажем, ткани древесного стебля. Соответственно, листья занимают важное место в рационе многих животных .

Соответственно, листья представляют собой тяжелые инвестиции со стороны растений, несущих их, а их сохранение или утилизация являются предметом тщательно продуманных стратегий борьбы с давлением вредителей, сезонными условиями и защитными мерами, такими как рост шипов и образование фитолитов . лигнины , дубильные вещества и яды .
Лиственные растения в регионах с холодным или умеренным климатом обычно сбрасывают листья осенью, тогда как в районах с суровым засушливым сезоном некоторые растения могут сбрасывать листья до тех пор, пока засушливый сезон не закончится. В любом случае можно ожидать, что сброшенные листья внесут сохраненные питательные вещества в почву, куда они падают.

Напротив, многие другие несезонные растения, такие как пальмы и хвойные деревья, сохраняют листья в течение длительного времени; Вельвичия сохраняет два основных листа на протяжении всей жизни, которая может превышать тысячу лет.
Листовидные органы мохообразных (например, мхов и печеночников ), известные как филлиды , сильно отличаются морфологически от листьев сосудистых растений . В большинстве случаев они лишены сосудистой ткани, имеют толщину всего в одну клетку и не имеют кутикулы , устьиц или внутренней системы межклеточных пространств. (Филлиды семейства мхов Polytrichaceae являются заметным исключением.) Филлиды мохообразных присутствуют только на гаметофитах , тогда как листья сосудистых растений присутствуют только на спорофитах . В дальнейшем они могут развиваться либо в вегетативные, либо в репродуктивные структуры. [14]
Simple, vascularized leaves (microphylls), such as those of the early Devonian lycopsid Baragwanathia, first evolved as enations, extensions of the stem. True leaves or euphylls of larger size and with more complex venation did not become widespread in other groups until the Devonian period, by which time the carbon dioxide concentration in the atmosphere had dropped significantly. This occurred independently in several separate lineages of vascular plants, in progymnosperms like Archaeopteris, in Sphenopsida, ferns and later in the gymnosperms and angiosperms. Euphylls are also referred to as macrophylls or megaphylls (large leaves).[6]
Morphology
[edit]

A structurally complete leaf of an angiosperm consists of a petiole (leaf stalk), a lamina (leaf blade), stipules (small structures located to either side of the base of the petiole) and a sheath. Not every species produces leaves with all of these structural components. The proximal stalk or petiole is called a stipe in ferns. The lamina is the expanded, flat component of the leaf which contains the chloroplasts. The sheath is a structure, typically at the base that fully or partially clasps the stem above the node, where the leaf is attached. Leaf sheathes typically occur in Poaceae (grasses) and Apiaceae (umbellifers). Between the sheath and the lamina, there may be a pseudopetiole, a petiole like structure. Pseudopetioles occur in some monocotyledons including bananas, palms and bamboos.[18] Stipules may be conspicuous (e.g. beans and roses), soon falling or otherwise not obvious as in Moraceae or absent altogether as in the Magnoliaceae. A petiole may be absent (apetiolate), or the blade may not be laminar (flattened). The petiole mechanically links the leaf to the plant and provides the route for transfer of water and sugars to and from the leaf. The lamina is typically the location of the majority of photosynthesis. The upper (adaxial) angle between a leaf and a stem is known as the axil of the leaf. It is often the location of a bud. Structures located there are called "axillary".
External leaf characteristics, such as shape, margin, hairs, the petiole, and the presence of stipules and glands, are frequently important for identifying plants to family, genus or species levels, and botanists have developed a rich terminology for describing leaf characteristics. Leaves almost always have determinate growth. They grow to a specific pattern and shape and then stop. Other plant parts like stems or roots have non-determinate growth, and will usually continue to grow as long as they have the resources to do so.
The type of leaf is usually characteristic of a species (monomorphic), although some species produce more than one type of leaf (dimorphic or polymorphic). The longest leaves are those of the Raffia palm, R. regalis which may be up to 25 m (82 ft) long and 3 m (9.8 ft) wide.[19] The terminology associated with the description of leaf morphology is presented, in illustrated form, at Wikibooks.

Where leaves are basal, and lie on the ground, they are referred to as prostrate.
Basic leaf types
[edit]
Perennial plants whose leaves are shed annually are said to have deciduous leaves, while leaves that remain through winter are evergreens. Leaves attached to stems by stalks (known as petioles) are called petiolate, and if attached directly to the stem with no petiole they are called sessile.[20]
- Ferns have fronds.
- Conifer leaves are typically needle- or awl-shaped or scale-like, they are usually evergreen, but can sometimes be deciduous. Usually, they have a single vein.
- Flowering plant (Angiosperm) leaves: the standard form includes stipules, a petiole, and a lamina.
- Lycophytes have microphylls.
- Sheath leaves are the type found in most grasses and many other monocots.
- Other specialized leaves include those of Nepenthes, a pitcher plant.
Dicot leaves have blades with pinnate venation (where major veins diverge from one large mid-vein and have smaller connecting networks between them). Less commonly, dicot leaf blades may have palmate venation (several large veins diverging from petiole to leaf edges). Finally, some exhibit parallel venation.[20]
Monocot leaves in temperate climates usually have narrow blades, and usually parallel venation converging at leaf tips or edges. Some also have pinnate venation.[20]
Arrangement on the stem
[edit]The arrangement of leaves on the stem is known as phyllotaxis.[21] A large variety of phyllotactic patterns occur in nature:


- Alternate
- One leaf, branch, or flower part attaches at each point or node on the stem, and leaves alternate direction, to a greater or lesser degree, along the stem.
- Basal
- Arising from the base of the plant.
- Cauline
- Attached to the aerial stem.
- Opposite
- Two leaves, branches, or flower parts attach at each point or node on the stem. Leaf attachments are paired at each node.
- Decussate
- An opposite arrangement in which each successive pair is rotated 90° from the previous.
- Whorled, or verticillate
- Three or more leaves, branches, or flower parts attach at each point or node on the stem. As with opposite leaves, successive whorls may or may not be decussate, rotated by half the angle between the leaves in the whorl (i.e., successive whorls of three rotated 60°, whorls of four rotated 45°, etc.). Opposite leaves may appear whorled near the tip of the stem. Pseudoverticillate describes an arrangement only appearing whorled, but not actually so.
- Rosulate
- Leaves form a rosette.
- Rows
- The term, distichous, literally means two rows. Leaves in this arrangement may be alternate or opposite in their attachment. The term, 2-ranked, is equivalent. The terms, tristichous and tetrastichous, are sometimes encountered. For example, the "leaves" (actually microphylls) of most species of Selaginella are tetrastichous, but not decussate.
In the simplest mathematical models of phyllotaxis, the apex of the stem is represented as a circle. Each new node is formed at the apex, and it is rotated by a constant angle from the previous node. This angle is called the divergence angle. The number of leaves that grow from a node depends on the plant species. When a single leaf grows from each node, and when the stem is held straight, the leaves form a helix.
The divergence angle is often represented as a fraction of a full rotation around the stem. A rotation fraction of 1/2 (a divergence angle of 180°) produces an alternate arrangement, such as in Gasteria or the fan-aloe Kumara plicatilis. Rotation fractions of 1/3 (divergence angles of 120°) occur in beech and hazel. Oak and apricot rotate by 2/5, sunflowers, poplar, and pear by 3/8, and in willow and almond the fraction is 5/13.[22] These arrangements are periodic. The denominator of the rotation fraction indicates the number of leaves in one period, while the numerator indicates the number of complete turns or gyres made in one period. For example:
- 180° (or 1⁄2): two leaves in one circle (alternate leaves)
- 120° (or 1⁄3): three leaves in one circle
- 144° (or 2⁄5): five leaves in two gyres
- 135° (or 3⁄8): eight leaves in three gyres.
Most divergence angles are related to the sequence of Fibonacci numbers Fn. This sequence begins 1, 1, 2, 3, 5, 8, 13; each term is the sum of the previous two. Rotation fractions are often quotients Fn / Fn + 2 of a Fibonacci number by the number two terms later in the sequence. This is the case for the fractions 1/2, 1/3, 2/5, 3/8, and 5/13. The ratio between successive Fibonacci numbers tends to the golden ratio φ = (1 + √5)/2. When a circle is divided into two arcs whose lengths are in the ratio 1:φ, the angle formed by the smaller arc is the golden angle, which is 1/φ2 × 360° ≈ 137.5°. Because of this, many divergence angles are approximately 137.5°.
In plants where a pair of opposite leaves grows from each node, the leaves form a double helix. If the nodes do not rotate (a rotation fraction of zero and a divergence angle of 0°), the two helices become a pair of parallel lines, creating a distichous arrangement as in maple or olive trees. More common in a decussate pattern, in which each node rotates by 1/4 (90°) as in the herb basil. The leaves of tricussate plants such as Nerium oleander form a triple helix.
The leaves of some plants do not form helices. In some plants, the divergence angle changes as the plant grows.[23] In orixate phyllotaxis, named after Orixa japonica, the divergence angle is not constant. Instead, it is periodic and follows the sequence 180°, 90°, 180°, 270°.[24]
Divisions of the blade
[edit]
Two basic forms of leaves can be described considering the way the blade (lamina) is divided. A simple leaf has an undivided blade. However, the leaf may be dissected to form lobes, but the gaps between lobes do not reach to the main vein. A compound leaf has a fully subdivided blade, each leaflet of the blade being separated along a main or secondary vein. The leaflets may have petiolules and stipels, the equivalents of the petioles and stipules of leaves. Because each leaflet can appear to be a simple leaf, it is important to recognize where the petiole occurs to identify a compound leaf. Compound leaves are a characteristic of some families of higher plants, such as the Fabaceae. The middle vein of a compound leaf or a frond, when it is present, is called a rachis.
- Palmately compound
- The leaflets all have a common point of attachment at the end of the petiole, radiating like fingers of a hand; for example, Cannabis (hemp) and Aesculus (buckeyes).
- Pinnately compound
- Leaflets are arranged either side of the main axis, or rachis.
- Odd pinnate
- With a terminal leaflet; for example, Fraxinus (ash).
- Even pinnate
- Lacking a terminal leaflet; for example, Swietenia (mahogany). A specific type of even pinnate is bifoliolate, where leaves only consist of two leaflets; for example, Hymenaea.
- Bipinnately compound
- Leaves are twice divided: the leaflets (technically "subleaflets") are arranged along a secondary axis that is one of several branching off the rachis. Each leaflet is called a pinnule. The group of pinnules on each secondary vein forms a pinna; for example, Albizia (silk tree).
- Trifoliate (or trifoliolate)
- A pinnate leaf with just three leaflets; for example, Trifolium (clover), Laburnum (laburnum), and some species of Toxicodendron (for instance, poison ivy).
- Pinnatifid
- Pinnately dissected to the central vein, but with the leaflets not entirely separate; for example, Polypodium, some Sorbus (whitebeams). In pinnately veined leaves the central vein is known as the midrib.
Characteristics of the petiole
[edit]
Leaves which have a petiole (leaf stalk) are said to be petiolate.
Sessile (epetiolate) leaves have no petiole and the blade attaches directly to the stem. Subpetiolate leaves are nearly petiolate or have an extremely short petiole and may appear to be sessile.
In clasping or decurrent leaves, the blade partially surrounds the stem.
When the leaf base completely surrounds the stem, the leaves are said to be perfoliate, such as in Eupatorium perfoliatum.
In peltate leaves, the petiole attaches to the blade inside the blade margin.
In some Acacia species, such as the koa tree (Acacia koa), the petioles are expanded or broadened and function like leaf blades; these are called phyllodes. There may or may not be normal pinnate leaves at the tip of the phyllode.
A stipule, present on the leaves of many dicotyledons, is an appendage on each side at the base of the petiole, resembling a small leaf. Stipules may be lasting and not be shed (a stipulate leaf, such as in roses and beans), or be shed as the leaf expands, leaving a stipule scar on the twig (an exstipulate leaf).The situation, arrangement, and structure of the stipules is called the "stipulation".
- Free, lateral
- As in Hibiscus.
- Adnate
- Fused to the petiole base, as in Rosa.
- Ochreate
- Provided with ochrea, or sheath-formed stipules, as in Polygonaceae; e.g., rhubarb.
- Encircling the petiole base
- Interpetiolar
- Between the petioles of two opposite leaves, as in Rubiaceae.
- Intrapetiolar
- Between the petiole and the subtending stem, as in Malpighiaceae.
Veins
[edit]


Veins (sometimes referred to as nerves) constitute one of the most visible features of leaves. The veins in a leaf represent the vascular structure of the organ, extending into the leaf via the petiole and providing transportation of water and nutrients between leaf and stem, and play a crucial role in the maintenance of leaf water status and photosynthetic capacity. They also play a role in the mechanical support of the leaf.[25][26] Within the lamina of the leaf, while some vascular plants possess only a single vein, in most this vasculature generally divides (ramifies) according to a variety of patterns (venation) and form cylindrical bundles, usually lying in the median plane of the mesophyll, between the two layers of epidermis.[27] This pattern is often specific to taxa, and of which angiosperms possess two main types, parallel and reticulate (net like). In general, parallel venation is typical of monocots, while reticulate is more typical of eudicots and magnoliids ("dicots"), though there are many exceptions.[28][27][29]
The vein or veins entering the leaf from the petiole are called primary or first-order veins. The veins branching from these are secondary or second-order veins. These primary and secondary veins are considered major veins or lower order veins, though some authors include third order.[30] Each subsequent branching is sequentially numbered, and these are the higher order veins, each branching being associated with a narrower vein diameter.[31]
In parallel veined leaves, the primary veins run parallel and equidistant to each other for most of the length of the leaf and then converge or fuse (anastomose) towards the apex. Usually, many smaller minor veins interconnect these primary veins, but may terminate with very fine vein endings in the mesophyll. Minor veins are more typical of angiosperms, which may have as many as four higher orders.[30]
In contrast, leaves with reticulate venation have a single (sometimes more) primary vein in the centre of the leaf, referred to as the midrib or costa, which is continuous with the vasculature of the petiole. The secondary veins, also known as second order veins or lateral veins, branch off from the midrib and extend toward the leaf margins. These often terminate in a hydathode, a secretory organ, at the margin. In turn, smaller veins branch from the secondary veins, known as tertiary or third order (or higher order) veins, forming a dense reticulate pattern. The areas or islands of mesophyll lying between the higher order veins, are called areoles. Some of the smallest veins (veinlets) may have their endings in the areoles, a process known as areolation.[31] These minor veins act as the sites of exchange between the mesophyll and the plant's vascular system.[26] Thus, minor veins collect the products of photosynthesis (photosynthate) from the cells where it takes place, while major veins are responsible for its transport outside of the leaf. At the same time water is being transported in the opposite direction.[32][28][27]
The number of vein endings is very variable, as is whether second order veins end at the margin, or link back to other veins.[29] There are many elaborate variations on the patterns that the leaf veins form, and these have functional implications. Of these, angiosperms have the greatest diversity.[30] Within these the major veins function as the support and distribution network for leaves and are correlated with leaf shape. For instance, the parallel venation found in most monocots correlates with their elongated leaf shape and wide leaf base, while reticulate venation is seen in simple entire leaves, while digitate leaves typically have venation in which three or more primary veins diverge radially from a single point.[33][26][31][34]
In evolutionary terms, early emerging taxa tend to have dichotomous branching with reticulate systems emerging later. Veins appeared in the Permian period (299–252 mya), prior to the appearance of angiosperms in the Triassic (252–201 mya), during which vein hierarchy appeared enabling higher function, larger leaf size and adaption to a wider variety of climatic conditions.[30] Although it is the more complex pattern, branching veins appear to be plesiomorphic and in some form were present in ancient seed plants as long as 250 million years ago. A pseudo-reticulate venation that is actually a highly modified penniparallel one is an autapomorphy of some Melanthiaceae, which are monocots; e.g., Paris quadrifolia (True-lover's Knot). In leaves with reticulate venation, veins form a scaffolding matrix imparting mechanical rigidity to leaves.[35]
Morphology changes within a single plant
[edit]- Homoblasty
- Characteristic in which a plant has small changes in leaf size, shape, and growth habit between juvenile and adult stages, in contrast to;
- Heteroblasty
- Characteristic in which a plant has marked changes in leaf size, shape, and growth habit between juvenile and adult stages.
Anatomy
[edit]Medium-scale features
[edit]Leaves are normally extensively vascularized and typically have networks of vascular bundles containing xylem, which supplies water for photosynthesis, and phloem, which transports the sugars produced by photosynthesis. Many leaves are covered in trichomes (small hairs) which have diverse structures and functions.

Small-scale features
[edit]The major tissue systems present are
- The epidermis, which covers the upper and lower surfaces
- The mesophyll tissue, which consists of photosynthetic cells rich in chloroplasts. (also called chlorenchyma)
- The arrangement of veins (the vascular tissue)
These three tissue systems typically form a regular organization at the cellular scale. Specialized cells that differ markedly from surrounding cells, and which often synthesize specialized products such as crystals, are termed idioblasts.[36]
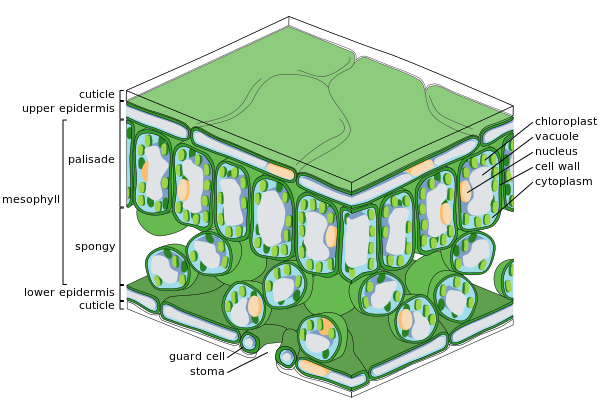
Major leaf tissues
[edit] Cross-section of a leaf
Cross-section of a leaf Epidermal cells
Epidermal cells Spongy mesophyll cells
Spongy mesophyll cells



Epidermis
[edit]
The epidermis is the outer layer of cells covering the leaf. It is covered with a waxy cuticle which is impermeable to liquid water and water vapor and forms the boundary separating the plant's inner cells from the external world. The cuticle is in some cases thinner on the lower epidermis than on the upper epidermis, and is generally thicker on leaves from dry climates as compared with those from wet climates.[37] The epidermis serves several functions: protection against water loss by way of transpiration, regulation of gas exchange and secretion of metabolic compounds. Most leaves show dorsoventral anatomy: The upper (adaxial) and lower (abaxial) surfaces have somewhat different construction and may serve different functions.
The epidermis tissue includes several differentiated cell types; epidermal cells, epidermal hair cells (trichomes), cells in the stomatal complex; guard cells and subsidiary cells. The epidermal cells are the most numerous, largest, and least specialized and form the majority of the epidermis. They are typically more elongated in the leaves of monocots than in those of dicots.
Chloroplasts are generally absent in epidermal cells, the exception being the guard cells of the stomata. The stomatal pores perforate the epidermis and are surrounded on each side by chloroplast-containing guard cells, and two to four subsidiary cells that lack chloroplasts, forming a specialized cell group known as the stomatal complex. The opening and closing of the stomatal aperture is controlled by the stomatal complex and regulates the exchange of gases and water vapor between the outside air and the interior of the leaf. Stomata therefore play the important role in allowing photosynthesis without letting the leaf dry out. In a typical leaf, the stomata are more numerous over the abaxial (lower) epidermis than the adaxial (upper) epidermis and are more numerous in plants from cooler climates.
Mesophyll
[edit]Most of the interior of the leaf between the upper and lower layers of epidermis is a parenchyma (ground tissue) or chlorenchyma tissue called the mesophyll (Greek for "middle leaf"). This assimilation tissue is the primary location of photosynthesis in the plant. The products of photosynthesis are called "assimilates".
In ferns and most flowering plants, the mesophyll is divided into two layers:
- An upper palisade layer of vertically elongated cells, one to two cells thick, directly beneath the adaxial epidermis, with intercellular air spaces between them. Its cells contain many more chloroplasts than the spongy layer. Cylindrical cells, with the chloroplasts close to the walls of the cell, can take optimal advantage of light. The slight separation of the cells provides maximum absorption of carbon dioxide. Sun leaves have a multi-layered palisade layer, while shade leaves or older leaves closer to the soil are single-layered.
- Beneath the palisade layer is the spongy layer. The cells of the spongy layer are more branched and not so tightly packed, so that there are large intercellular air spaces between them. The pores or stomata of the epidermis open into substomatal chambers, which are connected to the intercellular air spaces between the spongy and palisade mesophyll cell, so that oxygen, carbon dioxide and water vapor can diffuse into and out of the leaf and access the mesophyll cells during respiration, photosynthesis and transpiration.
Leaves are normally green, due to chlorophyll in chloroplasts in the mesophyll cells. Some plants have leaves of different colours due to the presence of accessory pigments such as carotenoids in their mesophyll cells.
Vascular tissue
[edit]
The veins are the vascular tissue of the leaf and are located in the spongy layer of the mesophyll. The pattern of the veins is called venation. In angiosperms the venation is typically parallel in monocotyledons and forms an interconnecting network in broad-leaved plants. They were once thought to be typical examples of pattern formation through ramification, but they may instead exemplify a pattern formed in a stress tensor field.[38][39][40]
A vein is made up of a vascular bundle. At the core of each bundle are clusters of twodistinct types of conducting cells:
- Xylem
- Cells that bring water and minerals from the roots into the leaf.
- Phloem
- Cells that usually move sap, with dissolved sucrose (glucose to sucrose) produced by photosynthesis in the leaf, out of the leaf.
The xylem typically lies on the adaxial side of the vascular bundle and the phloem typically lies on the abaxial side. Both are embedded in a dense parenchyma tissue, called the sheath, which usually includes some structural collenchyma tissue.
Leaf development
[edit]According to Agnes Arber's partial-shoot theory of the leaf, leaves are partial shoots,[41] being derived from leaf primordia of the shoot apex. Early in development they are dorsiventrally flattened with both dorsal and ventral surfaces.[14] Compound leaves are closer to shoots than simple leaves. Developmental studies have shown that compound leaves, like shoots, may branch in three dimensions.[42][43] On the basis of molecular genetics, Eckardt and Baum (2010) concluded that "it is now generally accepted that compound leaves express both leaf and shoot properties."[44] Many dicotyledonous leaves show endogenously driven daily rhythmicity in growth.[45][46][47]
Ecology
[edit]Biomechanics
[edit]Plants respond and adapt to environmental factors, such as light and mechanical stress from wind. Leaves need to support their own mass and align themselves in such a way as to optimize their exposure to the sun, generally more or less horizontally. However, horizontal alignment maximizes exposure to bending forces and failure from stresses such as wind, snow, hail, falling debris, animals, and abrasion from surrounding foliage and plant structures. Overall leaves are relatively flimsy with regard to other plant structures such as stems, branches and roots.[48]
Both leaf blade and petiole structure influence the leaf's response to forces such as wind, allowing a degree of repositioning to minimize drag and damage, as opposed to resistance. Leaf movement like this may also increase turbulence of the air close to the surface of the leaf, which thins the boundary layer of air immediately adjacent to the surface, increasing the capacity for gas and heat exchange, as well as photosynthesis. Strong wind forces may result in diminished leaf number and surface area, which while reducing drag, involves a trade off of also reducing photosynthesis. Thus, leaf design may involve compromise between carbon gain, thermoregulation and water loss on the one hand, and the cost of sustaining both static and dynamic loads. In vascular plants, perpendicular forces are spread over a larger area and are relatively flexible in both bending and torsion, enabling elastic deforming without damage.[48]
Many leaves rely on hydrostatic support arranged around a skeleton of vascular tissue for their strength, which depends on maintaining leaf water status. Both the mechanics and architecture of the leaf reflect the need for transportation and support. Read and Stokes (2006) consider two basic models, the "hydrostatic" and "I-beam leaf" form (see Fig 1).[48] Hydrostatic leaves such as in Prostanthera lasianthos are large and thin, and may involve the need for multiple leaves rather single large leaves because of the amount of veins needed to support the periphery of large leaves. But large leaf size favors efficiency in photosynthesis and water conservation, involving further trade offs. On the other hand, I-beam leaves such as Banksia marginata involve specialized structures to stiffen them. These I-beams are formed from bundle sheath extensions of sclerenchyma meeting stiffened sub-epidermal layers. This shifts the balance from reliance on hydrostatic pressure to structural support, an obvious advantage where water is relatively scarce.[48] Long narrow leaves bend more easily than ovate leaf blades of the same area. Monocots typically have such linear leaves that maximize surface area while minimising self-shading. In these a high proportion of longitudinal main veins provide additional support.[48]
Interactions with other organisms
[edit]
Although not as nutritious as other organs such as fruit, leaves provide a food source for many organisms. The leaf is a vital source of energy production for the plant, and plants have evolved protection against animals that consume leaves, such as tannins, chemicals which hinder the digestion of proteins and have an unpleasant taste. Animals that are specialized to eat leaves are known as folivores.
Some species have cryptic adaptations by which they use leaves in avoiding predators. For example, the caterpillars of some leaf-roller moths will create a small home in the leaf by folding it over themselves. Some sawflies similarly roll the leaves of their food plants into tubes. Females of the Attelabidae, so-called leaf-rolling weevils, lay their eggs into leaves that they then roll up as means of protection. Other herbivores and their predators mimic the appearance of the leaf. Reptiles such as some chameleons, and insects such as some katydids, also mimic the oscillating movements of leaves in the wind, moving from side to side or back and forth while evading a possible threat.
Seasonal leaf loss
[edit]
Leaves in temperate, boreal, and seasonally dry zones may be seasonally deciduous (falling off or dying for the inclement season). This mechanism to shed leaves is called abscission. When the leaf is shed, it leaves a leaf scar on the twig. In cold autumns, they sometimes change color, and turn yellow, bright-orange, or red, as various accessory pigments (carotenoids and xanthophylls) are revealed when the tree responds to cold and reduced sunlight by curtailing chlorophyll production. Red anthocyanin pigments are now thought to be produced in the leaf as it dies, possibly to mask the yellow hue left when the chlorophyll is lost—yellow leaves appear to attract herbivores such as aphids.[49] Optical masking of chlorophyll by anthocyanins reduces risk of photo-oxidative damage to leaf cells as they senesce, which otherwise may lower the efficiency of nutrient retrieval from senescing autumn leaves.[50]
Evolutionary adaptation
[edit]
In the course of evolution, leaves have adapted to different environments in the following ways:[citation needed]
- Waxy micro- and nanostructures on the surface reduce wetting by rain and adhesion of contamination (See Lotus effect).
- Divided and compound leaves reduce wind resistance and promote cooling.
- Hairs on the leaf surface trap humidity in dry climates and create a boundary layer reducing water loss.
- Waxy plant cuticles reduce water loss.
- Large surface area provides a large area for capture of sunlight.
- In harmful levels of sunlight, specialized leaves, opaque or partly buried, admit light through a translucent leaf window for photosynthesis at inner leaf surfaces (e.g. Fenestraria).
- Kranz leaf anatomy in plants which perform C4 carbon fixation
- Succulent leaves store water and organic acids for use in CAM photosynthesis.
- Aromatic oils, poisons or pheromones produced by leaf borne glands deter herbivores (e.g. eucalypts).
- Inclusions of crystalline minerals deter herbivores (e.g. silica phytoliths in grasses, raphides in Araceae).
- Petals attract pollinators.
- Spines protect the plants from herbivores (e.g. cacti).
- Stinging hairs to protect against herbivory, e.g. in Urtica dioica and Dendrocnide moroides (Urticaceae).
- Special leaves on carnivorous plants are adapted for trapping food, mainly invertebrate prey, though some species trap small vertebrates as well (see carnivorous plants).
- Bulbs store food and water (e.g. onions).
- Tendrils allow the plant to climb (e.g. peas).
- Bracts and pseudanthia (false flowers) replace normal flower structures when the true flowers are greatly reduced (e.g. spurges, spathes in the Araceae and floral heads in the Asteraceae).
Terminology
[edit]
Shape
[edit]
Edge (margin)
[edit]The edge or margin is the outside perimeter of a leaf. The terms are interchangeable.
| Image | Term | Latin | Description |
|---|---|---|---|
| Entire | Forma integra | Even; with a smooth margin; without toothing | |
| Ciliate | ciliatus | Fringed with hairs | |
| Crenate | crenatus | Wavy-toothed; dentate with rounded teeth | |
| crenulate | crenulatus | Finely crenate | |
| crisped | crispus | Curly | |
| Dentate | dentatus | Toothed; may be coarsely dentate, having large teeth or glandular dentate, having teeth which bear glands | |
| Denticulate | denticulatus | Finely toothed | |
| Doubly serrate | duplicato-dentatus | Each tooth bearing smaller teeth | |
| Serrate | serratus | Saw-toothed; with asymmetrical teeth pointing forward | |
| Serrulate | serrulatus | Finely serrate | |
| Sinuate | sinuosus | With deep, wave-like indentations; coarsely crenate | |
| Lobate | lobatus | Indented, with the indentations not reaching the center | |
| Undulate | undulatus | With a wavy edge, shallower than sinuate | |
| Spiny or pungent | spiculatus | With stiff, sharp points such as thistles |
Apex (tip)
[edit]| Image | Term | Latin | Description |
|---|---|---|---|
| Acuminate | _ | Long-pointed, prolonged into a narrow, tapering point in a concave manner | |
| Acute | _ | Ending in a sharp, but not prolonged point | |
| Cuspidate | _ | With a sharp, elongated, rigid tip; tipped with a cusp | |
| Emarginate | _ | Indented, with a shallow notch at the tip | |
| Mucronate | _ | Abruptly tipped with a small short point | |
| Mucronulate | _ | Mucronate, but with a noticeably diminutive spine | |
| Obcordate | _ | Inversely heart-shaped | |
| Obtuse | _ | Rounded or blunt | |
| Truncate | _ | Ending abruptly with a flat end |
Base
[edit]- Acuminate
- Coming to a sharp, narrow, prolonged point.
- Acute
- Coming to a sharp, but not prolonged point.
- Auriculate
- Ear-shaped.
- Cordate
- Heart-shaped with the notch towards the stalk.
- Cuneate
- Wedge-shaped.
- Hastate
- Shaped like an halberd and with the basal lobes pointing outward.
- Oblique
- Slanting.
- Reniform
- Kidney-shaped but rounder and broader than long.
- Rounded
- Curving shape.
- Sagittate
- Shaped like an arrowhead and with the acute basal lobes pointing downward.
- Truncate
- Ending abruptly with a flat end, that looks cut off.
Surface
[edit]
The leaf surface is also host to a large variety of microorganisms; in this context it is referred to as the phyllosphere.
- Lepidote
- Covered with fine scurfy scales.
Hairiness
[edit]


"Hairs" on plants are properly called trichomes. Leaves can show several degrees of hairiness. The meaning of several of the following terms can overlap.
- Arachnoid, or arachnose
- With many fine, entangled hairs giving a cobwebby appearance.
- Barbellate
- With finely barbed hairs (barbellae).
- Bearded
- With long, stiff hairs.
- Bristly
- With stiff hair-like prickles.
- Canescent
- Hoary with dense grayish-white pubescence.
- Ciliate
- Marginally fringed with short hairs (cilia).
- Ciliolate
- Minutely ciliate.
- Floccose
- With flocks of soft, woolly hairs, which tend to rub off.
- Glabrescent
- Losing hairs with age.
- Glabrous
- No hairs of any kind present.
- Glandular
- With a gland at the tip of the hair.
- Hirsute
- With rather rough or stiff hairs.
- Hispid
- With rigid, bristly hairs.
- Hispidulous
- Minutely hispid.
- Hoary
- With a fine, close grayish-white pubescence.
- Lanate, or lanose
- With woolly hairs.
- Pilose
- With soft, clearly separated hairs.
- Puberulent, or puberulous
- With fine, minute hairs.
- Pubescent
- With soft, short and erect hairs.
- Scabrous, or scabrid
- Rough to the touch.
- Sericeous
- Silky appearance through fine, straight and appressed (lying close and flat) hairs.
- Silky
- With adpressed, soft and straight pubescence.
- Stellate, or stelliform
- With star-shaped hairs.
- Strigose
- With appressed, sharp, straight and stiff hairs.
- Tomentose
- Densely pubescent with matted, soft white woolly hairs.
- Cano-tomentose
- Between canescent and tomentose.
- Felted-tomentose
- Woolly and matted with curly hairs.
- Tomentulose
- Minutely or only slightly tomentose.
- Villous
- With long and soft hairs, usually curved.
- Woolly
- With long, soft and tortuous or matted hairs.
Timing
[edit]- Hysteranthous
- Developing after the flowers [51]
- Synanthous
- Developing at the same time as the flowers [52]
Venation
[edit]Classification
[edit]





A number of different classification systems of the patterns of leaf veins (venation or veination) have been described,[29] starting with Ettingshausen (1861),[53] together with many different descriptive terms, and the terminology has been described as "formidable".[29] One of the commonest among these is the Hickey system, originally developed for "dicotyledons" and using a number of Ettingshausen's terms derived from Greek (1973–1979):[54][55][56] (see also: Simpson Figure 9.12, p. 468)[29]
Hickey system
[edit]- 1. Pinnate (feather-veined, reticulate, pinnate-netted, penniribbed, penninerved, or penniveined)
- The veins arise pinnately (feather like) from a single primary vein (mid-vein) and subdivide into secondary veinlets, known as higher order veins. These, in turn, form a complicated network. This type of venation is typical for (but by no means limited to) "dicotyledons" (non monocotyledon angiosperms). E.g., Ostrya. There are three subtypes of pinnate venation:
- Craspedodromous (Greek: kraspedon – edge, dromos – running)
- The major veins reach to the margin of the leaf.
- Camptodromous
- Major veins extend close to the margin, but bend before they intersect with the margin.
- Hyphodromous
- All secondary veins are absent, rudimentary or concealed
These in turn have a number of further subtypes such as eucamptodromous, where secondary veins curve near the margin without joining adjacent secondary veins.



- 2. Параллелодромные (параллельножилковые, параллельноребристые, параллельнонервные, пеннипараллельные, полосатые).
- Две или несколько первичных жилок, начинающихся рядом друг с другом у основания листа, идущих параллельно друг другу до верхушки, а затем сходящихся там. Комиссуральные вены (мелкие вены) соединяют большие параллельные вены. Типично для большинства однодольных растений , например трав . Также используются дополнительные термины маргинальный (первичные жилки достигают края) и сетчатый (сетчатый).

- 3. Кампилодромный ( campylos – изогнутый)
- Несколько первичных жилок или ветвей, берущих начало в одной точке или вблизи нее и идущих изогнутыми дугами, а затем сходящихся на вершине. Например, Майантемум .

- 4. Акродромный
- Две или более первичных или хорошо развитые вторичные жилки в сходящихся дугах к вершине, без базальной кривизны, как у кампилодромного. Могут быть базальными или супрабазальными в зависимости от происхождения, совершенными или несовершенными в зависимости от того, достигают ли они 2/3 длины вершины. Например, Miconia (базальный тип), Endlicheria (супрабазальный тип).




- 5. Актинодромный
- Три и более первичных жилок, радиально расходящихся из одной точки. Например, Arcangelisia (базальный тип), Givotia (супрабазальный тип).


- 6. Палинактодромный
- Первичные жилки с одной или несколькими точками вторичного дихотомического ветвления за пределами первичного расхождения, расположенные близко или более удаленно. Например, Платан .


Типы 4–6 аналогичным образом можно подразделить на базальные (первичные первичные волокна соединяются у основания лопасти) или супрабазальные (расходящиеся над основанием лопасти), а также совершенные или несовершенные, а также флебеллатные.
Примерно в то же время Мелвилл (1976) описал систему, применимую ко всем покрытосеменным растениям, используя латинскую и английскую терминологию. [57] В Мелвилле также было шесть подразделений в зависимости от порядка развития вен.
- Арбускулярный (arbuscularis)
- Многократное ветвление с помощью регулярной дихотомии с образованием трехмерной кустообразной структуры, состоящей из линейных сегментов (2 подкласса)
- Флабеллат (флабеллат)
- Первичные жилки прямые или лишь слегка изогнутые, веерообразно расходящиеся от основания (4 подкласса).
- Пальматус (пальматус)
- Изогнутые первичные вены (3 подкласса)
- Перистый (перистый)
- Единственная первичная жилка, средняя жилка, вдоль которой через более или менее равные промежутки расположены прямые или дугообразные вторичные жилки (6 подклассов).
- Коллимат (коллимат)
- Многочисленные продольно-параллельные первичные жилки, отходящие от поперечной меристемы (5 подклассов).
- Конглютинат (conglutinatus)
- Произведено от сросшихся перистых листочков (3 подкласса).
Модифицированная форма системы Хики была позже включена в Смитсоновскую классификацию (1999), которая предложила семь основных типов жилкования, основанных на архитектуре первичных жилок, с добавлением Flabellate в качестве дополнительного основного типа. Затем была проведена дальнейшая классификация на основе вторичных жил с еще 12 типами, такими как;
- Брохидодромный
- Закрытая форма, в которой второстепенные ветви соединены в ряд выступающих арок, как у Хильдегардии .
- Краспедодромный
- Открытая форма с вторичными ветвями, заканчивающимися по краю зубчатыми листьями, как у Celtis .
- эвкамптодромный
- Промежуточная форма с перевернутыми вторичными ветвями, которые постепенно уменьшаются на вершине, но внутри края, и соединены промежуточными третичными жилками, а не петлями между вторичными ветвями, как у Cornus .
- кладодромный
- Вторичные ветви свободно ветвятся к краю, как у Rhus .
термины, которые использовались в качестве подтипов в исходной системе Хики. [58]





Хильдегардия мигеоди

Западные кельты

Корнус лекарственный

Русус яйцевидный
Дальнейшие описания включали жилки более высокого порядка или второстепенные жилки и узоры ареол ( см. Рабочую группу по архитектуре листьев, рисунки 28–29). [58]

- Фантастически
- От нескольких до многих одинаковых тонких базальных жилок, расходящихся радиально под небольшими углами и ветвящихся на вершине. Например, Параномус .

При анализе структуры жилок часто учитываются порядки жилок, тип первичных жил, тип вторичных жил (основные жилы) и плотность второстепенных вен. Ряд авторов приняли упрощенные варианты этих схем. [59] [29] В простейшем случае основные типы жилок можно разделить на три или четыре группы в зависимости от рассматриваемых подразделений растения;
- перистый
- пальмовидный
- параллельный
где пальчатая относится к множеству первичных жилок, отходящих от черешка, в отличие от ответвлений от центральной главной жилки в перистой форме, и включает оба типа засоса 4 и 5, которые сохраняются как подтипы; например, пальчато-акродромный ( см. Справочник Службы национальных парков). [60]

- Пальчато-сетчатые, пальчато-сетчатые, пальчато-жилковые, веерообразно-жилковые.
- Несколько основных жилок примерно одинакового размера расходятся от общей точки у основания листа, где прикрепляется черешок, и расходятся к краю листа. Листья с пальчатыми жилками часто лопастные или разделенные, лопасти расходятся от общей точки. Они могут различаться по числу первичных жилок (3 и более), но всегда расходятся из общей точки. [61] например, большинство Acer (клены).

Другие системы
[ редактировать ]Альтернативно, Симпсон использует: [29]
- Хладнокровный
- Центральная средняя жилка без боковых жилок ( микрофиллезная ), наблюдается у несемянных трахеофитов , таких как хвощи.
- дихотомический
- Вены последовательно разветвляются на вены одинакового размера из общей точки, образуя Y-образное соединение и расходясь веером. Среди древесных растений умеренного пояса гинкго двулопастный — единственный вид с дихотомическим жилкованием. Также некоторые птеридофиты (папоротники). [61]
- Параллельно
- Первичные и вторичные жилки примерно параллельны друг другу, проходят по всей длине листа, часто соединены короткими перпендикулярными звеньями, а не образуют сети. У некоторых видов параллельные жилки соединяются у основания и вершины, например, у игольчатых вечнозеленых растений и трав. Характерно для однодольных, но исключения включают Arisaema и, как показано ниже, сетчатую. [61]
- Сетчатые (сетчатые, перистые)
- Выраженная средняя жилка, по обеим сторонам которой отходят вторичные жилки. Название происходит от конечных прожилок, которые образуют взаимосвязанную сеть, подобную узору или сети. (Первичное и вторичное жилкование можно назвать перистым, а сетчатое, как и более тонкие жилки, - сетчатым или сетчатым); большинство неоднодольных покрытосеменных, за исключением Calophyllum . Некоторые однодольные имеют сетчатое жилкование, в том числе Colocasia , Dioscorea и Smilax . [61]

Редуцированные микрофилловые листья (L), отходящие в мутовке от узла.

Дихотомическое жилкование
Однако эти упрощенные системы допускают дальнейшее разделение на несколько подтипов. Симпсон, [29] (и другие) [62] разделяет параллельные и сетчатые (некоторые используют только эти два термина для покрытосеменных) [63] по числу первичных жилок (коста) следующим образом;
- Параллельно
- Penni-parallel (pinnate, pinnate parallel, unicostate parallel)
- Единственная центральная выступающая средняя жилка, отходящие от нее вторичные вены отходят перпендикулярно ей и идут параллельно друг другу по направлению к краю или кончику, но не соединяются (анастомозируют). Термин однокостный относится к выступающей единственной средней жилке (косте), проходящей по всей длине листа от основания до вершины. например, Zingiberales , такие как бананы и т. д.
- Пальчато-параллельный (многосостоятельный параллельный)
- Несколько одинаково выраженных первичных жилок, исходящих из одной точки у основания и идущих параллельно кончику или краю. Термин «мультикостный» относится к наличию более одной выступающей главной вены. например, «веерные» (пальчатые) пальмы (Arecaceae)
- Многоуровневая параллельная конвергенция
- Средние жилки сходятся на вершине, например, Bambusa arundinacea = B. bambos (Aracaceae), Eichornia.
- Многосостояние параллельное расхождение
- Средние жилки расходятся более или менее параллельно к краю, например, Borassus (Poaceae), веерные пальмы.
- Сетчатый (сетчатый)
- Перистолистные (жилковые, сетчатые, одностворчатые).
- Одна выступающая средняя жилка, идущая от основания к вершине, вторичные жилки, отходящие с обеих сторон по длине первичной средней жилки, идущие к краю или вершине (кончику), с сетью более мелких жилок, образующих сетку (сетку или сеть). например Mangifera , Ficus religiosa , Psidium guajava , Hibiscus rosa-sinensis , Salix alba.
- Пальчатые (многодольные сетчатые)
- Несколько первичных жилок, исходящих из одной точки и идущих от основания к вершине. например, Liquidambar styraciflua. Его можно подразделить дальше;
- Мультикостный конвергентный
- Основные жилки расходятся от начала основания, затем сходятся к кончику. например , Зизифус , Смилакс , Корица
- Многосостояние расходящееся
- Все основные жилки расходятся к кончику. например , Gossypium , Cucurbita , Carica papaya , Ricinus communis
- Тройчатые (тройчато-сетчатые)
- Три первичные жилки, как указано выше, например ( см .) Ceanothus leucodermis , [64] C. tomentosus , [65] Энселия Фаринаса


Пальчато-параллельный

Многоуровневая параллельная конвергенция

Многосостояние параллельное расхождение
Перисто сетчатый

Пальчато-сетчатый

Многореберная пальчатая конвергенция

Многореберные пальчаторасходящиеся
Эти сложные системы мало используются в морфологическом описании таксонов, но полезны при идентификации растений. [29] хотя его критиковали за чрезмерное обременение жаргоном. [66]
Более старая и еще более простая система, используемая в некоторых растениях. [67] использует только две категории: открытую и закрытую.
- Открытый: жилки более высокого порядка имеют свободные окончания среди клеток и более характерны для неоднодольных покрытосеменных растений. Они чаще связаны с формой листьев: зубчатыми, лопастными или сложными. Их можно подразделить на;
- Перистые (перисто-жилковые) листья с главной центральной жилкой или ребром (средней жилкой), от которого отходит остальная жилковая система.
- Пальчатые, у которых три и более основных ребра поднимаются вместе у основания листа и расходятся вверх.
- Дихотомический, как у папоротников, где жилки многократно разветвляются.
- Закрытый: жилки высшего порядка соединены петлями, не заканчиваясь свободно между клетками. Обычно они имеют листья с гладкими очертаниями и характерны для однодольных растений.
- Их можно подразделить по тому, идут ли жилки параллельно, как у трав, или имеют другой рисунок.
Другие описательные термины
[ редактировать ]Существует также множество других описательных терминов, часто очень специализированных и привязанных к конкретным таксономическим группам. [68] Выраженность вен зависит от ряда особенностей. К ним относятся ширина вен, их выступание по отношению к поверхности пластинки и степень непрозрачности поверхности, которая может скрывать более мелкие вены. В связи с этим вены называются неясными , а порядок скрытых вен и верхняя, нижняя или обе поверхности уточняются дополнительно. [69] [61]
Термины, описывающие выступающие вены, включают буллезную , желобчатую , плоскую , желобчатую , вдавленную , выступающую и утопленную ( рис . 6.1 Hawthorne & Lawrence 2013). [66] [70] На разных участках листа жилки могут проявляться по-разному. Например, Pimenta Racemosa имеет желобчатую среднюю жилку на верхней поверхности, но она заметна на нижней поверхности. [66]
Описание выступания вен:
- Буллатировать
- Поверхность листа приподнята в виде ряда куполов между жилками на верхней поверхности и поэтому также с выраженными углублениями. например, Rytigynia pauciflora , [71] Винная лоза
- Направленный (каналикулирующий)
- Вены погружаются под поверхность, образуя округлый канал. Иногда путают с «желобами», потому что каналы могут функционировать как желоба для стока дождя и обеспечения высыхания, как у многих Melastomataceae . [72] например ( см .) перец кистевидный (Myrtaceae), [73] Клидемия гирта (Melastomataceae).
- с желобами
- Жилки частично выступающие, гребень над поверхностью листовой пластинки, но с каналами, идущими вдоль каждой стороны, как желоба.
- Впечатленный
- Жилка образует приподнятую линию или гребень, лежащий ниже плоскости несущей ее поверхности, как бы вдавленной в нее, и часто обнажается на нижней поверхности. Ткани возле вен часто сморщиваются, придавая им впалый или рельефный вид.
- Затемнять
- Вены не видны или совсем нечеткие; если не указано, то не видно невооруженным глазом. например, Berberis gagnepainii . У этого барбериса жилки неясны только на нижней поверхности. [74]
- Видный
- Вена приподнята над окружающей поверхностью, чтобы ее можно было легко прощупать пальцем. например ( см .) Пимента кистевидная , [73] Спатифиллум каннифолиум [75]
- Утопленный
- Вена погружена под поверхность, более выражена, чем окружающие ткани, но более углублена в русле, чем при вдавленных венах. например, Калина складчатая .


Неясный (под поверхностью)

Видный

Утопленный
Описание других особенностей:
- Плинервий (плинервед)
- Более одной главной вены (нерва) у основания. Боковые вторичные жилки ветвятся от точки над основанием листа. Обычно выражается суффиксом , например, в трех- или трехслойном листе. В трехслойном (триплинервном) листе три основные жилки разветвляются над основанием пластинки (две вторичные жилки и главная жилка) и впоследствии идут по существу параллельно, как у Ceanothus и у Celtis . Точно так же пятижилковый (пятижилковый) лист имеет четыре второстепенные жилки и одну главную жилку. особенно заметен рисунок из 3–7 жилок У Melastomataceae . Этот термин также использовался в Vaccinieae . Этот термин использовался как синоним акродромного, пальчато-акродромного или супрабазального акродромного, и считается, что он имеет слишком широкое определение. [76] [76]
- Лестничная форма
- Вены расположены как ступеньки лестницы, особенно вены более высокого порядка.
- субмаргинальный
- Жилки идут близко к краю листа.
- Тринервед
- 2 основных базальных нерва, кроме среднего ребра
Схемы жилкования
[ редактировать ]| Изображение | Срок | Описание |
|---|---|---|
 | дугообразный | Вторичное выгибание к вершине |
 | дихотомический | Вены разделяются на две части |
 | Продольный | Все вены расположены преимущественно вдоль средней вены. |
 | Параллельно | Все вены параллельны и не пересекаются. |
 | Перистый | Вторичные вены, отходящие от средней жилки |
 | сетчатый | Все жилки многократно ветвятся, сетчатые. |
 | Поворот | Жилки идут от центра листа и расходятся к краям. |
 | поперечный | Третичные вены идут перпендикулярно оси главной вены, соединяя вторичные вены. |
Размер
[ редактировать ]Термины мегафилл , макрофилл , мезофилл , нотофилл , микрофилл , нанофилл и лептофилл используются для описания размеров листьев (в порядке убывания) в классификации, разработанной в 1934 году Кристеном К. Раункером и с тех пор модифицированной другими. [77] [78]
См. также
[ редактировать ]- Глоссарий морфологии листьев
- Глоссарий морфологии растений § Листья
- Корона (ботаника)
- Эволюционная история листьев
- Эволюционное развитие листьев
- Индекс площади листьев
- Концентрат листового белка
- Датчик листьев – устройство, измеряющее уровень влажности в листьях растений.
- Форма листа
- Вернация – прорастание листьев, а также расположение листьев в бутоне.
- гладкий лист как музыкальный инструмент
Ссылки
[ редактировать ]- ^ Jump up to: а б Исав 2006 .
- ^ Главный 1953 год .
- ^ Jump up to: а б Маусет 2009 .
- ^ «Стреляющая система» . Словарь ботанической терминологии . Питомник искусств кактусов. nd Архивировано из оригинала 4 мая 2021 года . Проверено 4 мая 2021 г.
- ^ Джеймс и др. 1999 .
- ^ Jump up to: а б с д и Стюарт и Ротвелл 1993 .
- ^ Куни-Советтс и Саттлер, 1987 .
- ^ Цукая 2013 .
- ^ Фейжер 2006 .
- ^ Перселл 2016 .
- ^ Виллерт и др. 1992 .
- ^ Байер 1982 .
- ^ Марлот 1913–1932 .
- ^ Jump up to: а б с Симпсон 2011 , с. 356.
- ^ Крог 2010 .
- ^ Джеймс и Белл 2000 .
- ^ Хейвуд и др. 2007 .
- ^ Симпсон 2011 , стр. 356–357.
- ^ Халле 1977 .
- ^ Jump up to: а б с Иллюстрированная ботаника: знакомство с растениями. Основные группы, семейства цветковых растений . Томсон Сайенс. 1984. с. 21.
- ^ Дидье Рейнхардт и Крис Кулемайер, «Филлотаксис у высших растений», в книге Майкла Т. Макмануса, Брюса Вейта, ред., Меристематические ткани в росте и развитии растений , январь 2002 г., ISBN 978-1-84127-227-6 , Уайли-Блэквелл.
- ^ Коксетер Х.С. (1961). Введение в геометрию . Уайли. п. 169.
- ^ Рейнхардт и Кулемайер, стр. 175
- ^ Ёнекура, Такааки; Ивамото, Акитоши; Фудзита, Хиронори; Сугияма, Мунетака (6 июня 2019 г.). Умулис, Дэвид (ред.). «Математические модельные исследования комплексного формирования основных и второстепенных филлотаксисов у растений с преобладающим акцентом на филлотаксис orixate» . PLOS Вычислительная биология . 15 (6): e1007044. Бибкод : 2019PLSCB..15E7044Y . дои : 10.1371/journal.pcbi.1007044 . ISSN 1553-7358 . ПМК 6553687 . ПМИД 31170142 .
- ^ Роллан-Лаган и др. 2009 .
- ^ Jump up to: а б с Стены 2011 .
- ^ Jump up to: а б с Дикисон 2000 .
- ^ Jump up to: а б Рудалль 2007 .
- ^ Jump up to: а б с д и ж г час я Симпсон 2011 , Жилкование листьев, стр. 465–468.
- ^ Jump up to: а б с д Сак и Скоффони, 2013 .
- ^ Jump up to: а б с Рот-Небельсик и др . , 2001 г.
- ^ Уэно и др. 2006 .
- ^ Рунионс и др. 2005 .
- ^ Мэсси и Мерфи 1996 , Текстура поверхностного жилкования
- ^ Багчи и др. 2016 .
- ^ Кот 2009 .
- ^ Клементс 1905 .
- ^ Кудер и др. 2002 .
- ^ Корсон и др. 2009 .
- ^ Лагуна и др. 2008 .
- ^ Арбер 1950 .
- ^ Рутисхаузер и Саттлер 1997 .
- ^ Лакруа и др. 2003 .
- ^ Экардт и Баум 2010 .
- ^ Пуаре, Ришар; Визе-Клинкенберг, Аника; Родитель Борис; Мелевчик, Майкл; Шурр, Ульрих; Тардье, Франсуа; Уолтер, Ахим (2010). «Суточная динамика роста листьев у однодольных и двудольных видов: эндогенные ритмы и температурные эффекты» . Журнал экспериментальной ботаники . 61 (6): 1751–1759. дои : 10.1093/jxb/erq049 . ISSN 1460-2431 . ПМЦ 2852670 . ПМИД 20299442 .
- ^ Мелевчик, Майкл; Фридли, Майкл; Кирхгесснер, Норберт; Уолтер, Ахим (25 июля 2013 г.). «Дневной рост листьев сои: новый метод анализа двумерного расширения листьев с высоким временным разрешением на основе подхода отслеживания маркеров (Martrack Leaf)» . Растительные методы . 9 (1): 30. дои : 10.1186/1746-4811-9-30 . hdl : 20.500.11850/76534 . ISSN 1746-4811 . ПМЦ 3750653 . ПМИД 23883317 .
- ^ Фридли, Майкл; Уолтер, Ахим (2015). «Суточная динамика роста молодых листочков сои (G lycine max) синхронна в разных положениях на растении» . Растение, клетка и окружающая среда . 38 (3): 514–524. дои : 10.1111/шт.12407 . ISSN 0140-7791 . ПМИД 25041284 .
- ^ Jump up to: а б с д и Рид и Стоукс 2006 .
- ^ Доринг и др. 2009 .
- ^ Фейлд и др. 2001 .
- ^ «Глоссарий Кью – определение истеричного состояния» . 3 декабря 2013. Архивировано из оригинала 3 декабря 2013 года . Проверено 12 мая 2017 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ «Глоссарий Кью - определение синантуса» . 3 декабря 2013. Архивировано из оригинала 3 декабря 2013 года . Проверено 12 мая 2017 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ Эттингсгаузен 1861 .
- ^ Хикки 1973 .
- ^ Хики и Вулф 1975 .
- ^ Хикки 1979 .
- ^ Мелвилл 1976 .
- ^ Jump up to: а б Рабочая группа по листовой архитектуре, 1999 .
- ^ Джадд и др. 2007 .
- ^ Флориссан Leaf Key 2016 .
- ^ Jump up to: а б с д и Клинг и др. 2005 , Жилкование листьев.
- ^ Берг 2007 .
- ^ Морфология покрытосеменных 2017 , Жилкование.
- ^ Симпсон 2017 , Ceanothus leucodermis
- ^ Симпсон 2017 , Ceanothus tomentosus
- ^ Jump up to: а б с Hawthorne & Lawrence 2013 , Жилкование листьев, стр. 135–136.
- ^ Каллен и др. 2011 .
- ^ Неотропики 2017 .
- ^ Оксфордский глоссарий гербариев, 2017 г ..
- ^ Оксфордский глоссарий гербариев, 2017 г. , Выдающееся положение вен.
- ^ Вердкорт и Бридсон 1991 .
- ^ Hemsley & Poole 2004 , Морфология листьев и сушка, с. 254
- ^ Jump up to: а б Хьюз 2017 , Пимента кистевидная
- ^ Каллен и др. 2011 , Berberis gagnepainii vol. 2 р. 398
- ^ Квантлен 2015 , Spathiphyllum cannifolium
- ^ Jump up to: а б Педраса-Пеньалоса 2013 .
- ^ Уиттен и др. 1997 .
- ^ Уэбб, Лен (1 октября 1959 г.). «Физиономическая классификация тропических лесов Австралии». Журнал экологии . 47 (3). Британское экологическое общество: Журнал экологии Vol. 47, № 3, стр. 551–570: 555. Бибкод : 1959JEcol..47..551W . дои : 10.2307/2257290 . JSTOR 2257290 .
Библиография
[ редактировать ]Книги и главы
[ редактировать ]- Арбер, Агнес (1950). Естественная философия растительной формы . Архив Кубка . GGKEY:HCBB8RZREL4.
- Байер, МБ (1982). Новый справочник по Хавортии . Кирстенбош: Национальный ботанический сад Южной Африки . ISBN 978-0-620-05632-8 . Архивировано из оригинала 06 сентября 2023 г. Проверено 25 июля 2018 г.
- Берг, Линда (23 марта 2007 г.). Вводная ботаника: растения, люди и окружающая среда, издание для СМИ . Cengage Обучение. ISBN 978-1-111-79426-2 .
- Каллен, Джеймс; Колени, Сабина Г.; Кьюби, Х. Сюзанна Кьюби, ред. (2011) [1984–2000]. Европейская садовая флора, Цветковые растения: Руководство по идентификации растений, выращиваемых в Европе как на открытом воздухе, так и под стеклом. 5 томов (2-е изд.). Кембридж: Издательство Кембриджского университета . Архивировано из оригинала 28 декабря 2016 г. Проверено 8 марта 2017 г.
- Каттер, Э. Г. (1969). Анатомия растений, эксперимент и интерпретация. Часть 2. Органы . Лондон: Эдвард Арнольд. п. 117. ИСБН 978-0-7131-2302-9 .
- Дикисон, Уильям К. (2000). Интегративная анатомия растений . Академическая пресса . ISBN 978-0-08-050891-7 .
- Исав, Кэтрин (2006) [1953]. Эверт, Рэй Ф. (ред.). Анатомия растений Исава: меристемы, клетки и ткани тела растения: их структура, функции и развитие (3-е изд.). John Wiley & Sons Inc. Нью-Йорк: ISBN 978-0-470-04737-8 . Архивировано из оригинала 06 сентября 2023 г. Проверено 2 сентября 2017 г.
- Эттингсгаузен, К. (1861). Скелеты листьев двудольных растений с особым упором на изучение и идентификацию остатков ископаемых растений . Вена: Классификация архитектуры двудольных.
- Хаупт, Артур Винг (1953). Морфология растений . МакГроу-Хилл .
- Хоторн, Уильям; Лоуренс, Анна (2013). Идентификация растений: создание удобных для пользователя полевых руководств по управлению биоразнообразием . Рутледж. ISBN 978-1-136-55972-3 .
- Хемсли, Алан Р.; Пул, Имоджен, ред. (2004). Эволюция физиологии растений . Академическая пресса . ISBN 978-0-08-047272-0 .
- Хейвуд, Вирджиния ; Браммитт, РК; Калхэм, А .; Себерг, О. (2007). Семейства цветковых растений мира . Нью-Йорк: Книги Светлячка. п. 287. ИСБН 978-1-55407-206-4 .
- Хикки, ЖЖ. Пересмотренная классификация строения двудольных листьев . стр. я 5–39. , в Меткалф и Мел (1979)
- Джадд, Уолтер С .; Кэмпбелл, Кристофер С.; Келлог, Элизабет А.; Стивенс, Питер Ф .; Донохью, Майкл Дж. (2007) [1-е изд. 1999, 2-е 2002]. Систематика растений: филогенетический подход (3-е изд.). Синауэр Ассошиэйтс. ISBN 978-0-87893-407-2 . Архивировано из оригинала 06 сентября 2023 г. Проверено 2 сентября 2017 г.
- Крог, Дэвид (2010), Биология: Путеводитель по миру природы (5-е изд.), Издательство Benjamin-Cummings Publishing Company, стр. 463, ISBN 978-0-321-61655-5 , заархивировано из оригинала 24 января 2023 г. , получено 24 мая 2016 г.
- Рабочая группа по листовой архитектуре (1999). Руководство по архитектуре листа - морфологическое описание и классификация двудольных и однодольных покрытосеменных с сетчатыми жилками (PDF) . Смитсоновский институт . ISBN 978-0-9677554-0-3 . Архивировано (PDF) из оригинала 20 октября 2016 г. Проверено 15 февраля 2017 г.
- Марлот, Рудольф (1913–1932). Флора Южной Африки: с синопическими таблицами родов высших растений. 6 томов . Кейптаун: Darter Bros. & Co. Архивировано из оригинала 6 сентября 2023 г. Проверено 27 августа 2020 г.
- Маусет, Джеймс Д. (2009). Ботаника: введение в биологию растений (4-е изд.). Садбери, Массачусетс: Издательство Jones and Bartlett. ISBN 978-0-7637-5345-0 .
- Меткалф, Чехия; Мел, Л., ред. (1979) [1957]. Анатомия двудольных: листья, стебель и древесина в связи с таксономией, с примечаниями об экономическом использовании. 2 тома (2-е изд.). Оксфорд: Кларендон Пресс. ISBN 978-0-19-854383-1 .
- Пранс, Гиллин Толми (1985). Листья: формирование, характеристики и использование сотен листьев, встречающихся во всех частях света . Фотографии Кьелла Б. Сандведа. Лондон: Темза и Гудзон. ISBN 978-0-500-54104-3 .
- Рудалл, Паула Дж. (2007). Анатомия цветковых растений: введение в строение и развитие (3-е изд.). Кембридж: Издательство Кембриджского университета . ISBN 978-0-521-69245-8 . Архивировано из оригинала 06 сентября 2023 г. Проверено 27 августа 2020 г.
- Симпсон, Майкл Г. (2011). Систематика растений . Академическая пресса. ISBN 978-0-08-051404-8 . Архивировано из оригинала 17 января 2023 г. Проверено 24 мая 2016 г.
- Стюарт, Уилсон Н; Ротвелл, Гар В. (1993) [1983]. Палеоботаника и эволюция растений (2-е изд.). Издательство Кембриджского университета . ISBN 978-0-521-38294-6 .
- Вердкур, Бернар; Бридсон, Дайан М. (1991). Флора тропической Восточной Африки - Rubiaceae Том 3 . ЦРК Пресс. ISBN 978-90-6191-357-3 .
- Уиттен, Тони; Соериатмаджа, Рохаят Эмон; Афифф, Сурая А. (1997). Экология Явы и Бали . Издательство Оксфордского университета. п. 505. ИСБН 978-962-593-072-5 . Архивировано из оригинала 06 сентября 2023 г. Проверено 27 августа 2020 г.
- Виллерт, Дитер Дж. фон; Эллер, Б.М.; Вергер, MJA; Бринкманн, Э; Иленфельдт, HD (1992). Жизненные стратегии суккулентов в пустынях: с особым упором на пустыню Намиб . Архив Кубка . ISBN 978-0-521-24468-8 .
Статьи и тезисы
[ редактировать ]- Багчи, Дебжани; Дасгупта, Авик; Гондалия, Амит Д.; Раджпут, Кишор С. (2016). «Идеи из мира растений: подход к фрактальному анализу для настройки механической жесткости матрицы каркаса в тонких пленках». Передовые исследования материалов . 1141 : 57–64. дои : 10.4028/www.scientific.net/AMR.1141.57 . S2CID 138338270 .
- Клементс, Эдит Шварц (декабрь 1905 г.). «Связь структуры листа с физическими факторами» . Труды Американского микроскопического общества . 26 : 19–98. дои : 10.2307/3220956 . JSTOR 3220956 . Архивировано из оригинала 4 августа 2023 г. Проверено 6 сентября 2023 г.
- Куни-Советс, К.; Саттлер, Р. (1987). «Развитие филлоклады у Asparagaceae: пример гомеоза». Ботанический журнал Линнеевского общества . 94 (3): 327–371. дои : 10.1111/j.1095-8339.1986.tb01053.x .
- Корсон, Фрэнсис; Адда-Бедиа, Мохтар; Будауд, Арезки (2009). «Сети жилкования листьев In silico: рост и реорганизация, вызванные механическими силами» (PDF) . Журнал теоретической биологии . 259 (3): 440–448. Бибкод : 2009JThBi.259..440C . дои : 10.1016/j.jtbi.2009.05.002 . ПМИД 19446571 . S2CID 25560670 . Архивировано из оригинала (PDF) 9 декабря 2017 г.
- Кот, Г.Г. (2009). «Разнообразие и распределение идиобластов, продуцирующих кристаллы оксалата кальция, у Dieffenbachia seguine (Araceae)» . Американский журнал ботаники . 96 (7): 1245–1254. дои : 10.3732/ajb.0800276 . ПМИД 21628273 .
- Кудер, Ю.; Пошар, Л.; Аллен, К.; Адда-Бедиа, М.; Дуади, С. (1 июля 2002 г.). «Жилкование листа, сформированное в тензорном поле» (PDF) . Европейский физический журнал Б. 28 (2): 135–138. Бибкод : 2002EPJB...28..135C . дои : 10.1140/epjb/e2002-00211-1 . S2CID 51687210 . Архивировано из оригинала (PDF) 9 декабря 2017 года.
- Дёринг, Т.Ф; Арчетти, М.; Харди, Дж. (7 января 2009 г.). «Осенние листья глазами травоядных» . Труды Королевского общества B: Биологические науки . 276 (1654): 121–127. дои : 10.1098/rspb.2008.0858 . ПМК 2614250 . ПМИД 18782744 .
- Экардт, Северная Каролина; Баум, Д. (20 июля 2010 г.). «Загадка подостемад: эволюция необычной морфологии Podostemaceae» . Растительная клетка онлайн . 22 (7): 2104. doi : 10.1105/tpc.110.220711 . ПМЦ 2929115 . ПМИД 20647343 .
- Фейжье, Франсуа (14 декабря 2006 г.). Модели формирования сосудистого рисунка листьев (кандидатская диссертация) . Парижский университет VI . Архивировано (PDF) из оригинала 7 марта 2017 года . Проверено 6 марта 2017 г.
- Филд, Т.С.; Ли, Д.В.; Холбрук, Нью-Мексико (1 октября 2001 г.). «Почему листья краснеют осенью. Роль антоцианов в старении листьев кизила красно-ивового» . Физиология растений . 127 (2): 566–574. дои : 10.1104/стр.010063 . ПМК 125091 . ПМИД 11598230 .
- Халле, Ф. (1977). «Самый длинный лист на ладони». Принципы . 21:18 .
- Хики, Лео Дж. (1 января 1973 г.). «Классификация строения двудольных листьев» (PDF) . Американский журнал ботаники . 60 (1): 17–33. дои : 10.2307/2441319 . JSTOR 2441319 . Архивировано (PDF) из оригинала 11 августа 2017 года . Проверено 14 февраля 2017 г.
- Хики, Лео Дж.; Вулф, Джек А. (1975). «Основы филогении покрытосеменных: вегетативная морфология» . Анналы ботанического сада Миссури . 62 (3): 538–589. дои : 10.2307/2395267 . JSTOR 2395267 . Архивировано из оригинала 31 октября 2022 г. Проверено 6 сентября 2023 г.
- Ингерсолл, Эрнест (1920). . Американская энциклопедия . Том. XVII.
- Джеймс, ЮАР; Белл, Д.Т. (2000). «Влияние доступности света на структуру листьев и рост двух сортов Eucalyptus globulus ssp. globulus » (PDF) . Физиология дерева . 20 (15): 1007–1018. дои : 10.1093/treephys/20.15.1007 . ПМИД 11305455 . Архивировано (PDF) из оригинала 1 апреля 2022 г. Проверено 10 мая 2019 г.
- Лакруа, К.; Жен, Б.; Перселл-Макдональд, С. (2003). «Сравнение побегов и сложных листьев эвдикотов: динамическая морфология как альтернативный подход» . Ботанический журнал Линнеевского общества . 143 (3): 219–230. дои : 10.1046/j.1095-8339.2003.00222.x . Архивировано из оригинала 22 июня 2020 г. Проверено 8 сентября 2019 г.
- Лагуна, Мария Ф.; Бон, Штеффен; Ягла, Эдуардо А.; Борн, Филип Э. (2008). «Роль упругих напряжений в морфогенезе жилкования листьев» . PLOS Вычислительная биология . 4 (4): e1000055. arXiv : 0705.0902 . Бибкод : 2008PLSCB...4E0055L . дои : 10.1371/journal.pcbi.1000055 . ПМК 2275310 . ПМИД 18404203 .
- Мелвилл, Р. (ноябрь 1976 г.). «Терминология листовой архитектуры». Таксон . 25 (5/6): 549–561. дои : 10.2307/1220108 . JSTOR 1220108 .
- Педраса-Пеньалоса, Паола; Салинас, Нельсон Р.; Уилер, Уорд К. (26 апреля 2013 г.). «Схемы жилкования неотропической голубики (Vaccinieae: Ericaceae) и их филогенетическая полезность» (PDF) . Фитотаксы . 96 (1): 1. doi : 10.11646/phytotaxa.96.1.1 . Архивировано (PDF) из оригинала 18 февраля 2017 года . Проверено 17 февраля 2017 г. .
- Рид, Дж.; Стоукс, А. (1 октября 2006 г.). «Биомеханика растений в экологическом контексте» . Американский журнал ботаники . 93 (10): 1546–1565. дои : 10.3732/ajb.93.10.1546 . ПМИД 21642101 .
- Роллан-Лаган, Анн-Гаэль; Амин, Мира; Пакульска, Малгосия (январь 2009 г.). «Количественная оценка структуры жилкования листьев: двумерные карты». Заводской журнал . 57 (1): 195–205. дои : 10.1111/j.1365-313X.2008.03678.x . ПМИД 18785998 .
- Рот-Небельсик, А; Уль, Дитер; Мосбругер, Волкер; Керп, Ганс (май 2001 г.). «Эволюция и функции архитектуры жилкования листьев: обзор» . Анналы ботаники . 87 (5): 553–566. дои : 10.1006/anbo.2001.1391 .
- Рунионс, Адам; Фюрер, Мартин; Лейн, Брендан; Федерл, Павол; Роллан-Лаган, Анн-Гаэль; Прусинкевич, Пшемыслав (1 января 2005 г.). «Моделирование и визуализация узоров жилкования листьев». Доклады ACM SIGGRAPH 2005 . Том. 24. С. 702–711. CiteSeerX 10.1.1.102.1926 . дои : 10.1145/1186822.1073251 . ISBN 9781450378253 . S2CID 2629700 .
- Рутисхаузер, Р.; Саттлер, Р. (1997). «Экспрессия побеговых процессов в развитии листьев Polemonium caeruleum ». Ботанические ежегодники по систематике . 119 :563-582.
- Сак, Лоурен; Скоффони, Кристина (июнь 2013 г.). «Жилкование листьев: строение, функции, развитие, эволюция, экология и применение в прошлом, настоящем и будущем» . Новый фитолог . 198 (4): 983–1000. дои : 10.1111/nph.12253 . ПМИД 23600478 .
- Шелли, Эй Джей; Смит, ВК; Фогельманн, TC (1998). «Онтогенетические различия в структуре мезофилла и распределении хлорофилла у Eucalyptus globulus ssp. globulus (Myrtaceae)» . Американский журнал ботаники . 86 (2): 198–207. дои : 10.2307/2656937 . JSTOR 2656937 . ПМИД 21680359 .
- Цукая, Хирокадзу (январь 2013 г.). «Развитие листьев» . Книга «Арабидопсис» . 11 : e0163. дои : 10.1199/tab.0163 . ПМЦ 3711357 . ПМИД 23864837 .
- Уэно, Осаму; Кавано, Юкико; Вакаяма, Масатака; Такеда, Томосиро (1 апреля 2006 г.). «Сосудистые системы листьев у трав C 3 и C 4 : двумерный анализ» . Анналы ботаники . 97 (4): 611–621. дои : 10.1093/aob/mcl010 . ПМЦ 2803656 . ПМИД 16464879 .
- Уоллс, РЛ (25 января 2011 г.). «Модели жилок листьев покрытосеменных связаны с функциями листьев в наборе данных глобального масштаба». Американский журнал ботаники . 98 (2): 244–253. дои : 10.3732/ajb.1000154 . ПМИД 21613113 .
Веб-сайты
[ редактировать ]- Букш, Александр; Блондер, Бенджамин; Прайс, Чарльз; Винг, Скотт; Вайц, Джошуа; Дас, Абхирам (2017). «Очищенная база данных изображений листьев» . Школа биологии Технологического института Джорджии . Архивировано из оригинала 25 сентября 2014 года . Проверено 12 марта 2017 г.
- Женева, Роберт. «Лист» (PDF) . PLS 220: Введение в идентификацию растений . Университет Кентукки: факультет садоводства. Архивировано из оригинала (PDF) 15 марта 2016 г.
- Клинг, Гэри Дж.; Хайден, Лаура Л.; Поттс, Джошуа Дж. (2005). «Ботаническая терминология» . Университет Иллинойса , Урбана-Шампейн. Архивировано из оригинала 8 марта 2017 года . Проверено 7 марта 2017 г.
- де Кок, Рожье; Биффин, Эд (ноябрь 2007 г.). «The Pea Key: интерактивный ключ к австралийским бобовым с горошком» . Австралийская исследовательская группа по бобовым с цветками гороха. Архивировано из оригинала 26 февраля 2017 года . Проверено 9 марта 2017 г.
- Кранц, Лаура. «Женковые узоры листьев» (рисунки) . Архивировано из оригинала 6 марта 2017 г. Проверено 05 марта 2017 г.
- Мэсси, Джимми Р.; Мерфи, Джеймс К. (1996). «Систематика сосудистых растений» . НК-ботнет . Университет Северной Каролины в Чапел-Хилл. Архивировано из оригинала 17 января 2016 года . Проверено 19 января 2016 г.
- "Листья" . Архивировано из оригинала 25 июля 2016 г. Проверено 19 января 2016 г. , в Мэсси и Мерфи (1996)
- Перселл, Адам (16 января 2016 г.). "Листья" . Базовая биология . Адам Перселл. Архивировано из оригинала 19 апреля 2015 года . Проверено 17 февраля 2017 г. .
- Симпсон, Майкл Г. «Растения округа Сан-Диего, Калифорния» . Колледж наук Государственного университета Сан-Диего . Архивировано из оригинала 3 марта 2017 года . Проверено 2 марта 2017 г.
- «Кнопка листьев ископаемых пластов Флориссана» (PDF) . Национальный памятник «Ископаемые пласты Флориссана» . национальных парков Служба Министерства внутренних дел США . Архивировано (PDF) из оригинала 16 февраля 2017 года . Проверено 16 февраля 2017 г.
- «База данных растений» . Факультет садоводства Квантленского политехнического университета . 2015. Архивировано из оригинала 21 сентября 2017 года . Проверено 9 марта 2017 г.
- «Морфология покрытосеменных» . TutorVista. 2017. Архивировано из оригинала 21 июня 2020 г. Проверено 9 марта 2017 г.
- Глоссарии
- Хьюз, Колин. «Виртуальный полевой гербарий» . Гербарии Оксфордского университета . Архивировано из оригинала 5 марта 2017 года . Проверено 4 марта 2017 г.
- «Характеристика растений» . Архивировано из оригинала (Глоссарий) 5 марта 2017 года . Проверено 4 марта 2017 г. , в Хьюзе (2017)
- «Словарь ботанических терминов» . Неотропики . Королевский ботанический сад, Кью. Архивировано из оригинала 21 января 2017 года . Проверено 18 февраля 2017 г.
- «Иллюстрированный словарь форм листьев» (PDF) . Центр водных и инвазивных растений, Институт пищевых и сельскохозяйственных наук , Университет Флориды. 2009. Архивировано (PDF) из оригинала 10 января 2020 года . Проверено 8 января 2020 г.
- «Листья» . Донсгарден . Архивировано из оригинала 4 февраля 2016 года . Проверено 9 января 2020 г.
Внешние ссылки
[ редактировать ]- Рендл, Альфред Бартон (1911). . Британская энциклопедия . Том. 15 (11-е изд.). п. 322-329.
- Ингерсолл, Эрнест (1920). . Американская энциклопедия . Том. XVII.