Эволюционная история растений

Эволюция морские растений пресноводные привела к широкому спектру сложностей: от самых ранних водорослевых матов одноклеточных архепластид, развившихся в результате эндосимбиоза , через многоклеточные и и зеленые водоросли до спороносных наземных мохообразных , плаунов и папоротников , в конечном итоге, до сложных семян. -плодоносящие голосеменные и покрытосеменные ( цветковые растения) наших дней. Хотя многие из самых ранних групп продолжают процветать, о чем свидетельствуют красные и зеленые водоросли в морской среде, возникшие позднее группы вытеснили ранее экологически доминирующие; например, превосходство цветковых растений над голосеменными в наземной среде. [1] : 498
Есть свидетельства того, что цианобактерии и многоклеточные слоевищные эукариоты жили в пресноводных сообществах на суше еще 1 миллиард лет назад. [2] и что сообщества сложных, многоклеточных фотосинтезирующих организмов существовали на суше в позднем докембрии , около 850 миллионов лет назад . [3]
Свидетельства появления наземных растений -эмбриофитов впервые встречаются в среднем ордовике (~ 470 миллионов лет назад ), а к середине девона (~ 390 миллионов лет назад ) уже присутствовали многие черты, распознаваемые сегодня у наземных растений, в том числе корни и листья. К концу девона (~ 370 миллионов лет назад ) некоторые растения со свободными спорами, такие как Archaeopteris, имели вторичную сосудистую ткань , которая производила древесину и образовывала леса из высоких деревьев. Также к концу девона у Элкинсии , раннего семенного папоротника , появились семена. [4] Эволюционные инновации продолжались на протяжении всего фанерозоя и продолжаются до сих пор. Большинство групп растений относительно не пострадало от пермо-триасового вымирания , хотя структура сообществ изменилась. Возможно, это подготовило почву для появления цветковых растений в триасе (~ 200 миллионов лет назад ) и их более поздней диверсификации в меловом и палеогеновом периодах . Последней крупной группой растений, появившейся в процессе эволюции, были травы , которые приобрели важное значение в середине палеогена, примерно 40 миллионов лет назад . Травы, как и многие другие группы, развили новые механизмы метаболизма, чтобы выжить в условиях низкого содержания CO 2 и теплых и засушливых условиях тропиков за последние 10 миллионов лет .
Колонизация земель
[ редактировать ]Дивергенция
[ редактировать ]Наземные растения произошли от группы пресноводных зеленых водорослей , возможно, уже в 850 млн лет назад. [3] но водорослеподобные растения могли появиться еще 1 миллиард лет назад. [2] Ближайшими ныне живущими родственниками наземных растений являются харофиты , особенно Charales ; если современные Charales похожи на своих далеких предков, которых они разделяют с наземными растениями, это означает, что наземные растения произошли от разветвленной нитчатой водоросли, обитающей на мелководье в пресной воде, [5] возможно, на краю сезонно высыхающих водоемов. [6] Однако некоторые недавние данные свидетельствуют о том, что наземные растения могли произойти от одноклеточных наземных харофитов, сходных с современными Klebsormidiophyceae . [7] У водоросли был гаплонтический жизненный цикл . У него были парные хромосомы ( диплоидное состояние) лишь на очень короткое время, когда яйцеклетка и сперматозоид впервые слились с образованием зиготы , которая сразу же разделилась бы путем мейоза с образованием клеток с половиной количества непарных хромосом ( гаплоидное состояние). Совместное взаимодействие с грибами , возможно, помогло ранним растениям адаптироваться к стрессам наземного мира. [8]

Проблемы колонизации земель
[ редактировать ]Растения не были первыми фотосинтезаторами на суше. Скорость выветривания позволяет предположить, что организмы, способные к фотосинтезу, уже жили на земле 1200 миллионов лет назад . [6] а окаменелости микробов были обнаружены в отложениях пресноводных озер 1000 миллионов лет назад . [9] но данные по изотопам углерода позволяют предположить, что их было слишком мало, чтобы влиять на состав атмосферы примерно 850 миллионов лет назад . [3] Эти организмы, хотя и филогенетически разнообразны, [10] вероятно, были маленькими и простыми, образуя не более чем водорослевую пену. [6]
Поскольку лишайники инициируют первый шаг первичной экологической сукцессии в современных условиях, одна из гипотез заключалась в том, что лишайники первыми вышли на сушу и способствовали колонизации растениями; однако и молекулярная филогения, и летопись окаменелостей, похоже, противоречат этому. [11]
Существует множество потенциальных причин, по которым появление наземных растений заняло так много времени. Возможно, атмосферное «отравление» помешало эукариотам колонизировать землю до появления наземных растений. [12] или, возможно, просто потребовалось много времени, чтобы возникла необходимая сложность. [13] Серьезной проблемой адаптации земель было бы отсутствие подходящей почвы . [14] На протяжении всей летописи окаменелостей почва сохраняется, что дает информацию о том, какими были ранние почвы. До появления наземных растений почва на суше была бедна необходимыми для жизни ресурсами, такими как азот и фосфор , и имела небольшую способность удерживать воду.
Адаптации к колонизации земель
[ редактировать ]Свидетельства существования самых ранних наземных растений встречаются примерно 470 миллионов лет назад в породах нижнего и среднего ордовика Саудовской Аравии. [15] и Гондвана [16] в форме спор, известных как криптоспоры . Эти споры имеют стенки из спорополленина , чрезвычайно устойчивого к гниению материала, что означает, что они хорошо сохранились в летописи окаменелостей.Эти споры образовывались поодиночке (монады), парами (диады) или группами по четыре (тетрады), и их микроструктура напоминает структуру современных спор печеночника , что позволяет предположить, что они имеют одинаковую степень организации. [17] Их стенки содержат спорополленин – еще одно свидетельство родства с эмбриофитами. [18]
Трилетные споры, похожие на споры сосудистых растений , появляются вскоре после этого в породах верхнего ордовика около 455 миллионов лет назад. [19] [20] В зависимости от того, когда тетрада разделяется, каждая из четырех спор может иметь «тройную метку», Y-образную форму, отражающую точки, в которых каждая клетка сталкивалась со своими соседями. [21] Однако для этого необходимо, чтобы стенки спор на ранней стадии были прочными и устойчивыми. Эта устойчивость тесно связана с наличием устойчивой к высыханию внешней стенки — черта, которую можно использовать только тогда, когда споры должны выжить вне воды. Действительно, даже те эмбриофиты , которые вернулись в воду, не имеют устойчивой стенки и поэтому не несут трилетных меток. [21] Тщательное изучение спор водорослей показывает, что ни одна из них не имеет трилетных спор либо потому, что их стенки недостаточно устойчивы, либо, в тех редких случаях, когда они есть, потому, что споры рассеиваются до того, как сжимаются настолько, чтобы образовать след, или не помещаются в тетраэдрическая тетрада. [21]
Самыми ранними мегаокаменелостями наземных растений были слоевищные организмы, которые обитали в речных водно-болотных угодьях и, как обнаружено, покрывали большую часть раннего силура поймы . Они могли выжить только тогда, когда земля была затоплена. [22] Были еще микробные коврики. [23]
Когда растения достигли земли, появилось два подхода к борьбе с высыханием. Современные мохообразные либо избегают этого, либо поддаются ему, ограничивая свой ареал влажными условиями или высыхая и «приостанавливая» свой метаболизм до тех пор, пока не поступит больше воды, как у печеночников из рода Targionia . Трахеофиты сопротивляются высыханию, контролируя скорость потери воды. Все они имеют водонепроницаемый внешний слой кутикулы везде, где они подвергаются воздействию воздуха (как и некоторые мохообразные), чтобы уменьшить потерю воды, но поскольку полное покрытие отрезало бы их от CO 2 в атмосфере, трахеофиты используют переменные отверстия, устьица , для регулируют скорость газообмена. У трахеофитов также развилась сосудистая ткань, способствующая передвижению воды внутри организмов (см. Ниже ), и они отошли от жизненного цикла, в котором доминируют гаметофиты (см. Ниже ). Сосудистая ткань в конечном итоге также способствовала вертикальному росту без поддержки воды и проложила путь к эволюции более крупных растений на суше. [ нужна ссылка ]
Последствия
[ редактировать ]Считается, что глобальное оледенение под названием «Земля-снежок» , произошедшее примерно в 720–635 млн лет назад в криогенный период, было, по крайней мере частично, вызвано ранними фотосинтезирующими организмами, которые снизили концентрацию углекислого газа и уменьшили парниковый эффект в атмосфере. [24] что приводит к ледниковому климату . Основываясь на исследованиях молекулярных часов , проведенных примерно за предыдущее десятилетие, исследование 2022 года показало, что предполагаемое время происхождения многоклеточных стрептофитов (всех, кроме одноклеточной базальной клады Mesostigmatophyceae ) приходилось на холодный криогенный период, в то время как время последующего разделения стрептофитов упало. в теплом Эдиакаре , что они интерпретировали как указание на избирательное давление ледникового периода на фотосинтезирующие организмы, группе которых удалось выжить в относительно более теплых эдафических рефугиумах, впоследствии процветая в позднем эдиакаре и фанерозое на суше в качестве эмбриофитов. Исследование также предположило, что одноклеточная морфология и другие уникальные особенности Zygnematophyceae могут отражать дальнейшую адаптацию к криофильной жизни. [25] Создание наземной флоры увеличило скорость накопления кислорода в атмосфере, поскольку наземные растения производили кислород в качестве отходов. Когда эта концентрация превысила 13%, около 0,45 миллиарда лет назад, [26] стали возможны лесные пожары, о чем свидетельствует древесный уголь в летописи окаменелостей. [27] Помимо спорного пробела в позднем девоне, с тех пор здесь присутствует древесный уголь.
Уголизация - важный тафономический способ. Лесной пожар или захоронение в горячем вулканическом пепле удаляют летучие соединения, оставляя только остаток чистого углерода. Это неподходящий источник пищи для грибов, травоядных и детритофагов, поэтому его легко сохранить. Он также прочен и может выдерживать давление, демонстрируя изысканные, иногда субклеточные, детали останков. [ нужна ссылка ]
Эволюция жизненных циклов
[ редактировать ]
Все многоклеточные растения имеют жизненный цикл, состоящий из двух поколений или фаз. Фаза гаметофита . имеет один набор хромосом (обозначается 1n ) и производит гаметы (сперматозоиды и яйцеклетки) Фаза спорофита имеет парные хромосомы (обозначенные 2n ) и производит споры. Фазы гаметофита и спорофита могут быть гомоморфными и выглядеть идентичными у некоторых водорослей, таких как Ulva Lactuca , но сильно различаться у всех современных наземных растений — состояние, известное как гетероморфия . [ нужна ссылка ]
В эволюции растений произошел переход от гомоморфии к гетероморфии. Водорослевые предки наземных растений почти наверняка были гаплобионтными , будучи гаплоидными на протяжении всего жизненного цикла, с одноклеточной зиготой, обеспечивающей стадию 2N. Все наземные растения (т. е. эмбриофиты ) диплобионтны , то есть как гаплоидная, так и диплоидная стадии являются многоклеточными. [1] Очевидны две тенденции: у мохообразных ( печеночников , мхов и роголистников ) гаметофит стал доминирующей фазой жизненного цикла, при этом спорофит становится почти полностью зависимым от него; У сосудистых растений спорофит является доминирующей фазой, при этом количество гаметофитов особенно редуцируется у семенных растений . [ нужна ссылка ]
В качестве основы возникновения диплоидной фазы жизненного цикла как доминирующей фазы было предложено то, что диплоидия позволяет маскировать проявление вредных мутаций посредством генетической комплементации . [28] [29] Таким образом, если один из родительских геномов в диплоидных клетках содержит мутации, приводящие к дефектам одного или нескольких генных продуктов , эти недостатки могут быть компенсированы другим родительским геномом (который, тем не менее, может иметь свои собственные дефекты в других генах). Поскольку диплоидная фаза становилась преобладающей, эффект маскировки, вероятно, позволил увеличить размер генома и, следовательно, информационное содержание без необходимости повышения точности репликации. Возможность увеличить содержание информации при низких затратах является преимуществом, поскольку позволяет кодировать новые адаптации. Эта точка зрения была оспорена: данные показывают, что отбор не более эффективен на гаплоидных, чем на диплоидных фазах жизненного цикла мхов и покрытосеменных. [30]
Есть две конкурирующие теории, объясняющие появление дилобионтического жизненного цикла. [ нужна ссылка ]
Теория интерполяции (также известная как антитетическая или интеркалярная теория) [31] считает, что интерполяция фазы многоклеточного спорофита между двумя последовательными поколениями гаметофита была инновацией, вызванной предшествующим мейозом в только что проросшей зиготе с одним или несколькими раундами митотического деления, в результате чего образуется некоторое количество диплоидной многоклеточной ткани, прежде чем, наконец, мейоз приведет к образованию спор. Эта теория подразумевает, что первые спорофиты имели совершенно другую и более простую морфологию по сравнению с гаметофитом, от которого они зависели. [31] Кажется, это хорошо согласуется с тем, что известно о мохообразных, у которых вегетативный слоевищный гаметофит питает простой спорофит, который состоит не более чем из неразветвленного спорангия на стебле. Возрастающая сложность изначально простого спорофита, включая возможное приобретение фотосинтетических клеток, освободит его от зависимости от гаметофита, как это наблюдается у некоторых роголистников ( Anthoceros ), и в конечном итоге приведет к тому, что спорофит разовьет органы и сосудистую ткань и станет доминантная фаза, как у трахеофитов (сосудистых растений). [1] Эта теория может быть подтверждена наблюдениями о том, что более мелкие особи Cooksonia, должно быть, поддерживались поколением гаметофитов. Наблюдаемое появление более крупных осевых размеров, в которых есть место для фотосинтетической ткани и, следовательно, самоподдержания, обеспечивает возможный путь развития самодостаточной фазы спорофита. [31]
Альтернативная гипотеза, называемая теорией трансформации (или гомологичной теорией), утверждает, что спорофит мог появиться внезапно, отложив возникновение мейоза до тех пор, пока не сформировался полностью развитый многоклеточный спорофит. Поскольку и гаплоидная, и диплоидная фазы будут использовать один и тот же генетический материал, они будут выглядеть одинаково. Это объясняет поведение некоторых водорослей, таких как Ulvalactuca , которые производят чередующиеся фазы идентичных спорофитов и гаметофитов. Последующая адаптация к высыхающей почве, затрудняющей половое размножение, могла привести к упрощению сексуально активного гаметофита и развитию фазы спорофита для лучшего рассеивания водонепроницаемых спор. [1] Ткань спорофитов и гаметофитов сосудистых растений, таких как Rhynia, сохранившаяся в кремнях Рини, имеет аналогичную сложность, что считается подтверждением этой гипотезы. [31] [32] [33] Напротив, современные сосудистые растения, за исключением Psilotum , имеют гетероморфные спорофиты и гаметофиты, в которых гаметофиты редко имеют сосудистую ткань. [34]
Эволюция анатомии растений
[ редактировать ]Арбускулярный микоризный симбиоз
[ редактировать ]Нет никаких доказательств того, что ранние наземные растения силура и раннего девона имели корни, хотя ископаемые свидетельства наличия ризоидов встречаются у нескольких видов, таких как Horneophyton . Самые ранние наземные растения также не имели сосудистой системы для транспорта воды и питательных веществ. Аглаофитон , бескорневое сосудистое растение, известное по девонским окаменелостям в кремнях Рини. [35] было первым наземным растением, у которого было обнаружено симбиотическое отношение к грибам. [36] которые образовывали арбускулярные микоризы , буквально «древесные корни грибов», в четко выраженном цилиндре клеток (кольцо на поперечном сечении) в коре его стеблей. Грибы питались сахарами растений в обмен на питательные вещества, вырабатываемые или извлекаемые из почвы (особенно фосфаты ), к которым в противном случае растение не имело бы доступа. Как и другие бескорневые наземные растения силура и раннего девона, аглаофитон, возможно, полагался на арбускулярные микоризные грибы для получения воды и питательных веществ из почвы. [37]
Грибы принадлежали к типу Glomeromycota . [38] группа, которая, вероятно, впервые появилась 1 миллиард лет назад и до сих пор образует арбускулярные микоризные ассоциации со всеми основными группами наземных растений от мохообразных до птеридофитов, голосеменных и покрытосеменных растений, а также с более чем 80% сосудистых растений. [39]
Данные анализа последовательности ДНК показывают, что арбускулярный микоризный мутуализм возник у общего предка этих групп наземных растений во время их перехода на сушу. [40] и, возможно, это даже был решающий шаг, который позволил им колонизировать эту землю. [41] Появившись до того, как у этих растений появились корни, микоризные грибы помогали растениям усваивать воду и минеральные питательные вещества, такие как фосфор , в обмен на органические соединения, которые они не могли синтезировать самостоятельно. [39] Такие грибы повышают продуктивность даже таких простых растений, как печеночники. [42] [43]
Кутикула, устьица и межклеточные пространства
[ редактировать ]Для фотосинтеза растения должны поглощать CO2 из атмосферы. Однако обеспечение доступности тканей для поступления CO 2 позволяет воде испаряться, поэтому за это приходится платить. [44] Вода теряется гораздо быстрее, чем . поглощается CO2, поэтому растениям необходимо ее восполнять Ранние наземные растения переносили воду апопластически , внутри пористых стенок своих клеток. Позже они развили три анатомические особенности, которые обеспечили возможность контролировать неизбежную потерю воды, сопровождающую поглощение CO 2 . Во-первых, появилось водонепроницаемое внешнее покрытие или кутикула , которая уменьшила потерю воды. Во-вторых, переменные отверстия, устьица , которые могли открываться и закрываться, чтобы регулировать количество воды, теряемой при испарении во время поглощения CO 2 и, в-третьих, межклеточное пространство между фотосинтезирующими клетками паренхимы, что позволяло улучшить внутреннее распределение CO 2 в хлоропластах. Эта трехкомпонентная система обеспечивала улучшенную гомогидрию, регулирование содержания воды в тканях, обеспечивая особое преимущество, когда подача воды непостоянна. [45] Высокие концентрации CO 2 в силурийском и раннем девоне, когда растения впервые колонизировали сушу, означали, что они использовали воду относительно эффективно. Поскольку CO 2 извлекался из атмосферы растениями, при его улавливании терялось больше воды, и появились более элегантные механизмы сбора и транспортировки воды. [44] Растениям, растущим вверх в воздух, требовалась система транспортировки воды из почвы ко всем различным частям надпочвенного растения, особенно к фотосинтезирующим частям. К концу каменноугольного периода , когда концентрация CO 2 снизилась до уровня, приближающегося к сегодняшнему, на единицу поглощения CO 2 терялось примерно в 17 раз больше воды . [44] Однако даже в «легкие» первые дни вода всегда была в дефиците, и ее приходилось доставлять к частям растения из влажной почвы, чтобы избежать высыхания. [45]
Вода может проникать капиллярным путем по ткани с небольшими пространствами. В узких столбах воды, например, в стенках растительных клеток или в трахеидах, когда молекулы испаряются с одного конца, они тянут молекулы за собой по каналам. Следовательно, только испарение является движущей силой водного транспорта в растениях. [44] Однако без специализированных транспортных сосудов этот механизм сцепления-напряжения может вызывать отрицательное давление, достаточное для разрушения водопроводящих клеток, ограничивая транспортируемую воду до уровня не более нескольких см и, следовательно, ограничивая размер самых ранних растений. [44]

Ксилема
[ редактировать ]Чтобы освободиться от ограничений небольшого размера и постоянной влажности, которые накладывала паренхиматозная транспортная система, растениям требовалась более эффективная система водного транспорта. По мере роста растений вверх развивались специализированные сосудистые ткани , транспортирующие воду , сначала в виде простых гидроидов, подобных тем, которые встречаются в щетинках спорофитов мхов. Эти простые удлиненные клетки в зрелом возрасте были мертвыми и заполненными водой, обеспечивая канал для транспорта воды, но их тонкие, неармированные стенки разрушались при умеренном натяжении воды, ограничивая высоту растения. Ксилемные трахеиды , более широкие клетки с клеточными стенками, усиленными лигнином , которые были более устойчивы к разрушению под действием напряжения, вызванного водным стрессом, встречаются более чем в одной группе растений к середине силура и могут иметь единое эволюционное происхождение, возможно, внутри роголистников. [46] объединяющий все трахеофиты. Альтернативно, они могли эволюционировать более одного раза. [44] Значительно позже, в меловом периоде, за трахеидами последовали сосуды у цветковых растений . [44] По мере развития механизмов переноса воды и водонепроницаемой кутикулы растения смогли выжить, не будучи постоянно покрыты пленкой воды. Этот переход от пойкилогидрии к гомойогидрии открыл новый потенциал для колонизации. [44] [45]
Ранние девонские претрахеофиты Aglaophyton и Horneophyton имеют неармированные водотранспортные трубки со структурой стенок, очень похожими на моховые гидроиды, но они росли рядом с несколькими видами трахеофитов , такими как Rhynia gwynne-vaughanii, у которых были ксилемные трахеиды, хорошо укрепленные полосами лигнина. Самыми ранними макрофоссилиями, имеющими ксилемные трахеиды, являются небольшие среднесилурийские растения рода Cooksonia . [47] Однако начиная с раннего силура заметны утолщенные полосы на стенках отдельных фрагментов трубок. [48]
Растения продолжали изобретать способы снижения сопротивления потоку внутри своих клеток, постепенно увеличивая эффективность транспорта воды и увеличивая сопротивление трахеид схлопыванию под напряжением. [49] [50] возможно, стабилизировался в зостерофиллах . В раннем девоне максимальный диаметр трахеид со временем увеличивался, но к середине девона, [49] Общая скорость транспорта также зависит от общей площади поперечного сечения самого пучка ксилемы, а некоторые растения среднего девона, такие как тримерофиты, имели гораздо большие стелы, чем их ранние предки. [49] Хотя более широкие трахеиды обеспечивали более высокие скорости переноса воды, они увеличивали риск кавитации — образования пузырьков воздуха в результате разрыва водного столба под напряжением. [44] Небольшие ямки в стенках трахеиды позволяют воде обходить дефектную трахеиду, предотвращая при этом прохождение пузырьков воздуха. [44] но за счет ограничения скорости потока. К каменноугольному периоду у голосеменных появились окаймленные ямки . [51] [52] клапаноподобные структуры, которые позволяют ямкам с высокой проводимостью закрываться при разгерметизации одной стороны трахеиды.
Трахеиды имеют неперфорированные торцевые стенки с ямками, которые оказывают большое сопротивление потоку воды. [49] но, возможно, имело преимущество изолировать воздушную эмболию, вызванную кавитацией или замораживанием. Сосуды впервые появились в засушливые периоды с низким содержанием CO 2 в поздней перми независимо у хвощей, папоротников и Selaginellales , а позже появились в середине мела у гнетофитов и покрытосеменных растений. [44] Элементы резервуара представляют собой открытые трубы без торцевых стенок и расположены встык, чтобы работать так, как если бы они были одним непрерывным резервуаром. [49] Суда позволяли древесине той же площади поперечного сечения переносить гораздо больше воды, чем трахеиды. [44] Это позволило растениям заполнить большую часть своих стеблей структурными волокнами , а также открыло новую нишу для виноградных лоз, которые могли переносить воду, не будучи такими толстыми, как дерево, на котором они росли. [44] Несмотря на эти преимущества, древесина на основе трахеид намного легче и, следовательно, дешевле в производстве, поскольку сосуды необходимо гораздо более армировать, чтобы избежать кавитации. [44] Как только растения развили этот уровень контроля над испарением и транспортом воды, они стали по-настоящему гомойогидридными , способными извлекать воду из окружающей среды через корнеподобные органы, а не полагаться на пленку поверхностной влаги, что позволило им вырасти до гораздо больших размеров. [45] [44] но в результате возросшей независимости от окружающей среды большинство сосудистых растений утратили способность переживать высыхание, а потеря этой черты обходится дорого. [44] У ранних наземных растений поддержка в основном обеспечивалась тургорным давлением, особенно внешнего слоя клеток, известного как стеромные трахеиды, а не ксилемой, которая была слишком маленькой, слишком слабой и находилась в слишком центральном положении, чтобы обеспечить значительную структурную поддержку. . [44] Растения со вторичной ксилемой , появившиеся в середине девона, такие как тримерофиты и прогимноспермы, имели гораздо большие поперечные сечения сосудов, образующие прочную древесную ткань. [ нужна ссылка ]
эндодерма
[ редактировать ]Эндодерма , возможно, возникла в самых ранних корнях растений в девонском периоде, но первые ископаемые свидетельства такой структуры относятся к каменноугольному периоду. [44] Эндодерма в корнях окружает водную транспортную ткань и регулирует ионный обмен между грунтовыми водами и тканями, а также предотвращает попадание нежелательных патогенов и т. д. в водную транспортную систему. Энтодерма также может оказывать давление вверх, вытесняя воду из корней, когда транспирация недостаточна.
Эволюция морфологии растений
[ редактировать ]Листья
[ редактировать ]


Листья – основные фотосинтезирующие органы современного растения. Появление листьев почти наверняка было вызвано падением концентрации CO 2 в атмосфере в девонский период, что привело к увеличению эффективности улавливания углекислого газа для фотосинтеза . [53] [54]
Листья развивались более одного раза. По своей структуре их подразделяют на два типа: микрофиллы , лишенные сложного жилкования и, возможно, возникшие в виде колючих выростов, известных как энации, и мегафиллы , которые имеют большие размеры и имеют сложное жилкование , которые могли возникнуть в результате модификации групп ветвей. . Было высказано предположение, что эти структуры возникли независимо. [55] Мегафиллы, согласно теломной теории Уолтера Циммермана, [56] произошли от растений, демонстрировавших трехмерную архитектуру ветвления, посредством трех трансформаций — переворачивания , которое привело к латеральному положению, типичному для листьев, плантации , которая включала образование плоской архитектуры, перепонки или слияния , которые объединяли плоские ветви, таким образом что приводит к формированию правильной листовой пластинки. Все три этапа происходили несколько раз в эволюции сегодняшних листьев. [57]
Широко распространено мнение, что теломная теория хорошо подтверждается ископаемыми свидетельствами. Однако Вольфганг Хагеманн поставил под сомнение это по морфологическим и экологическим причинам и предложил альтернативную теорию. [58] [59] Если согласно теломной теории самые примитивные наземные растения имеют трехмерную ветвящуюся систему радиально-симметричных осей (теломов), то согласно альтернативе Хагемана предполагается обратное: наиболее примитивные наземные растения, давшие начало сосудистым растениям, были плоскими, слоевищными. , листовидная, без осей, чем-то похожая на заросток печеночника или папоротника. Такие оси, как стебли и корни, позже развились как новые органы. Рольф Саттлер предложил всеобъемлющий, ориентированный на процесс взгляд, который оставляет некоторое ограниченное пространство как для теломной теории, так и для альтернативы Хагемана, и, кроме того, принимает во внимание весь континуум между дорзивентральными (плоскими) и радиальными (цилиндрическими) структурами, которые можно обнаружить в ископаемых и живых организмах. наземные растения. [60] [61] Эта точка зрения подтверждается исследованиями в области молекулярной генетики. Таким образом, Джеймс (2009) [62] пришел к выводу, что «в настоящее время широко признано, что... радиальность [характеристика осей, таких как стебли] и дорзивентральность [характеристика листьев] являются лишь крайностями непрерывного спектра. Фактически, это просто время экспрессии гена KNOX» .
До появления листьев у растений был фотосинтетический аппарат на стеблях, который они сохранили, хотя эту работу в основном взяли на себя листья. Сегодняшние листья мегафилла, вероятно, стали обычным явлением примерно 360 млн лет назад, примерно через 40 млн лет после того, как простые безлистные растения колонизировали землю в раннем девоне . Это распространение было связано с падением концентрации углекислого газа в атмосфере в позднепалеозойскую эру , связанное с увеличением плотности устьиц на поверхности листьев. [53] Это привело бы к увеличению скорости транспирации и газообмена, но особенно при высоких концентрациях CO 2 большие листья с меньшим количеством устьиц нагревались бы до смертельных температур под ярким солнечным светом. Увеличение плотности устьиц позволило лучше охлаждать лист, что сделало возможным его распространение, но увеличило поглощение CO 2 за счет снижения эффективности использования воды. [54] [63]
Риниофиты Rhynie chert состояли только из тонких неорнаментированных топоров. раннего и среднего девона Тримерофиты можно считать листоватыми. Эту группу сосудистых растений можно узнать по массе терминальных спорангиев, украшающих концы осей, которые могут раздваиваться или раздваиваться. [1] Некоторые организмы, такие как Psilophyton , имели энации . Это небольшие колючие выросты стебля, лишенные собственного кровоснабжения. [ нужна ссылка ]
Зостерофиллы играли важную роль уже в позднем силуре, намного раньше , чем любые риниофиты сопоставимой сложности. [64] Эту группу можно узнать по почковидным спорангиям, которые росли на коротких боковых ветвях рядом с главными осями, иногда разветвляясь в характерной H-образной форме. [1] Многие зостерофиллы имели на своей оси энации (небольшие тканевые выросты на поверхности с различной морфологией), но ни один из них не имел сосудистого следа. Первые свидетельства васкуляризированных енаций встречаются в ископаемых плаунах, известных как Baragwanathia , которые уже появились в летописи окаменелостей в позднем силуре. [65] В этом организме эти следы листьев продолжаются в листе, образуя среднюю жилку. [66] Одна теория, «теория энации», утверждает, что микрофилловые листья плаунов развиваются в результате выростов протостелы, соединяющейся с существующими энациями. [1] Листья рода Rhynie Asteroxylon , сохранившегося в рини-кремне почти на 20 миллионов лет позже, чем Baragwanathia , имели примитивное кровоснабжение – в виде листовых следов, отходящих от центральной протостелы в сторону каждого отдельного «листа». [67] Asteroxylon и Baragwanathia широко рассматриваются как примитивные плауноногие. [1] группа, существующая до сих пор, представлена иголками , колючими мхами и плаунами . Плауноподы несут характерные микрофиллы , представляющие собой листья с единственным сосудистым следом. Микрофиллы могут достигать определенных размеров, у Lepidodendrales достигают более метра в длину, но почти все несут только один сосудистый пучок. Исключением является редкое ветвление у некоторых видов Selaginella .
Считается, что более знакомые листья, мегафиллы , возникли четыре раза независимо: у папоротников, хвощей, прогимноспермовых и семенных растений. [68] По-видимому, они возникли в результате модификации дихотомических ветвей, которые сначала перекрывали (или «перекрывали») друг друга, становились уплощенными или плоскими и в конечном итоге развили «перепонку» и постепенно превратились в более листообразные структуры. [66] Мегафиллы, согласно Циммермана теломной теории , состоят из группы перепончатых ветвей. [66] и, следовательно, «листовой зазор», оставшийся там, где сосудистый пучок листа выходит из основной ветви, напоминает расщепление двух осей. [66] В каждой из четырех групп, развивших мегафиллы, их листья впервые появились в период от позднего девона до раннего карбона, быстро диверсифицируясь, пока конструкции не утвердились в середине карбона. [68]
Прекращение дальнейшей диверсификации можно объяснить ограничениями развития, [68] поднимая вопрос о том, почему листьям потребовалось так много времени для развития. Растения существовали на суше не менее 50 миллионов лет, прежде чем мегафиллы приобрели большое значение. Однако из раннедевонского рода Eophyllophyton известны мелкие, редкие мезофиллы – поэтому развитие не могло быть препятствием для их появления. [69] Лучшим объяснением на данный момент является то, что содержание CO 2 в атмосфере в это время быстро сокращалось – примерно на 90% в девонском периоде. [70] Это потребовало увеличения плотности устьиц в 100 раз для поддержания скорости фотосинтеза. Когда устьица открываются, позволяя воде испаряться из листьев, это оказывает охлаждающий эффект в результате потери скрытой теплоты испарения. Похоже, что низкая плотность устьиц в раннем девоне означала, что испарение и испарительное охлаждение были ограничены, и что листья перегрелись бы, если бы они выросли до какого-либо размера. Плотность устьиц не могла увеличиться, поскольку примитивные стелы и ограниченная корневая система не могли бы поставлять воду достаточно быстро, чтобы соответствовать скорости транспирации. [54] Очевидно, что листья не всегда полезны, о чем свидетельствует частая вторичная потеря листьев, примером которой являются кактусы и «папоротник-меток» Psilotum .
Вторичная эволюция может скрыть истинное эволюционное происхождение некоторых листьев. Некоторые роды папоротников имеют сложные листья, которые прикрепляются к псевдостеле за счет выроста проводящего пучка, не оставляя промежутков между листьями. [66]
Лиственные деревья имеют еще один недостаток, связанный с наличием листьев. Распространенное мнение о том, что растения сбрасывают листья, когда дни становятся слишком короткими, ошибочно; вечнозеленые растения процветали за Полярным кругом во время последней теплицы на Земле . [71] Общепринятой причиной сбрасывания листьев зимой является попытка справиться с погодой: сила ветра и вес снега гораздо легче переносят без листьев, чтобы увеличить площадь поверхности. Сезонная потеря листьев развивалась независимо несколько раз и проявляется у гинкговых , некоторых пинофитов и некоторых покрытосеменных растений. [72] Потеря листьев также могла возникнуть в ответ на давление насекомых; возможно, было бы дешевле полностью потерять листья зимой или в засушливый сезон, чем продолжать вкладывать ресурсы в их восстановление. [73]
Корни
[ редактировать ]

Эволюция корней имела последствия в глобальном масштабе. Нарушая почву и способствуя ее подкислению (поглощая питательные вещества, такие как нитраты и фосфаты). [74] ), они позволили ему выветриться глубже, вводя соединения углерода глубже в почву. [75] с огромными последствиями для климата. [76] Эти последствия, возможно, были настолько глубокими, что привели к массовому вымиранию . [77]
Хотя в ископаемых почвах позднего силура имеются следы корнеобразных отпечатков, [78] окаменелости тел показывают, что самые ранние растения были лишены корней. У многих были распростертые ветви, раскинувшиеся по земле, с вертикальными осями или слоевищами, разбросанными тут и там, а у некоторых даже были нефотосинтезирующие подземные ветви без устьиц. Корни имеют корневой чехлик , в отличие от специализированных ветвей. [6] Таким образом, хотя силуро-девонские растения, такие как Rhynia и Horneophyton, обладали физиологическим эквивалентом корней, [79] [80] корни, определяемые как органы, отличающиеся от стеблей, появились позже. [6] К сожалению, корни редко сохраняются в летописи окаменелостей. [6]
Ризоиды — небольшие структуры, выполняющие ту же роль, что и корни, обычно размером с клетку, — вероятно, возникли очень рано, возможно, даже до того, как растения колонизировали сушу; они относятся к Characeae , родственной группе водорослей, наземным растениям. [6] Тем не менее, ризоиды, вероятно, эволюционировали более одного раза; лишайников , . ризины Аналогичную роль выполняют, например Даже некоторые животные ( ламеллибрахии ) имеют корнеподобные структуры. [6] Ризоиды отчетливо видны в окаменелостях кремней Рини, они присутствовали в большинстве самых ранних сосудистых растений и на этом основании, по-видимому, предвещали настоящие корни растений. [81]
Более продвинутые структуры распространены в кремне Райни, и многие другие окаменелости сопоставимого раннего девонского возраста имеют структуры, которые выглядят и действуют как корни. [6] Риниофиты имели мелкие ризоиды, а тримерофиты и травянистые плауны кремня имели корневидную структуру, проникающую в почву на несколько сантиметров. [82] Однако ни одна из этих окаменелостей не демонстрирует всех особенностей современных корней. [6] за исключением Asteroxylon , который недавно был признан имеющим корни, которые развились независимо от корней современных сосудистых растений. [83] Корни и корнеподобные структуры становились все более распространенными и глубже проникающими в девонский период , при этом деревья плаунов образовывали корни длиной около 20 см в эйфельском и живетском периоде. На последующем франском этапе к ним присоединились прогимноспермы, укоренившиеся на глубину примерно метра. [82] Настоящие голосеменные и зигоптеридные папоротники также в фамене образовали неглубокую корневую систему. [82]
Ризофоры плаунов обеспечивают несколько иной подход к укоренению. Они были эквивалентны стеблям, а органы, эквивалентные листьям, выполняли роль корешков. [6] Подобная конструкция наблюдается у современных плаунов Isoetes , и это, по-видимому, свидетельствует о том, что корни развивались независимо по крайней мере дважды: у плаунов и других растений. [6] Это предположение подтверждается исследованиями, показывающими, что у ликофитов и эуфиллофитов зарождение корней и их рост способствуют различным механизмам. [84]
Растения с ранними корнями немного более развиты, чем их силурийские предки, и не имеют выделенной корневой системы; однако на плосколежащих осях ясно видно, что они имеют наросты, похожие на ризоиды современных мохообразных. [85]
К среднему и позднему девону у большинства групп растений независимо развилась корневая система той или иной природы. [85] По мере того, как корни становились больше, они могли поддерживать более крупные деревья, а почва выветривалась на большую глубину. [77] Это более глубокое выветривание повлияло не только на вышеупомянутое сокращение CO 2 , но также открыло новые среды обитания для колонизации грибов и животных. [82]
Самые узкие корни современных растений имеют диаметр всего 40 мкм и физически не могли бы переносить воду, даже если бы они были хоть немного уже. [6] Напротив, самые ранние обнаруженные ископаемые корни сузились с 3 мм до менее 700 мкм в диаметре; конечно, тафономия — это окончательный контроль того, какую толщину можно увидеть. [6]
Форма дерева
[ редактировать ]

Ландшафт раннего девона был лишен растительности выше пояса. Большая высота обеспечивала конкурентное преимущество в сборе солнечного света для фотосинтеза, затмении конкурентов и в распространении спор, поскольку споры (а позже и семена) могли быть перенесены на большие расстояния, если они начинали выше. Для достижения больших высот требовалась эффективная сосудистая система. Чтобы достичь древовидности , растения должны были развить древесную ткань, которая обеспечивала бы как поддержку, так и транспорт воды, и, таким образом, им необходимо было развить способность к вторичному росту . Стела растений, подвергающихся вторичному росту, окружена сосудистым камбием , кольцом меристематических клеток, которое производит больше ксилемы внутри и флоэмы снаружи. Поскольку клетки ксилемы состоят из мертвой, одревесневшей ткани, последующие кольца ксилемы добавляются к уже присутствующим, образуя древесину. Окаменелости растений раннего девона показывают, что простая форма древесины впервые появилась по крайней мере 400 миллионов лет назад, в то время, когда все наземные растения были маленькими и травянистыми. [86] Поскольку древесина возникла задолго до появления кустарников и деревьев, вполне вероятно, что ее первоначальное предназначение было водным транспортом, а позже использовалось только для механической поддержки. [87]
Первыми растениями, у которых развился вторичный рост и древесный облик, были, по-видимому, папоротники, а уже в среднем девоне один вид, Wattieza , достигал высоты 8 м и древовидного облика. [88]

Другим кладам не потребовалось много времени, чтобы развить древовидный рост. Позднедевонский Archaeopteris , предшественник голосеменных растений , произошедший от тримерофитов. [89] достигал 30 м в высоту. Прогимноспермы были первыми растениями, у которых образовалась настоящая древесина, выросшая из двустороннего камбия. Первое появление одной из них, Rellimia , пришлось на средний девон. [90] Считается, что настоящая древесина возникла только один раз, что дало начало концепции клады «лигнофитов». [ нужна ссылка ]
Леса Archaeopteris вскоре пополнились древесными плауноногими в форме Lepidodendrales , высота которых превышала 50 м, а ширина у основания - 2 м. Эти древесные плауны стали доминировать в лесах позднего девона и каменноугольного периода, которые дали начало месторождениям угля . [91] Lepidodendrales отличаются от современных деревьев тем, что демонстрируют детерминированный рост: после накопления запаса питательных веществ на более низкой высоте растения «выбегают» единым стволом на генетически определенную высоту, разветвляются на этом уровне, распространяют свои споры и умирают. [92] Они состояли из «дешевой» древесины, что позволяло им быстро расти, причем по крайней мере половина их стеблей содержала полость, заполненную сердцевиной. [1] Их древесина также была образована односторонним сосудистым камбием — он не производил новой флоэмы, а это означает, что стволы не могли со временем расти шире. [ нужна проверка ]
Хвощевые . каламиты появились в каменноугольном периоде В отличие от современного хвоща Equisetum , каламиты имели односторонний сосудистый камбий, что позволяло им развивать древесину и расти на высоту более 10 м, а также многократно ветвиться.
Хотя форма ранних деревьев была похожа на форму сегодняшних, сперматофиты или семенные растения, группа, в которую входят все современные деревья, еще не развились. Доминирующими группами деревьев сегодня являются все семенные растения, голосеменные, к которым относятся хвойные деревья, и покрытосеменные, к которым относятся все плодоносящие и цветущие деревья. не существует деревьев со свободными спорами, таких как Archaeopteris В современной флоре . Долгое время считалось, что покрытосеменные произошли из голосеменных, но недавние молекулярные данные свидетельствуют о том, что их нынешние представители образуют две отдельные группы. [93] [94] [95] Молекулярные данные еще предстоит полностью согласовать с морфологическими данными. [96] [97] [98] но становится общепризнанным, что морфологическое подтверждение парафилии не особенно сильно. [99] Это привело бы к выводу, что обе группы возникли из птеридоспермов, вероятно, еще в перми . [99]
Покрытосеменные растения и их предки играли очень незначительную роль, пока они не диверсифицировались в меловом периоде. Они возникли как маленькие, любящие влагу организмы в подлеске и начали диверсифицироваться начиная с мелового периода. [100] сегодня стать доминирующим представителем небореальных лесов .
Семена
[ редактировать ]

Ранние наземные растения размножались по типу папоротников: споры прорастали в мелкие гаметофиты, которые производили яйца и/или сперму. Эти сперматозоиды плавали по влажной почве, чтобы найти женские органы (архегонии) на том же или другом гаметофите, где они сливались с яйцеклеткой, образуя зародыш, который прорастал в спорофит. [82]
Гетероспоровые растения, как следует из названия, несут споры двух размеров – микроспоры и мегаспоры. Они прорастают с образованием микрогаметофитов и мегагаметофитов соответственно. Эта система проложила путь к семязачаткам и семенам: в крайнем случае мегаспорангии могли нести только одну тетраду мегаспор, а для завершения перехода к настоящим семяпочкам три мегаспоры в исходной тетраде могли быть прерваны, оставив одну мегаспору на каждую. мегаспорангий. [ нужна ссылка ]
Переход к семязачаткам продолжился, когда мегаспора была «заперта» в спорангии, пока она прорастала. Тогда мегагаметофит находился внутри водонепроницаемого покрова, окружавшего семя. Пыльцевое зерно, содержащее микрогаметофит, проросший из микроспоры, использовалось для распространения мужской гаметы, высвобождая склонную к высыханию жгутиковую сперму только тогда, когда она достигала восприимчивого мегагаметофита. [1]
Плауноподы и сфенопсиды прошли долгий путь к семенной привычке, так и не переступив порога. Известны ископаемые мегаспоры плаунов, достигающие 1 см в диаметре и окруженные вегетативной тканью (лепидокарпон, ахламидокарпон); они даже прорастали в мегагаметофит in situ . Однако они не были семязачатками, поскольку нуцеллус, внутренний слой, покрывающий споры, не полностью заключает спору. Остается очень маленькая щель (микропиле), а это означает, что мегаспорангий все еще подвергается воздействию атмосферы. Это имеет два последствия: во-первых, это означает, что он не полностью устойчив к высыханию, а во-вторых, сперматозоидам не нужно «зарываться», чтобы получить доступ к архегониям мегаспоры. [1]
среднедевонский семенных растений из Бельгии , предшественник Был идентифицирован который появился примерно на 20 миллионов лет раньше самых ранних семенных растений . Рункария , маленькая и радиально-симметричная, представляет собой интегументированный мегаспорангий, окруженный купулой. Мегаспорангий многолопастным имеет нераскрытый дистальный отросток, выступающий над покровом . Предполагается, что это расширение участвовало в анемофильном опылении . Рункария проливает новый свет на последовательность приобретения персонажей, ведущую к появлению семени. Рункария обладает всеми качествами семенных растений, за исключением твердой семенной кожуры и системы направления пыльцы к семязачатку. [101]
Первые сперматофиты (буквально: «семенные растения»), то есть первые растения, несущие настоящие семена, называются птеридоспермами : буквально «семенные папоротники», называемые так потому, что их листва состояла из папоротниковидных листьев, хотя они не были тесно связан с папоротниками. Самые старые ископаемые свидетельства существования семенных растений относятся к позднему девонскому периоду и, по-видимому, произошли от более ранней группы, известной как прогимноспермовые растения . Эти ранние семенные растения варьировались от деревьев до небольших вьющихся кустарников; как и большинство ранних прогимноспермовых растений, это были древесные растения с листвой, напоминающей папоротник. Все они имели семяпочки, но не имели шишек, плодов и тому подобного. Хотя трудно проследить раннюю эволюцию семян, происхождение семенных папоротников можно проследить от простых тримерофитов до гомоспоровых аневрофитов . [1] Семенные растения претерпели первую крупную эволюционную радиацию в фаменскую эпоху. [102]
Эта модель семян свойственна практически всем голосеменным растениям (буквально: «голые семена»), большинство из которых заключают свои семена в деревянистую шишку или мясистый прицветник ( например, тис ), но ни одно из них не заключает семена полностью. Покрытосеменные («семена сосудов») — единственная группа, полностью заключающая семя в плодолистик. [ нужна ссылка ]
Полностью закрытые семена открыли для растений новый путь: путь покоя семян . Зародыш, полностью изолированный от внешней атмосферы и, следовательно, защищенный от высыхания, мог пережить несколько лет засухи, прежде чем прорасти.Было обнаружено, что семена голосеменных растений позднего карбона содержат зародыши, что позволяет предположить длительный промежуток между оплодотворением и прорастанием. [103] Этот период связан с вступлением в парниковый период Земли , с соответствующим увеличением засушливости. Это говорит о том, что покой возник как реакция на более засушливые климатические условия, когда стало выгоднее дождаться влажного периода, прежде чем прорастать. [103] Этот эволюционный прорыв, по-видимому, открыл шлюзы: ранее негостеприимные территории, такие как сухие горные склоны, теперь можно было терпеть и вскоре они были покрыты деревьями. [103]
Семена давали своим носителям дополнительные преимущества: они увеличивали вероятность успеха оплодотворенных гаметофитов, а поскольку запас питательных веществ мог быть «упакован» в зародыш, семена могли быстро прорастать в негостеприимной среде, достигая размера, при котором они могли постоять за себя. быстрее. [82] Например, без эндосперма саженцы, растущие в засушливых условиях, не имели бы резервов для того, чтобы вырастить корни достаточно глубоко, чтобы достичь уровня грунтовых вод до того, как они умрут от обезвоживания. [82] Точно так же семенам, прорастающим в мрачном подлеске, требуется дополнительный запас энергии, чтобы быстро вырасти достаточно высоко и получить достаточно света для самообеспечения. [82] Сочетание этих преимуществ дало семенным растениям экологическое преимущество перед ранее доминирующим родом Archaeopteris , тем самым увеличив биоразнообразие ранних лесов. [82]
Несмотря на эти преимущества, оплодотворенные семяпочки часто не созревают в виде семян. [104] Также во время покоя семян (часто связанного с непредсказуемыми и стрессовыми условиями) накапливаются повреждения ДНК. [105] [106] [107] Таким образом, повреждение ДНК является основной проблемой выживания семенных растений, точно так же, как повреждение ДНК является основной проблемой для жизни в целом. [108]
Цветы
[ редактировать ]
Цветы — это видоизмененные листья, которыми обладают только покрытосеменные растения , которые относительно поздно появились в летописи окаменелостей. Группа возникла и разнообразилась в раннем меловом периоде и после этого стала экологически значимой. [109] Цветкоподобные структуры впервые появляются в летописях окаменелостей около 130 млн лет назад в меловом периоде . [110] Однако в 2018 году ученые сообщили об обнаружении ископаемого цветка, появившегося около 180 миллионов лет назад, то есть на 50 миллионов лет раньше, чем предполагалось ранее. [111] Однако эта интерпретация вызывает большие споры. [112]
Красочные и/или острые структуры окружают шишки таких растений, как саговники и гнеталесы , что делает строгое определение термина «цветок» неуловимым. [98]
Основная функция цветка — размножение , которое до эволюции цветка и покрытосеменных растений было делом микроспорофиллов и мегаспорофиллов. Цветок можно считать мощной эволюционной инновацией , поскольку его присутствие позволило растительному миру получить доступ к новым средствам и механизмам размножения. [ нужна ссылка ]
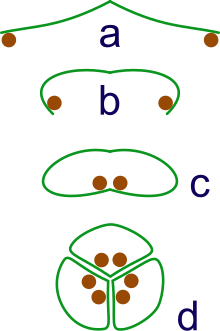
а: спорангии на кончиках листьев
б: Лист скручивается, чтобы защитить спорангии.
в: листья скручиваются, образуя закрытый рулон
г: группировка трех рулонов в синкарп
Долгое время считалось, что цветковые растения произошли из голосеменных ; согласно традиционному морфологическому взгляду, они тесно связаны с Gnetales . Однако, как отмечалось выше, недавние молекулярные данные противоречат этой гипотезе. [94] [95] и далее предполагает, что Gnetales более тесно связаны с некоторыми группами голосеменных, чем с покрытосеменными, [93] и что современные голосеменные образуют отдельную кладу покрытосеменных, [93] [94] [95] эти две клады разошлись около 300 миллионов лет назад . [113]
Связь групп стеблей с покрытосеменными важна для определения эволюции цветов. Стволовые группы дают представление о состоянии предыдущих «развилок» на пути к текущему состоянию. Конвергенция увеличивает риск неправильного определения стволовых групп. Поскольку защита мегагаметофита эволюционно желательна, вероятно, многие отдельные группы независимо развили защитные оболочки. У цветов эта защита принимает форму плодолистика , который развился из листа и выполняет защитную роль, защищая семязачатки. Эти семяпочки дополнительно защищены двустенным покровом .
Для проникновения через эти защитные слои требуется нечто большее, чем просто свободно плавающий микрогаметофит . У покрытосеменных пыльцевые зерна состоят всего из трех клеток. Одна клетка отвечает за просверливание покровов и создание канала для стекания вниз двух сперматозоидов. Мегагаметофит состоит всего из семи клеток; из них один сливается со сперматозоидом, образуя ядро самой яйцеклетки, а другой соединяется с другим сперматозоидом и участвует в формировании богатого питательными веществами эндосперма . Остальные клетки играют вспомогательную роль. [ нужны разъяснения ] Этот процесс « двойного оплодотворения » уникален и присущ всем покрытосеменным.

В летописи окаменелостей есть три интригующие группы, которые имели структуры, похожие на цветы. Первый — пермский птеридосперм Glossopteris , который уже имел загнутые назад листья, напоминающие плодолистики. Мезозойская все еще больше похожа на цветок, с закрытыми семязачатками, но только Caytonia с одним покровом. Кроме того, детали пыльцы и тычинок отличают их от настоящих цветковых растений.
были У Bennettitales удивительно похожие на цветы органы, защищенные мутовками прицветников , которые, возможно, играли ту же роль, что и лепестки и чашелистики настоящих цветов; однако эти цветкоподобные структуры развились независимо, поскольку Bennettitales более тесно связаны с саговниками и гинкго, чем с покрытосеменными. [114]
Однако ни в одной группе, кроме тех, что сохранились до наших дней, не обнаружено настоящих цветов. Большинство морфологических и молекулярных анализов помещают Amborella , нимфеи и Austrobaileyaceae в базальную кладу, называемую «ANA». Эта клада, по-видимому, разделилась в раннем меловом периоде, около 130 миллионов лет назад – примерно в то же время, что и самые ранние ископаемые покрытосеменные растения . [115] [116] и сразу после появления первой пыльцы, похожей на покрытосеменные , 136 миллионов лет назад. [99] разошлись Вскоре после этого магнолииды в результате быстрой радиации появились эвдикоты и однодольные растения , и 125 миллионов лет назад . [99] К концу мелового периода, 66 миллионов лет назад , развилось более 50% современных отрядов покрытосеменных, и на эту кладу приходилось 70% мировых видов. [117] Примерно в это же время цветущие деревья стали доминировать над хвойными . [1] : 498
Особенности базальных групп «АНА» позволяют предположить, что покрытосеменные произошли из темных, влажных, часто нарушенных территорий. [118] Похоже, что покрытосеменные оставались ограниченными такими средами обитания на протяжении всего мелового периода, занимая нишу мелких трав в начале сукцессионного ряда. [117] Возможно, это ограничило их первоначальное значение, но придало им гибкость, которая обусловила быстроту их более поздней диверсификации в других средах обитания. [118]
| Традиционный вид | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| Современный вид | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Некоторые предполагают, что покрытосеменные произошли от неизвестного семенного папоротника Pteridophyte, и рассматривают саговники как живые семенные папоротники как с семенными, так и со стерильными листьями (Cycas revoluta). [97]
В августе 2017 года ученые представили подробное описание и 3D-реконструкцию, возможно, первого цветка , жившего около 140 миллионов лет назад. [119] [120]
Происхождение цветка
[ редактировать ]Семейство Amborellaceae считается родственным всем другим живым цветковым растениям. Проект генома Amborella trichopoda был опубликован в декабре 2013 года. [121] Сравнивая его геном с геномом всех других живых цветковых растений, можно будет определить наиболее вероятные характеристики предка A. trichopoda и всех других цветковых растений, то есть предкового цветкового растения. [122]
Похоже, что на уровне органа лист может быть предком цветка или, по крайней мере, некоторых цветочных органов. Когда некоторые важные гены, участвующие в развитии цветков , мутируют , на месте цветков возникают группы листообразных структур. Таким образом, когда-то в истории программа развития, ведущая к образованию листа, должна была быть изменена, чтобы образовался цветок. Вероятно, также существует общая надежная основа, в рамках которой создается цветочное разнообразие. Примером этого является ген LEAFY (LFY) , который участвует в развитии цветка Arabidopsis thaliana . Гомологи , этого гена обнаружены у покрытосеменных растений таких разнообразных как томат , львиный зев , горох , кукуруза и даже голосеменные растения . Экспрессия Arabidopsis thaliana LFY в отдаленных растениях, таких как тополь и цитрусовые, также приводит к образованию цветов на этих растениях. Ген LFY регулирует экспрессию некоторых генов, принадлежащих к семейству MADS-боксов . Эти гены, в свою очередь, действуют как непосредственные регуляторы развития цветов. [ нужна ссылка ]
Эволюция семейства MADS-box
[ редактировать ]Члены семейства транскрипционных факторов MADS-box играют очень важную и эволюционно консервативную роль в развитии цветка. Согласно модели развития цветка ABC , три зоны — A, B и C — генерируются внутри развивающегося цветочного зачатка под действием некоторых транскрипционных факторов , которые являются членами семейства MADS-box . Среди них функции генов доменов B и C эволюционно более консервативны, чем гены домена A. Многие из этих генов возникли в результате дупликации генов предковых членов этого семейства. Многие из них демонстрируют избыточные функции. [ нужна ссылка ]
Эволюция семейства MADS-box тщательно изучалась. Эти гены присутствуют даже у птеридофитов , но распространение и разнообразие во много раз выше у покрытосеменных . [123] Кажется, существует определенная закономерность в том, как развивалась эта семья. Рассмотрим эволюцию гена C-области AGAMOUS (AG) . У современных цветов это выражено в тычинках и плодолистике , которые являются репродуктивными органами. Его предок голосеменных растений также имеет такой же образец экспрессии. Здесь он выражается в стробилах , органе, производящем пыльцу или семязачатки. [124] Аналогичным образом, предки B-генов (AP3 и PI) экспрессируются только в мужских органах голосеменных растений . Их потомки у современных покрытосеменных также выражены только в тычинках — мужском репродуктивном органе. Таким образом, те же самые, существовавшие тогда компоненты были использованы растениями по-новому для создания первого цветка. Это повторяющаяся закономерность в эволюции . [ нужна ссылка ]
Факторы, влияющие на цветочное разнообразие
[ редактировать ]В структуре цветков растений существуют огромные различия, обычно из-за изменений в генах MADS-бокса и характере их экспрессии. Например, травы обладают уникальной цветочной структурой. Плодолитики и тычинки окружены чешуйчатыми дольками и двумя прицветниками, цветковой леммой и палеей, но генетические данные и морфология позволяют предположить, что дольки гомологичны лепесткам эвдикота . [125] Палея и чешуйка могут быть гомологичны чашелистикам других групп или представлять собой уникальные травяные структуры. [ нужна ссылка ] Другой пример — Linaria vulgaris , у которой есть два типа симметрии цветков — радиальная и двусторонняя . Эта симметрия обусловлена эпигенетическими изменениями только в одном гене, называемом CYCLOIDEA . [110]

Arabidopsis thaliana имеет ген AGAMOUS , который играет важную роль в определении количества лепестков , чашелистиков и других органов. Мутации в этом гене приводят к тому, что цветочная меристема приобретает неопределенную судьбу, а также к пролиферации цветочных органов у махровых форм роз , гвоздик и ипомеи . Эти фенотипы были отобраны садоводами из-за увеличенного количества лепестков . [126] Несколько исследований различных растений, таких как петуния , помидоры , недотроги , кукуруза и т. д., показали, что огромное разнообразие цветов является результатом небольших изменений в генах, контролирующих их развитие. [127]
Проект «Цветочный геном» подтвердил, что модель развития цветков ABC не сохраняется для всех покрытосеменных растений . Иногда домены экспрессии меняются, как в случае многих однодольных , а также у некоторых базальных покрытосеменных, таких как Amborella . Различные модели развития цветков, такие как модель границ затухания или модель перекрывающихся границ , которые предлагают нежесткие области выражения, могут объяснить эту архитектуру. [128] Существует вероятность того, что области цветочной архитектуры, от базальных до современных покрытосеменных, в ходе эволюции становились все более и более фиксированными. [ нужна ссылка ]
Время цветения
[ редактировать ]Еще одна цветочная особенность, ставшая предметом естественного отбора, — это время цветения. Некоторые растения цветут в начале своего жизненного цикла, другим перед цветением требуется период яровизации . Этот результат основан на таких факторах, как температура , интенсивность света , присутствие опылителей и других сигналов окружающей среды: такие гены, как CONSTANS (CO) , цветочный локус C ( FLC ) и FRIGIDA, регулируют интеграцию сигналов окружающей среды в путь развития цветка. Вариации в этих локусах были связаны с различиями во времени цветения разных растений. Например, экотипы Arabidopsis thaliana , произрастающие в холодных регионах с умеренным климатом, требуют длительной яровизации перед цветением, в то время как тропические сорта и наиболее распространенные лабораторные штаммы этого не делают. Это изменение связано с мутациями в генах FLC и FRIGIDA , что делает их нефункциональными. [129]
Многие гены, участвующие в этом процессе, консервативны во всех изученных растениях. Однако иногда, несмотря на генетическую консервативность, механизм действия оказывается иным. Например, рис – растение короткого дня, а Arabidopsis thaliana – растение длинного дня. Оба растения имеют белки CO и ЦВЕТУЩЕЕ ЛОКУСТ T (FT) , но у Arabidopsis thaliana CO усиливает выработку FT , тогда как у риса гомолог CO подавляет выработку FT , что приводит к совершенно противоположным последующим эффектам. [130]
Теории эволюции цветов
[ редактировать ]Теория антофитов была основана на наблюдении, что голосеменные группы Gnetales похожий на цветок имеют семязачаток, . У него частично развиты сосуды , как у покрытосеменных , а мегаспорангий покрыт тремя оболочками, как и структура завязи цветков покрытосеменных. Однако многие другие данные показывают, что Gnetales не имеет отношения к покрытосеменным. [114]
Теория «преимущественно мужчины» имеет более генетическую основу. Сторонники этой теории отмечают, что у голосеменных есть две очень похожие копии гена LFY , а у покрытосеменных только одна. Анализ молекулярных часов показал, что другой паралог LFY был потерян у покрытосеменных растений примерно в то же время, когда окаменелости цветов стали многочисленными, что позволяет предположить, что это событие могло привести к эволюции цветов. [131] Согласно этой теории, потеря одного из LFY паралогов привела к тому, что цветки стали более мужскими, а семязачатки проявились эктопически. Эти семязачатки первоначально выполняли функцию привлечения опылителей , но некоторое время спустя, возможно, были интегрированы в сердцевину цветка. [ нужна ссылка ]
Механизмы и участники эволюции морфологии растений
[ редактировать ]
Хотя факторы окружающей среды в значительной степени ответственны за эволюционные изменения, они действуют лишь как агенты естественного отбора . Изменения по своей сути вызываются явлениями на генетическом уровне: мутациями , хромосомными перестройками и эпигенетическими изменениями. Хотя общие типы мутаций справедливы для всего живого мира, у растений некоторые другие механизмы имеют весьма важное значение. [ нужна ссылка ]
Удвоение генома является относительно распространенным явлением в эволюции растений и приводит к полиплоидии , которая, следовательно, является общей чертой растений. Подсчитано, что по крайней мере половина (а, возможно, и все) растений за свою историю наблюдали удвоение генома. Удвоение генома влечет за собой дупликацию генов , создавая тем самым функциональную избыточность в большинстве генов. Дуплицированные гены могут приобретать новую функцию либо за счет изменений в характере экспрессии, либо за счет изменений активности. Считается, что полиплоидия и дупликация генов являются одними из самых мощных сил в эволюции растительных форм; хотя неизвестно, почему удвоение генома является таким частым процессом у растений. Одной из вероятных причин является производство большого количества вторичных метаболитов в растительных клетках. Некоторые из них могут вмешиваться в нормальный процесс хромосомной сегрегации, вызывая дупликацию генома. [ нужна ссылка ]

В последнее время было показано, что растения обладают значительными семействами микроРНК , которые консервативны во многих линиях растений. По сравнению с животными , хотя количество семейств микроРНК растений меньше, чем у животных, размер каждого семейства намного больше. Гены микроРНК также гораздо более распространены в геноме, чем у животных, где они более кластеризованы. Было высказано предположение, что эти семейства микроРНК расширились за счет дупликации хромосомных областей. [132] многие гены микроРНК, участвующие в регуляции развития растений, вполне консервативны между изученными растениями. Было обнаружено, что [ нужна ссылка ]
Одомашнивание таких растений, как кукуруза , рис , ячмень , пшеница и т. д., также стало важной движущей силой их эволюции. Исследования происхождения кукурузы показали, что это одомашненная производная дикого растения из Мексики, называемого теосинте . Теосинте принадлежит к роду Zea , как и кукуруза, но имеет очень маленькое соцветие , 5–10 твердых початков и сильно разветвленный и распростертый стебель.

Скрещивание определенного сорта теосинте и кукурузы дает плодовитое потомство, занимающее по фенотипу промежуточное положение между кукурузой и теосинте. QTL- анализ также выявил некоторые локусы, которые при мутации в кукурузе дают теосинтеподобный стебель или теосинтеподобные початки. Анализ молекулярных часов этих генов оценивает их происхождение примерно в 9000 лет назад, что вполне соответствует другим данным о приручении кукурузы. Считается, что около 9000 лет назад в Мексике небольшая группа фермеров, должно быть, отобрала некий кукурузоподобный природный мутант теозинте и подвергла его непрерывному отбору, чтобы получить знакомое сегодня растение кукурузы. [133]
Съедобная цветная капуста — это одомашненная версия дикого растения Brassica oleracea , которое не обладает плотными недифференцированными соцветиями , называемыми творогом, которыми обладает цветная капуста.Цветная капуста обладает единственной мутацией в гене под названием CAL , контролирующем дифференцировку меристемы в соцветие . Это приводит к тому, что клетки цветочной меристемы приобретают недифференцированную идентичность и вместо того, чтобы превращаться в цветок , они превращаются в плотную массу клеток меристемы соцветия, находящихся в задержке развития. [134] Эта мутация была выбрана путем одомашнивания, по крайней мере, со времен Греческой империи.
Эволюция путей фотосинтеза
[ редактировать ]
C представляет собой ценную недавнюю эволюционную инновацию в растениях, включающую 4 Метаболический путь сложный набор адаптивных изменений в физиологии и моделях экспрессии генов . [135]
Фотосинтез — это сложный химический путь, которому способствует ряд ферментов и коферментов. Фермент – то есть он присоединяет его к молекуле RuBisCO отвечает за «фиксацию» CO 2 на основе углерода с образованием сахара, который может быть использован растением, выделяя молекулу кислорода. Однако этот фермент заведомо неэффективен и по мере повышения температуры окружающей среды все чаще фиксирует кислород вместо CO 2 в процессе, называемом фотодыхание . Это энергетически затратно, поскольку растению приходится использовать энергию, чтобы вернуть продукты фотодыхания в форму, способную реагировать с CO 2 . [ нужна ссылка ]
Концентрация углерода
[ редактировать ]Растения C 4 развили механизмы концентрации углерода, которые работают за счет увеличения концентрации CO 2 вокруг RuBisCO и исключения кислорода, тем самым увеличивая эффективность фотосинтеза за счет снижения фотодыхания. Процесс концентрации CO 2 вокруг RuBisCO требует больше энергии, чем диффузия газов , но при определенных условиях – например, при высоких температурах (>25 °C), низких концентрациях CO 2 или высоких концентрациях кислорода – окупается снижением потерь. сахаров посредством фотодыхания.
Один тип метаболизма C 4 основан на так называемой анатомии Кранца . Это транспортирует CO 2 через внешний слой мезофилла через ряд органических молекул к центральным клеткам оболочки пучка, где CO 2 высвобождается. Таким образом, CO 2 концентрируется вблизи места деятельности Рубиско. Поскольку RuBisCO работает в среде с гораздо большим количеством CO 2 , чем было бы в противном случае, он работает более эффективно.
Второй механизм, фотосинтез CAM , временно отделяет фотосинтез от действия RuBisCO. RuBisCO действует только в течение дня, когда устьица закрыты и CO 2 выделяется в результате распада химического малата . Больше CO 2 затем извлекается из атмосферы, когда устьица открываются в прохладные и влажные ночи, что снижает потерю воды.
Эволюционный рекорд
[ редактировать ]Эти два пути, оказывая одинаковое влияние на RuBisCO, несколько раз развивались независимо – действительно, один C 4 возникал 62 раза в 18 различных семействах растений . Ряд «предварительных адаптаций», по-видимому, проложил путь для C 4 , что привело к его кластеризации в определенные клады: он чаще всего появлялся в растениях, которые уже имели такие особенности, как обширная ткань оболочки сосудистых пучков. [136] Многие потенциальные эволюционные пути, приводящие к C4 фенотипу , возможны и охарактеризованы с использованием байесовского вывода . [135] подтверждая, что нефотосинтетические адаптации часто служат эволюционными ступеньками для дальнейшей эволюции C 4 .
Конструкция C 4 используется некоторыми видами трав, тогда как CAM используется многими суккулентами и кактусами . Признак C 4 , судя по всему, появился в олигоцене , примерно 25–32 миллиона лет назад ; [137] однако они не стали экологически значимыми до миоцена , 6–7 миллионов лет назад . [138] Примечательно, что некоторые обугленные окаменелости сохраняют ткань, организованную в анатомию Кранца, с неповрежденными клетками оболочки пучка, [139] наличие метаболизма C 4 позволяя идентифицировать . Изотопные маркеры используются для определения их распределения и значимости. Растения C 3 преимущественно используют более легкий из двух изотопов углерода в атмосфере, 12 C, который с большей готовностью участвует в химических путях его фиксации. Поскольку метаболизм C 4 включает в себя еще один химический этап, этот эффект усиливается. Растительный материал можно проанализировать, чтобы определить соотношение более тяжелых 13 С до 12 C. Это соотношение обозначается δ 13 С. Растения C 3 в среднем примерно на 14 ‰ (частей на тысячу) легче атмосферного, а растения C 4 примерно на 28 ‰ легче. δ 13 C CAM-растений зависит от процентного содержания углерода, фиксируемого ночью по отношению к тому, что фиксируется днем: он ближе к растениям C 3 , если они фиксируют большую часть углерода днем, и ближе к растениям C 4 , если они фиксируют весь углерод ночью. . [140]
Исходного ископаемого материала в достаточном количестве для анализа самой травы мало, но лошади являются хорошим показателем. В рассматриваемый период они были широко распространены по всему миру и питались почти исключительно травами. В изотопной палеонтологии есть старая фраза: «Вы — то, что вы едите (плюс немного)» — она относится к тому факту, что организмы отражают изотопный состав того, что они едят, плюс небольшой поправочный коэффициент. Во всем мире имеются хорошие данные о конских зубах, а их δ 13 Запись C показывает резкий отрицательный перегиб примерно 6–7 миллионов лет назад , во время мессинского периода, который интерпретируется как результат появления растений C 4 в глобальном масштабе. [138]
Преимущество C 4
[ редактировать ]Хотя C 4 повышает эффективность RuBisCO, концентрация углерода очень энергозатратна. Это означает, что растения C 4 имеют преимущество перед организмами C 3 только в определенных условиях, а именно: при высоких температурах и небольшом количестве осадков. Растениям C 4 также необходим высокий уровень солнечного света, чтобы процветать. [141] Модели предполагают, что без лесных пожаров, уничтожающих деревья и кустарники, создающие тень, не останется места для C 4 . растений [142] Но лесные пожары происходили на протяжении 400 миллионов лет. В каменноугольном периоде (около 300 миллионов лет назад ) было общеизвестно высокое содержание кислорода – почти достаточное, чтобы обеспечить самовозгорание. [143] – и очень низкий уровень CO 2 , но никаких изотопных признаков C 4 обнаружено не было. Похоже, что внезапного толчка для подъема миоцена также не было. [ нужна ссылка ]
В миоцене атмосфера и климат были относительно стабильными. Во всяком случае, концентрация CO 2 постепенно увеличивалась от 14 до 9 миллионов лет назад, прежде чем стабилизироваться до концентрации, аналогичной голоцену. [144] Это говорит о том, что он не сыграл ключевой роли в вызове эволюции C4 . [137] Сами травы (группа, которая привела к наибольшему распространению C 4 ), вероятно, существовали уже 60 миллионов лет или больше, поэтому у них было достаточно времени для эволюции C 4 , [145] [146] который, в любом случае, присутствует в самых разных группах и, таким образом, развивался независимо. В Южной Азии наблюдается сильный сигнал об изменении климата; [137] увеличение засушливости – а следовательно, увеличение частоты и интенсивности пожаров – могло привести к увеличению важности лугов. [147] Однако это трудно совместить с показателями Северной Америки. [137] Возможно, сигнал полностью биологический, вызванный огнем. [148] вызванное ускорением эволюции травы, которое, как за счет усиления выветривания, так и за счет включения большего количества углерода в отложения, снизило уровень CO 2 в атмосфере . [148] Наконец, есть свидетельства того, что появление C 4 от 9 до 7 миллионов лет назад является предвзятым сигналом, который справедлив только для Северной Америки, откуда происходит большинство образцов; Новые данные свидетельствуют о том, что луга превратились в доминирующее состояние в Южной Америке как минимум 15 млн лет назад. [ нужна ссылка ]
Эволюция регуляции транскрипции
[ редактировать ]Факторы транскрипции и сети регуляции транскрипции играют ключевую роль в развитии растений и реакциях на стресс, а также в их эволюции. Во время посадки растений появилось множество новых семейств транскрипционных факторов, которые преимущественно встраиваются в сети многоклеточного развития, размножения и развития органов, способствуя более сложному морфогенезу наземных растений. [149]
Эволюция вторичного метаболизма
[ редактировать ]
Вторичные метаболиты представляют собой по существу низкомолекулярные соединения , иногда имеющие сложную структуру, не необходимые для нормальных процессов роста , развития или размножения . Они участвуют в таких разнообразных процессах, как иммунитет , борьба с травоядностью, опылителей привлечение , связь между растениями, поддержание симбиотических связей с почвенной флорой или повышение скорости оплодотворения , и, следовательно, важны с точки зрения эво-дево. Вторичные метаболиты структурно и функционально разнообразны, и, по оценкам, в процессе их производства могут быть задействованы сотни тысяч ферментов, причем около 15–25% генома кодирует эти ферменты, и каждый вид имеет свой уникальный арсенал вторичные метаболиты. [150] Многие из этих метаболитов, такие как салициловая кислота, имеют медицинское значение для человека. [ нужна ссылка ]
Цель производства такого количества вторичных метаболитов, при этом значительная часть метаболома посвящена этой активности, неясна. Предполагается, что большинство этих химических веществ способствуют выработке иммунитета и, как следствие, разнообразие этих метаболитов является результатом постоянной гонки вооружений между растениями и их паразитами. Некоторые доказательства подтверждают это дело. Центральный вопрос касается репродуктивных затрат на поддержание такого большого количества генов, отвечающих за производство вторичных метаболитов. Были предложены различные модели, исследующие этот аспект вопроса, но консенсус относительно масштабов затрат еще не достигнут; [151] поскольку до сих пор трудно предсказать, увеличит ли растение с большим количеством вторичных метаболитов свою выживаемость или репродуктивный успех по сравнению с другими растениями, находящимися поблизости. [ нужна ссылка ]
Производство вторичных метаболитов, по-видимому, возникло довольно рано в ходе эволюции. У растений они, по-видимому, распространились с помощью таких механизмов, как дупликация генов или эволюция новых генов. Более того, исследования показали, что разнообразие некоторых из этих соединений может быть выбрано положительно. Хотя роль эволюции новых генов в эволюции вторичного метаболизма очевидна, есть несколько примеров, когда новые метаболиты образовывались в результате небольших изменений в реакции. Например, было высказано предположение, что цианогенгликозиды неоднократно эволюционировали в разных линиях растений. Есть несколько таких примеров конвергентной эволюции . Например, ферменты синтеза лимонена ( терпена ) у покрытосеменных и голосеменных более похожи, чем на их собственные ферменты синтеза терпена. Это предполагает независимую эволюцию пути биосинтеза лимонена в этих двух линиях. [152]
Эволюция растительно-микробных взаимодействий
[ редактировать ]Происхождение микробов на Земле, восходящее к зарождению жизни более 3,5 миллиардов лет назад, указывает на то, что межмикробные взаимодействия постоянно развивались и диверсифицировались с течением времени, задолго до того, как растения начали колонизировать землю 450 миллионов лет назад. Таким образом, вполне вероятно, что межмикробные взаимодействия как внутри, так и между царствами представляют собой сильные движущие силы создания связанных с растениями микробных консорциумов на границе раздела почва-корни. Тем не менее, остается неясным, в какой степени эти взаимодействия в ризосфере/филлосфере и в эндофитных компартментах растения (т. е. внутри хозяина) формируют микробные комплексы в природе и является ли адаптация микробов к местообитаниям растений движущей силой стратегий межмикробного взаимодействия, специфичных для среды обитания, которые влияют на растительный фитнес. Более того, вклад конкурентных и кооперативных микроб-микробных взаимодействий в общую структуру сообщества по-прежнему трудно оценить в природе из-за сильного шума окружающей среды. [153]
См. также
[ редактировать ]- Эволюция травоядных
- Эволюционная история жизни
- Палеоботаника
- Эволюционная биология развития растений
- Хронология эволюции растений
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м Стюарт, Западная Нью-Йорк; Ротвелл, GW (1993). Палеоботаника и эволюция растений (2-е изд.). Издательство Кембриджского университета. ISBN 978-0-521-23315-6 .
- ^ Jump up to: а б Стротер, Пол К.; Баттисон, Лейла; Брейзер, Мартин Д.; Веллман, Чарльз Х. (2011). «Самые ранние неморские эукариоты Земли» . Природа . 473 (7348): 505–509. Бибкод : 2011Natur.473..505S . дои : 10.1038/nature09943 . ПМИД 21490597 . S2CID 4418860 .
- ^ Jump up to: а б с Кнаут, Л. Пол; Кеннеди, Мартин Дж. (2009). «Позднедокембрийское позеленение Земли». Природа . 460 (7256): 728–732. Бибкод : 2009Natur.460..728K . дои : 10.1038/nature08213 . ПМИД 19587681 . S2CID 4398942 .
- ^ Ротвелл, GW; Шеклер, SE; Гиллеспи, Вашингтон (1989). « Elkinsia gen. nov., позднедевонское голосеменное растение с купулированными семязачатками». Ботанический вестник . 150 (2): 170–189. дои : 10.1086/337763 . S2CID 84303226 .
- ^ Кенрик, П.; Крейн, PR (1997). Происхождение и ранняя диверсификация наземных растений. Кладистическое исследование . Вашингтон: Издательство Смитсоновского института. ISBN 978-1-56098-729-1 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Рэйвен, Дж.А.; Эдвардс, Д. (2001). «Корни: эволюционное происхождение и биогеохимическое значение» . Журнал экспериментальной ботаники . 52 (90001): 381–401. doi : 10.1093/jexbot/52.suppl_1.381 . ПМИД 11326045 .
- ^ Дель-Бем, Ле (2018). «Эволюция ксилоглюканов и террестриализация зеленых растений» . Новый фитолог . 219 (4): 1150–1153. дои : 10.1111/nph.15191 . hdl : 1843/36860 . ПМИД 29851097 .
- ^ Хекман, Д.С.; Гейзер, Д.М.; Эйделл, БР; Стауффер, РЛ; Кардос, Нидерланды; Хеджес, С.Б. (август 2001 г.). «Молекулярные доказательства ранней колонизации земли грибами и растениями». Наука . 293 (5532): 1129–1133. дои : 10.1126/science.1061457 . ПМИД 11498589 . S2CID 10127810 .
- ^ Баттисон, Лейла; Брейзер, Мартин Д. (август 2009 г.). «Исключительная сохранность ранних наземных сообществ в озерном фосфате один миллиард лет назад» (PDF) . В Смите, Мартин Р.; О'Брайен, Лорна Дж.; Карон, Жан-Бернар (ред.). Аннотация Том . Международная конференция по кембрийскому взрыву (Уолкотт, 2009 г.) . Торонто, Онтарио, Канада: Консорциум Burgess Shale (опубликовано 31 июля 2009 г.). ISBN 978-0-9812885-1-2 .
- ^ Баттистуцци, ФУ; Фейджао, А.; Хеджес, С.Б. (2004). «Геномная временная шкала эволюции прокариот: понимание происхождения метаногенеза, фототрофии и колонизации суши» . Эволюционная биология BMC . 4:44 . дои : 10.1186/1471-2148-4-44 . ПМЦ 533871 . ПМИД 15535883 .
- ^ Нельсен, Мэтью П.; Удача, Роберт; Бойс, К. Кевин; Лумбш, Х. Торстен; Ри, Ричард Х. (2019). «Нет подтверждения появлению лишайников до эволюции сосудистых растений». Геобиология . 18 (1).
- ^ Камп, ЛР; Павлов А.; Артур, Массачусетс (2005). «Массовый выброс сероводорода на поверхность океана и в атмосферу в периоды океанической аноксии». Геология . 33 (5): 397. Бибкод : 2005Гео....33..397К . дои : 10.1130/G21295.1 .
- ^ Баттерфилд, Нью-Джерси (2009). «Кислород, животные и вентиляция океана: альтернативный взгляд». Геобиология . 7 (1): 1–7. Бибкод : 2009Gbio....7....1B . дои : 10.1111/j.1472-4669.2009.00188.x . ПМИД 19200141 . S2CID 31074331 .
- ^ Даль, Таис В.; Аренс, Сюзанна К.М. (2020). «Влияние эволюции наземных растений на климат Земли и состояние оксигенации - междисциплинарный обзор». Химическая геология . 547 .
- ^ Стротер, ПК; Аль-Хаджри, С.; Траверс, А. (1996). «Новые свидетельства существования наземных растений из нижнего среднего ордовика Саудовской Аравии». Геология . 24 (1): 55–59. Бибкод : 1996Гео....24...55С . doi : 10.1130/0091-7613(1996)024<0055:NEFLPF>2.3.CO;2 .
- ^ Рубинштейн, CV; Джерриенн, П.; де ла Пуэнте, GS; Астини, РА; Стиманс, П. (2010). «Свидетельства существования наземных растений в раннем среднем ордовике в Аргентине (восточная Гондвана)» . Новый фитолог . 188 (2): 365–369. дои : 10.1111/j.1469-8137.2010.03433.x . hdl : 11336/55341 . ПМИД 20731783 .
- ^ Веллман, Чарльз Х.; Остерлофф, Питер Л.; Мохиуддин, Узма (2003). «Фрагменты древнейших наземных растений» (PDF) . Природа . 425 (6955): 282–285. Бибкод : 2003Natur.425..282W . дои : 10.1038/nature01884 . ПМИД 13679913 . S2CID 4383813 .
- ^ Стиманс, П.; Лепота К.; Маршаллб, CP; Ле Эриссек, А.; Яваукса, EJ (2010). «FTIR-характеристика химического состава силурийских миоспор (криптоспор и трилетных спор) из Готланда, Швеция» . Обзор палеоботаники и палинологии . 162 (4): 577–590. Бибкод : 2010RPaPa.162..577S . дои : 10.1016/j.revpalbo.2010.07.006 . S2CID 128900940 .
- ^ Стиманс, П.; Херисс, Л.; Мелвин, Дж.; Миллер, А.; Пэрис, Ф.; Верньерс, Дж.; Веллман, Х. (апрель 2009 г.). «Происхождение и распространение древнейших сосудистых наземных растений» . Наука . 324 (5925): 353. Бибкод : 2009Sci...324..353S . дои : 10.1126/science.1169659 . hdl : 1854/LU-697223 . ISSN 0036-8075 . ПМИД 19372423 . S2CID 206518080 .
- ^ Рубинштейн, Клаудия; Вайда, Виви (2019). «Балтика – колыбель ранних наземных растений? Древнейшая запись трилетных спор и разнообразных комплексов криптоспор; свидетельства ордовикских последовательностей Швеции» . GFF (Журнал Геологического общества Швеции) . 141 (3): 181–190. Бибкод : 2019GFF...141..181R . дои : 10.1080/11035897.2019.1636860 . hdl : 11336/124409 .
- ^ Jump up to: а б с Грей, Дж.; Чалонер, В.Г.; Уэстолл, Т.С. (1985). «Летопись микрофоссилий ранних наземных растений: достижения в понимании ранней террестриализации, 1970–1984». Философские труды Королевского общества Б. 309 (1138): 167–195. Бибкод : 1985РСТБ.309..167Г . дои : 10.1098/rstb.1985.0077 . JSTOR 2396358 .
- ^ Томеску, АМФ (2006). «Водно-болотные угодья до появления трахеофитов: слоевищные наземные сообщества биоты раннего силура Пассаж-Крик (Вирджиния)» (PDF) . дои : 10.1130/2006.2399(02) . ISBN 9780813723990 . Проверено 28 мая 2014 г.
{{cite book}}:|journal=игнорируется ( помощь ) ; Отсутствует или пусто|title=( помощь ) - ^ Томеску, АМФ; Онеггер, Р.; Ротвелл, GW (2008). «Самая ранняя летопись окаменелостей консорциумов бактериально-цианобактериальных матов: ранняя силурийская биота Пассаж-Крик (440 млн лет назад, Вирджиния, США)». Геобиология . 6 (2): 120–124. Бибкод : 2008Gbio....6..120T . дои : 10.1111/j.1472-4669.2007.00143.x . ПМИД 18380874 . S2CID 10555223 .
- ^ «Первые наземные растения и грибы изменили климат Земли, открыв путь к взрывной эволюции наземных животных, как показывают новые исследования генов» . science.psu.edu . Архивировано из оригинала 15 декабря 2019 года . Проверено 9 апреля 2018 г.
- ^ Жарский, Ю.; Жарский, В.; Ганачек, М.; Жарский, В. (27 января 2022 г.). «Криогенные ледниковые среды обитания как колыбель террестриализации растений - происхождение раскола Anydropphytes и Zygnematophyceae» . Границы в науке о растениях . 12 . Границы : 735020. doi : 10.3389/fpls.2021.735020 . ISSN 1664-462X . ПМЦ 8829067 . ПМИД 35154170 .
- ^ Камп, Лео Р. (16 января 2008 г.). «Повышение содержания кислорода в атмосфере» . Природа . 451 (7176): 277–278. Бибкод : 2008Natur.451..277K . дои : 10.1038/nature06587 . ПМИД 18202642 . S2CID 5505876 .
- ^ Гласспул, Ай-Джей; Эдвардс, Д.; Акс, Л. (2004). «Древесный уголь в силурийском периоде как свидетельство самого раннего лесного пожара». Геология . 32 (5): 381–383. Бибкод : 2004Geo....32..381G . дои : 10.1130/G20363.1 .
- ^ Бернштейн, Х.; Байерс, Г.С.; Мишо, RE (1981). «Эволюция полового размножения: важность восстановления, комплементации и вариаций ДНК». Американский натуралист . 117 (4): 537–549. дои : 10.1086/283734 . S2CID 84568130 .
- ^ Мишо, RE; Гейли, ТВ (1992). «Маскировка мутаций и эволюция пола». Американский натуралист . 139 (4): 706–734. дои : 10.1086/285354 . S2CID 85407883 .
- ^ Шовеньи, Питер; Рикка, Мариана; Хок, Жофия; Шоу, Джонатан А.; Симидзу, Кентаро К.; Вагнер, Андреас (2013). «Отбор на гаплоидных стадиях жизни не более эффективен, чем на диплоидных стадиях жизни покрытосеменных растений и мхов» . Молекулярная биология и эволюция . 30 (8): 1929–39. дои : 10.1093/molbev/mst095 . ПМИД 23686659 .
- ^ Jump up to: а б с д Бойс, СК (2008). «Насколько зеленой была Куксония ? Важность размера для понимания ранней эволюции физиологии сосудистых растений». Палеобиология . 34 (2): 179–194. doi : 10.1666/0094-8373(2008)034[0179:HGWCTI]2.0.CO;2 . S2CID 36688488 .
- ^ Керп, Х.; Тревин, Нью-Хэмпшир; Хасс, Х. (2004). «Новые гаметофиты из раннедевонских рини-кремней». Труды Королевского общества Эдинбурга . 94 (4): 411–428. дои : 10.1017/s026359330000078x . S2CID 128629425 .
- ^ Тейлор, Теннесси; Керп, Х.; Хасс, Х. (2005). «Биология истории жизни ранних наземных растений: расшифровка фазы гаметофита» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (16): 5892–5897. Бибкод : 2005PNAS..102.5892T . дои : 10.1073/pnas.0501985102 . ПМК 556298 . ПМИД 15809414 .
- ^ Холлоуэй, Джон Э. (1939). «Гаметофит, зародыш и молодое корневище Psilotum triquetrum (Swartz)». Анналы ботаники . 3 (2): 313–336. doi : 10.1093/oxfordjournals.aob.a085063 .
- ^ Кидстон, Р. и Лэнг, WH (1920), «О растениях из старого красного песчаника, показывающих структуру, из пласта Райни Черт, Абердиншир. Часть II. Дополнительные примечания к Rhynia gwynne-vaughani , Кидстону и Лангу; с описаниями Rhynia major , n.sp. и Hornea lignieri , ng, n.sp" , Труды Королевского общества Эдинбурга , 52 (3): 603–627, doi : 10.1017/s0080456800004488 , S2CID 130329822
- ^ Реми, В.; Тейлор, Теннесси; Хасс, Х.; Керп, Х. (1994). «Везикулярно-арбускулярная микориза возрастом 4ста миллионов лет» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (25): 11841–11843. Бибкод : 1994PNAS...9111841R . дои : 10.1073/pnas.91.25.11841 . ПМЦ 45331 . ПМИД 11607500 .
- ^ Бонфанте, Паола; Жанр, Андреа (сентябрь 2008 г.). «Растения и арбускулярные микоризные грибы: взгляд на эволюционное развитие» . Тенденции в науке о растениях . 13 (9): 492–498. doi : 10.1016/j.tplants.2008.07.001 . ISSN 1360-1385 . ПМИД 18701339 .
- ^ Шюслер, А.; и др. (2001). «Новый тип грибов Glomeromycota : филогения и эволюция» . Микол. Рез . 105 (12): 1416. doi : 10.1017/S0953756201005196 .
- ^ Jump up to: а б Смит, Салли Э.; Прочтите, Дэвид Дж. (2008). Микоризный симбиоз (3-е изд.). Академическая пресса. ISBN 9780123705266 .
- ^ Саймон, Л.; Буске, Ж.; Левеск, Колорадо; Лалонд, М. (1993). «Происхождение и разнообразие эндомикоризных грибов и совпадение с сосудистыми наземными растениями». Природа . 363 (6424): 67–69. Бибкод : 1993Natur.363...67S . дои : 10.1038/363067a0 . S2CID 4319766 .
- ^ Саймон, Л.; Буске, Ж.; Левеск, Колорадо; Лалонд, М. (1993). «Происхождение и разнообразие эндомикоризных грибов и совпадение с сосудистыми наземными растениями». Природа . 363 (6424): 67–69. Бибкод : 1993Natur.363...67S . дои : 10.1038/363067a0 . S2CID 4319766 .
- ^ Хамфрис, CP; Фрэнкс, П.Дж.; Рис, М.; Бидартондо, Мичиган; Лик, младший; Бирлинг, диджей (2010). «Мутуалистический микоризоподобный симбиоз у древнейшей группы наземных растений» . Природные коммуникации . 1 (8): 103. Бибкод : 2010NatCo...1..103H . дои : 10.1038/ncomms1105 . ПМИД 21045821 .
- ^ Брундретт, MC (2002). «Коэволюция корней и микоризы наземных растений» . Новый фитолог . 154 (2): 275–304. дои : 10.1046/j.1469-8137.2002.00397.x . ПМИД 33873429 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р Сперри, Дж. С. (2003). «Эволюция водного транспорта и структуры ксилемы». Международный журнал наук о растениях . 164 (3): С115–С127. дои : 10.1086/368398 . JSTOR 3691719 . S2CID 15314720 .
- ^ Jump up to: а б с д Рэйвен, Джон (1977). «Эволюция сосудистых наземных растений в отношении процессов надклеточного транспорта». Достижения в ботанических исследованиях . 1977 : 153–219. дои : 10.1016/S0065-2296(08)60361-4 . ISBN 9780120059058 .
- ^ Цю, ЮЛ; Ли, Л.; Ван, Б.; Чен, З.; Кноп, В.; Грот-малонек, М.; Домбровская, О.; Ли, Дж.; Кент, Л.; Рест, Дж.; и др. (2006). «Самые глубокие различия у наземных растений, выявленные на основе филогеномных данных» . Труды Национальной академии наук . 103 (42): 15511–6. Бибкод : 2006PNAS..10315511Q . дои : 10.1073/pnas.0603335103 . ПМК 1622854 . ПМИД 17030812 .
- ^ Эдвардс, Д.; Дэвис, КЛ; Акс, Л. (1992). «Сосудистая проводящая нить у раннего наземного растения Cooksonia ». Природа . 357 (6380): 683–685. Бибкод : 1992Natur.357..683E . дои : 10.1038/357683a0 . S2CID 4264332 .
- ^ Никлас, К.Дж.; Смоковит, В. (1983). «Свидетельства наличия проводящей нити у раннесилурийских (лландоверских) растений: значение для эволюции наземных растений». Палеобиология . 9 (2): 126–137. Бибкод : 1983Pbio....9..126N . дои : 10.1017/S009483730000751X . JSTOR 2400461 . S2CID 35550235 .
- ^ Jump up to: а б с д и Никлас, KJ (1985). «Эволюция диаметра трахеиды у ранних сосудистых растений и ее влияние на гидравлическую проводимость первичной цепи ксилемы». Эволюция . 39 (5): 1110–1122. дои : 10.2307/2408738 . JSTOR 2408738 . ПМИД 28561493 .
- ^ Никлас, К.; Пратт, Л. (1980). «Доказательства наличия лигниноподобных компонентов в окаменелостях растений раннего силура (лландовера)». Наука . 209 (4454): 396–397. Бибкод : 1980Sci...209..396N . дои : 10.1126/science.209.4454.396 . ПМИД 17747811 . S2CID 46073056 .
- ^ Питтерманн Дж.; Сперри Дж.С.; Хаке У.Г.; Уиллер Дж.К.; Сиккема Э.Х. (декабрь 2005 г.). «Ямы Торус-Марго помогают хвойным деревьям конкурировать с покрытосеменными». Научный журнал . 310 (5756): 1924. doi : 10.1126/science.1120479 . ПМИД 16373568 . S2CID 28339481 .
- ^ Почему рождественские елки не вымерли
- ^ Jump up to: а б Кенрик, Пол (2001). «Открывая новый лист». Природа . 410 (6826): 309–310. Бибкод : 2001Natur.410..309K . дои : 10.1038/35066649 . ПМИД 11268183 . S2CID 4335355 .
- ^ Jump up to: а б с Берлинг Д .; Осборн, CP; Чалонер, WG (2001). «Эволюция формы листьев наземных растений связана с уменьшением содержания CO 2 в атмосфере в эпоху позднего палеозоя» (PDF) . Природа . 410 (6826): 352–354. Бибкод : 2001Natur.410..352B . дои : 10.1038/35066546 . ПМИД 11268207 . S2CID 4386118 .
- ^ Крейн, PR; Кенрик, П. (1997). «Отклоненное развитие репродуктивных органов: источник морфологических инноваций наземных растений». Систематика и эволюция растений . 206 (1): 161–174. Бибкод : 1997PSyEv.206..161C . дои : 10.1007/BF00987946 . S2CID 23626358 .
- ^ Циммерманн, В. (1959). Филогения растений (2-е изд.). Штутгарт: Густав Фишер Верлаг.
- ^ Пьяцца, П.; и др. (2005). «Эволюция механизмов развития листьев». Новый фитолог . 167 (3): 693–710. дои : 10.1111/j.1469-8137.2005.01466.x . ПМИД 16101907 .
- ^ Хагеманн, В. 1976. Являются ли папоротники листоносными? Альтернатива теломной теории.Систематика и эволюция растений 124: 251–277.
- ^ Хагеманн, В. (1999). «К организмической концепции наземных растений: маргинальная бластозона и развитие растительного тела избранных фрондозных гаметофитов печеночников и папоротников». Систематика и эволюция растений . 216 (1–2): 81–133. Бибкод : 1999PSyEv.216...81H . дои : 10.1007/bf00985102 . S2CID 21483458 .
- ^ Саттлер, Р. (1992). «Морфология процесса: структурная динамика в развитии и эволюции». Канадский журнал ботаники . 70 (4): 708–714. дои : 10.1139/b92-091 .
- ^ Саттлер, Р. 1998. О происхождении симметрии, ветвления и филлотаксиса у наземных растений. В: Р.В. Жан и Д. Барабе (ред.) Симметрия в растениях. World Scientific, Сингапур, стр. 775–793.
- ^ Джеймс, Пи Джей (2009). « Дерево и лист: другой ракурс». Линней . 25:17 .
- ^ «Взгляд на на CO 2 теорию ранней эволюции листьев, основанную » . Архивировано из оригинала 29 июня 2011 г. Проверено 8 марта 2011 г.
- ^ Котык, МЭ; Бейсингер, Дж. Ф.; Гензель, П.Г.; де Фрейтас, Т. А. (2002). «Морфологически сложные растительные макрофоссилии позднего силура арктической Канады» . Американский журнал ботаники . 89 (6): 1004–1013. дои : 10.3732/ajb.89.6.1004 . PMID 21665700 .
- ^ Рикардс, РБ (2000). «Возраст самых ранних плаунов: силурийская флора Baragwanathia в Виктории, Австралия» (аннотация) . Геологический журнал . 137 (2): 207–209. Бибкод : 2000GeoM..137..207R . дои : 10.1017/S0016756800003800 . S2CID 131287538 . Проверено 25 октября 2007 г.
- ^ Jump up to: а б с д и Каплан, Д.Р. (2001). «Наука о морфологии растений: определение, история и роль в современной биологии». Американский журнал ботаники . 88 (10): 1711–1741. дои : 10.2307/3558347 . JSTOR 3558347 . ПМИД 21669604 .
- ^ Тейлор, Теннесси; Хасс, Х.; Керп, Х.; Крингс, М.; Ханлин, RT (2005). «Пертециальные аскомицеты из кремня Рини возрастом 400 миллионов лет: пример наследственного полиморфизма». Микология . 97 (1): 269–285. дои : 10.3852/микология.97.1.269 . hdl : 1808/16786 . ПМИД 16389979 .
- ^ Jump up to: а б с Бойс, СК; Нолл, АХ (2002). «Эволюция потенциала развития и множественное независимое происхождение листьев палеозойских сосудистых растений». Палеобиология . 28 (1): 70–100. doi : 10.1666/0094-8373(2002)028<0070:EODPAT>2.0.CO;2 . S2CID 1650492 .
- ^ Хао, С.; Бек, CB; Деминг, В. (2003). «Структура самых ранних листьев: адаптация к высоким концентрациям атмосферного CO 2 ». Международный журнал наук о растениях . 164 (1): 71–75. дои : 10.1086/344557 . S2CID 84741832 .
- ^ Бернер, РА; Котавала, З. (2001). «Геокарб III: пересмотренная модель атмосферного CO 2 в фанерозойское время» . Американский научный журнал . 301 (2): 182–204. Бибкод : 2001AmJS..301..182B . CiteSeerX 10.1.1.393.582 . дои : 10.2475/ajs.301.2.182 . Архивировано из оригинала (аннотация) 29 января 2009 г. Проверено 7 апреля 2008 г.
- ^ Шеллито, CJ; Слоан, LC (2006). «Реконструкция утраченного эоценового рая: Часть I. Моделирование изменения в глобальном распределении флоры при начальном термическом максимуме эоцена». Глобальные и планетарные изменения . 50 (1–2): 1–17. Бибкод : 2006GPC....50....1S . дои : 10.1016/j.gloplacha.2005.08.001 .
- ^ Аэртс, Р. (1995). «Преимущества вечнозелености». Тенденции в экологии и эволюции . 10 (10): 402–407. дои : 10.1016/S0169-5347(00)89156-9 . ПМИД 21237084 .
- ^ Лабандейра, CC; Дилчер, Д.Л.; Дэвис, доктор медицинских наук; Вагнер, Д.Л. (1994). «Девяносто семь миллионов лет ассоциации покрытосеменных растений и насекомых: палеобиологическое понимание значения коэволюции» . ПНАС . 91 (25): 12278–12282. Бибкод : 1994PNAS...9112278L . дои : 10.1073/pnas.91.25.12278 . ПМК 45420 . ПМИД 11607501 .
- ^ де Соуза, Рошель; Амброзини, Адриана; Пассалья, член парламента Лучане (декабрь 2015 г.). «Бактерии, стимулирующие рост растений, в качестве инокулянтов в сельскохозяйственных почвах» . Генетика и молекулярная биология . 38 (4): 401–19. дои : 10.1590/S1415-475738420150053 . ПМЦ 4763327 . ПМИД 26537605 .
- ^ Мора, CI; Дриз, С.Г.; Коларуссо, Луизиана (1996). в атмосфере среднего и позднего палеозоя «Уровни CO 2 от карбоната почвы и органических веществ». Наука . 271 (5252): 1105–1107. Бибкод : 1996Sci...271.1105M . дои : 10.1126/science.271.5252.1105 . S2CID 128479221 .
- ^ Бернер, Р.А. (1994). «GEOCARB II: Пересмотренная модель атмосферного CO 2 в фанерозойское время» . Являюсь. J. Sci . 294 (1): 56–91. Бибкод : 1994AmJS..294...56B . дои : 10.2475/ajs.294.1.56 .
- ^ Jump up to: а б Алгео, ТиДжей; Бернер, РА; Мейнард, Дж.Б.; Шеклер, SE; Архивы GSA Today (1995). «Позднедевонские океанические аноксические явления и биотические кризисы: «коренятся» в эволюции сосудистых наземных растений?» . ГСА сегодня . 5 (3). Архивировано из оригинала 1 мая 2015 г. Проверено 29 августа 2019 г.
- ^ Реталлак, Дж.Дж. (1986). Райт, вице-президент (ред.). Палеопочвы: их распознавание и интерпретация . Оксфорд: Блэквелл.
- ^ «Трахеофита-2» . Палеос – Жизнь сквозь глубокое время .
- ^ Ричард М. Бейтман, Питер Р. Крейн, Уильям А. ДиМишель, Пол Р. Кенрик, Ник П. Роу, Томас Спек и Уильям Э. Стейн (1998). «РАННЯЯ ЭВОЛЮЦИЯ НАЗЕМНЫХ РАСТЕНИЙ: филогения, физиология и экология первичной земной радиации». Ежегодный обзор экологии и систематики . 29 : 263–292. doi : 10.1146/annurev.ecolsys.29.1.263 . S2CID 44508826 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хетерингтон, Александр Дж; Долан, Лиам (2017). «Двусторонне-симметричные оси с ризоидами составили корневую структуру общего предка сосудистых растений» . Философские труды Королевского общества B: Биологические науки . 373 (1739): 20170042. doi : 10.1098/rstb.2017.0042 . ПМЦ 5745339 . ПМИД 29254968 .
- ^ Jump up to: а б с д и ж г час я Алгео, ТиДжей; Шеклер, SE (1998). «Наземно-морские телекоммуникации в девоне: связи между эволюцией наземных растений, процессами выветривания и морскими бескислородными явлениями» . Философские труды Королевского общества Б. 353 (1365): 113–130. дои : 10.1098/rstb.1998.0195 . ПМК 1692181 .
- ^ Хетерингтон, Александр Дж.; Долан, Лиам (2018). «Ступенчатое и самостоятельное возникновение корней у наземных растений» . Природа . 561 (7722): 235–238. Бибкод : 2018Natur.561..235H . дои : 10.1038/s41586-018-0445-z . ПМК 6175059 . ПМИД 30135586 .
- ^ Кудерт, Джон; Энн Дивар; Гаэтан Дрок; Паскаль Гаунтлет (2012). «Филогения ASL/LBD предполагает, что генетические механизмы зарождения корней после ауксина различны у ликофитов и эуфиллофитов» . Молекулярная биология и эволюция . 30 (3): 569–72. дои : 10.1093/molbev/mss250 . ISSN 0737-4038 . ПМИД 23112232 .
- ^ Jump up to: а б Кенрик, П.; Крейн, PR (1997). «Происхождение и ранняя эволюция растений на суше». Природа . 389 (6646): 33–39. Бибкод : 1997Natur.389...33K . дои : 10.1038/37918 . S2CID 3866183 .
- ^ Жерриен, Филипп; Гензель, Патрисия Г.; Струллу-Дерриен, Кристина; Лардо, Юбер; Стиманс, Филипп; Престианни, Сирилл (2011). «Простой тип древесины у двух растений раннего девона». Наука . 333 (6044): 837. Бибкод : 2011Sci...333..837G . дои : 10.1126/science.1208882 . hdl : 2268/97121 . ПМИД 21836008 . S2CID 23513139 .
- ^ Кинвер, Марк (16 августа 2011 г.). « Образцы «ранней древесины» меняют историю растений» . Новости Би-би-си .
- ^ Штейн, МЫ; Маннолини, Ф.; Херник, Л.В.; Лендинг, Э.; Берри, CM (2007). «Гигантские деревья кладоксилопсиды раскрывают загадку самых ранних лесных пней на Земле в Гильбоа». Природа . 446 (7138): 904–907. Бибкод : 2007Natur.446..904S . дои : 10.1038/nature05705 . ПМИД 17443185 . S2CID 2575688 .
- ^ Реталлак, Дж.Дж.; Кэтт, Дж.А.; Чалонер, WG (1985). «Ископаемые почвы как основа для интерпретации появления крупных растений и животных на суше [и обсуждение]». Философские труды Королевского общества Б. 309 (1138): 105–142. Бибкод : 1985РСТБ.309..105Р . дои : 10.1098/rstb.1985.0074 . JSTOR 2396355 .
- ^ Данненхоффер, Дж. М.; Бонамо, премьер-министр (1989). « Rellimia thomsonii из Живета Нью-Йорка: вторичный рост в трех порядках ветвления». Американский журнал ботаники . 76 (9): 1312–1325. дои : 10.2307/2444557 . JSTOR 2444557 .
- ^ Дэвис, П; Кенрик, П. (2004). Ископаемые растения . Смитсоновские книги, ISBN Вашингтона, округ Колумбия 978-1-58834-156-3 .
- ^ Донохью, MJ (2005). «Ключевые инновации, конвергенция и успех: макроэволюционные уроки филогении растений» . Палеобиология . 31 (2): 77–93. doi : 10.1666/0094-8373(2005)031[0077:KICASM]2.0.CO;2 . S2CID 36988476 .
- ^ Jump up to: а б с Боу, LM; Пальто, Г.; Депамфилис, CW (2000). «Филогения семенных растений на основе всех трех геномных компартментов: современные голосеменные растения монофилетичны, а ближайшие родственники Gnetales - хвойные» . Труды Национальной академии наук . 97 (8): 4092–4097. Бибкод : 2000PNAS...97.4092B . дои : 10.1073/pnas.97.8.4092 . ЧВК 18159 . ПМИД 10760278 .
- ^ Jump up to: а б с Чау, С.М.; Паркинсон, CL; Ченг, Ю.; Винсент, ТМ; Палмер, доктор юридических наук (2000). «Филогения семенных растений, сделанная на основе всех трех геномов растений: монофилия современных голосеменных растений и происхождение Gnetales от хвойных» . Труды Национальной академии наук . 97 (8): 4086–91. Бибкод : 2000PNAS...97.4086C . дои : 10.1073/pnas.97.8.4086 . ЧВК 18157 . ПМИД 10760277 .
- ^ Jump up to: а б с Солтис, Делавэр; Солтис, П.С.; Занис, MJ (2002). «Филогения семенных растений на основе данных восьми генов». Американский журнал ботаники . 89 (10): 1670–81. дои : 10.3732/ajb.89.10.1670 . ПМИД 21665594 .
- ^ Фриис, Э.М.; Педерсен, КР; Крейн, PR (2006). «Меловые цветки покрытосеменных: инновации и эволюция в размножении растений». Палеогеография, Палеоклиматология, Палеоэкология . 232 (2–4): 251–293. Бибкод : 2006PPP...232..251F . дои : 10.1016/j.palaeo.2005.07.006 .
- ^ Jump up to: а б Хилтон, Дж.; Бейтман, РМ (2006). «Птеридоспермы являются основой филогении семенных растений». Журнал Ботанического общества Торри . 133 (1): 119–168. doi : 10.3159/1095-5674(2006)133[119:PATBOS]2.0.CO;2 . S2CID 86395036 .
- ^ Jump up to: а б Бейтман, РМ; Хилтон, Дж.; Рудалл, П.Дж. (2006). «Морфологический и молекулярно-филогенетический контекст покрытосеменных: сравнение подходов «сверху вниз» и «снизу вверх», используемых для вывода о вероятных характеристиках первых цветков» . Журнал экспериментальной ботаники . 57 (13): 3471–503. дои : 10.1093/jxb/erl128 . ПМИД 17056677 .
- ^ Jump up to: а б с д Фрелих, М.В.; Чейз, MW (2007). «После десяти лет прогресса происхождение покрытосеменных растений все еще остается большой загадкой». Природа . 450 (7173): 1184–1189. Бибкод : 2007Natur.450.1184F . дои : 10.1038/nature06393 . ПМИД 18097399 . S2CID 4373983 .
- ^ Магальон, Сусана; Кастильо, Аманда (1 января 2009 г.). «Диверсификация покрытосеменных растений во времени» . Американский журнал ботаники . 96 (1): 349–365. дои : 10.3732/ajb.0800060 . ПМИД 21628193 . Проверено 27 апреля 2021 г.
- ^ Джерриенн, П.; Мейер-Берто, Б.; Фэйрон-Демаре, М.; Стрил, М.; Стиманс, П. (2011). «Научный журнал» . Рункария — предшественник семенного растения среднего девона . 306 (5697). Американская ассоциация содействия развитию науки: 856–858. дои : 10.1126/science.1102491 . ПМИД 15514154 . S2CID 34269432 .
- ^ Ван, Деминг; Лю, Ле; Го, Юн; Сюэ, Цзиньчжуан; Мэн, Мейсен (2015). «Плодородный орган позднего девона, имеющий сходство с семенными растениями из Китая» . Научные отчеты . 5 : 10736. Бибкод : 2015NatSR...510736W . дои : 10.1038/srep10736 . ПМЦ 4448223 . ПМИД 26022973 .
- ^ Jump up to: а б с Мейпс, Г.; Ротвелл, GW; Хаворт, Монтана (1989). «Эволюция покоя семян». Природа . 337 (6208): 645–646. Бибкод : 1989Natur.337..645M . дои : 10.1038/337645a0 . S2CID 4358761 .
- ^ Бава, Канзас; Уэбб, CJ (1984). «Аборт цветов, фруктов и семян у тропических лесных деревьев. Последствия для эволюции отцовских и материнских репродуктивных моделей». Американский журнал ботаники . 71 (5): 736–751. дои : 10.2307/2443371 . JSTOR 2443371 .
- ^ Чеа, Канзас; Осборн, ди-джей (апрель 1978 г.). «Повреждения ДНК возникают при потере жизнеспособности зародышей стареющих семян ржи». Природа . 272 (5654): 593–599. Бибкод : 1978Natur.272..593C . дои : 10.1038/272593a0 . ПМИД 19213149 . S2CID 4208828 .
- ^ Коппен, Г.; Верчаево, Л. (2001). «Щелочной гель-электрофорез одиночных клеток/кометный анализ: способ изучения репарации ДНК в корешковых клетках прорастающих Vicia faba». Фолиа Биол. (Прага) . 47 (2): 50–54. ПМИД 11321247 .
- ^ Брей, СМ; Уэст, CE (декабрь 2005 г.). «Механизмы восстановления ДНК у растений: важнейшие сенсоры и эффекторы для поддержания целостности генома». Новый фитолог . 168 (3): 511–28. дои : 10.1111/j.1469-8137.2005.01548.x . ПМИД 16313635 .
- ^ Бернштейн С., Бернштейн Х. (1991) Старение, пол и восстановление ДНК. Академическое издательство, Сан-Диего. ISBN 0120928604 ISBN 978-0120928606
- ^ Филд, Т.С.; Бродрибб, Ти Джей; Иглесиас, А.; Шатле, Д.С.; Бареш, А.; Апчерч, Греция; Гомес, Б.; Мор, БАР; Койффард, К.; Квачек Дж.; Харамилло, К. (2011). «Ископаемые свидетельства меловой эскалации эволюции жилок листьев покрытосеменных» . Труды Национальной академии наук . 108 (20): 8363–8366. Бибкод : 2011PNAS..108.8363F . дои : 10.1073/pnas.1014456108 . ПМК 3100944 . ПМИД 21536892 .
- ^ Jump up to: а б Лоутон-Раух, А.; Альварес-Буйя, ER; Пуруганан, доктор медицины (2000). «Молекулярная эволюция развития цветов». Тенденции экологии и эволюции 15 (4): 144–1 дои : 10.1016/S0169-5347(99) 01816-9 ПМИД 10717683 .
- ^ Китайская академия наук (18 декабря 2018 г.). «Цветы возникли на 50 миллионов лет раньше, чем считалось ранее» . ЭврекАлерт! . Проверено 18 декабря 2018 г.
- ^ Койро, Марио; Дойл, Джеймс А.; Хилтон, Джейсон (25 января 2019 г.). «Насколько глубок конфликт между молекулярными и ископаемыми данными о возрасте покрытосеменных?» . Новый фитолог . 223 (1): 83–99. дои : 10.1111/nph.15708 . ПМИД 30681148 .
- ^ Нам, Дж.; Депамфилис, CW; Ма, Х; Ней, М (2003). «Древность и эволюция семейства генов MADS-Box, контролирующих развитие цветков у растений» . Мол. Биол. Эвол . 20 (9): 1435–1447. дои : 10.1093/molbev/msg152 . ПМИД 12777513 .
- ^ Jump up to: а б с Крепет, WL (2000). «Прогресс в понимании истории, успеха и взаимоотношений покрытосеменных: отвратительный «загадочный феномен» Дарвина » . Труды Национальной академии наук . 97 (24): 12939–41. Бибкод : 2000PNAS...9712939C . дои : 10.1073/pnas.97.24.12939 . ПМК 34068 . ПМИД 11087846 .
- ^ Сан, Г.; Джи, К.; Дилчер, Д.Л.; Чжэн, С.; Никсон, КЦ; Ван, X. (2002). «Архефруктовые, новое базальное семейство покрытосеменных». Наука . 296 (5569): 899–904. Бибкод : 2002Sci...296..899S . дои : 10.1126/science.1069439 . ПМИД 11988572 . S2CID 1910388 .
- ^ На самом деле, Archaefructus , вероятно, не имел настоящих цветов: см. Фриис, Э.М.; Дойл, Дж. А.; Эндресс, ПК; Ленг, К. (2003). «Archaefructus - предшественник покрытосеменных или специализированные ранние покрытосеменные?». Тенденции в науке о растениях . 8 (8): 369–373. дои : 10.1016/S1360-1385(03)00161-4 . ПМИД 12927969 .
- ^ Jump up to: а б Крыло, СЛ; Баучер, Л.Д. (1998). «Экологические аспекты радиации меловых цветковых растений». Ежегодный обзор наук о Земле и планетах . 26 (1): 379–421. Бибкод : 1998AREPS..26..379W . дои : 10.1146/annurev.earth.26.1.379 .
- ^ Jump up to: а б Филд, Т.С.; Аренс, Северная Каролина; Дойл, Дж. А.; Доусон, штат Техас; Донохью, MJ (2004). «Темно и тревожно: новый образ ранней экологии покрытосеменных» . Палеобиология . 30 (1): 82–107. doi : 10.1666/0094-8373(2004)030<0082:DADANI>2.0.CO;2 . S2CID 42855952 .
- ^ Габботт, профессор Сара (1 августа 2017 г.). «Первый цветок выглядел так?» . Новости Би-би-си . Проверено 1 августа 2017 г.
- ^ Соке, Эрве; и др. (1 августа 2017 г.). «Предковый цветок покрытосеменных и его раннее разнообразие» . Природные коммуникации . 8 : 16047. Бибкод : 2017NatCo...816047S . дои : 10.1038/ncomms16047 . ПМЦ 5543309 . ПМИД 28763051 .
- ^ Проект «Геном амбореллы» (20 декабря 2013 г.). « Геном амбореллы и эволюция цветковых растений» . Наука . 342 (6165): 1241089. doi : 10.1126/science.1241089 . ПМИД 24357323 . S2CID 202600898 . Проверено 1 февраля 2020 г.
- ^ Зукколо, А.; Бауэрс, Джон Э; Эстилл, Джеймс С.; и др. (2011). «Физическая карта генома Amborella trichopoda проливает свет на эволюцию структуры генома покрытосеменных» . Геномная биология . 12 (5): Р48. дои : 10.1186/gb-2011-12-5-r48 . ПМК 3219971 . ПМИД 21619600 .
- ^ Медарг, Н.Г.; Янофски, М. (март 2001 г.). «Функция и эволюция семейства генов MADS-бокса растений». Обзоры природы Генетика . 2 (3): 186–195. дои : 10.1038/35056041 . ПМИД 11256070 . S2CID 7399106 .
- ^ Джагер, Мюриэль; Хасанин, Александр; Мануэль, Майкл; и др. (2003). «Гены MADS-Box у гинкго двулопастного и эволюция семейства AGAMOUS» . Молекулярная биология и эволюция . 20 (5): 842–854. дои : 10.1093/molbev/msg089 . ПМИД 12679535 .
- ^ Трансляционная биология: от цветов арабидопсиса до архитектуры соцветий травы. Бет Э. Томпсон* и Сара Хейк, 2009 г. , Физиология растений, 149:38–45.
- ^ Китахара, К.; Мацумото, С. (2000). «Гены MADS-бокса розы 'MASAKO C1 и D1', гомологичные генам цветочной идентичности класса C». Наука о растениях . 151 (2): 121–134. дои : 10.1016/S0168-9452(99)00206-X . ПМИД 10808068 .
- ^ Катер, М.; и др. (1998). «Множественные АГАМНЫЕ гомологи огурца и петунии различаются по своей способности индуцировать судьбу репродуктивных органов» . Растительная клетка . 10 (2): 171–182. дои : 10.1105/tpc.10.2.171 . ПМЦ 143982 . ПМИД 9490741 .
- ^ Солтис, Д.; и др. (2007). «Цветочный геном: эволюционная история дупликации генов и изменения моделей экспрессии генов». Тенденции растениеводства . 12 (8): 358–367. doi : 10.1016/j.tplants.2007.06.012 . ПМИД 17658290 .
- ^ Паттерхилл, Джо; Лори, Ребекка; Макнайт, Ричард (2004). «Пришло время цвести: генетический контроль времени цветения» . Биоэссе . 26 (4): 353–363. doi : 10.1002/bies.20021 . ПМИД 15057934 . S2CID 43335309 . Архивировано из оригинала 16 декабря 2012 г.
- ^ Бласкес; и др. (2001). «Цветение вовремя: гены, регулирующие цветочный переход» . Отчеты ЭМБО . 2 (12): 1078–1082. doi : 10.1093/embo-reports/kve254 . ПМЦ 1084172 . ПМИД 11743019 .
- ^ Фрелих, Майкл В.; Паркер, Дэвид С. (2000). «Преимущественно мужская теория эволюционного происхождения цветов: от генов к окаменелостям». Сист. Бот . 25 (2): 155–170. дои : 10.2307/2666635 . JSTOR 2666635 . S2CID 85612923 .
- ^ Ли А, Мао Л (2007). «Эволюция семейств генов микроРНК растений» . Клеточные исследования . 17 (3): 212–218. дои : 10.1038/sj.cr.7310113 . ПМИД 17130846 .
- ^ Добли Дж. Ф. (2004). «Генетика эволюции кукурузы» . Анну. Преподобный Жене . 38 (1): 37–59. дои : 10.1146/annurev.genet.38.072902.092425 . ПМИД 15568971 .
- ^ Пуруганан; и др. (2000). «Вариации и отбор цветочного гомеотического гена цветной капусты, сопровождающие эволюцию одомашненной капусты капустной» . Генетика . 155 (2): 855–862. дои : 10.1093/генетика/155.2.855 . ПМЦ 1461124 . ПМИД 10835404 .
- ^ Jump up to: а б Уильямс, Бен П.; Джонстон, Иэн Г.; Ковшофф, Сара; Хибберд, Джулиан М. (28 сентября 2013 г.). «Фенотипический ландшафтный вывод раскрывает множество эволюционных путей фотосинтеза C4» . электронная жизнь . 2 . Публикации eLife Sciences: e00961. дои : 10.7554/elife.00961 . ПМЦ 3786385 . ПМИД 24082995 . S2CID 6552541 .
- ^ Кристин, Пенсильвания; Осборн, CP; Шатле, Д.С.; Колумбус, Джей Ти; Беснар, Г.; Ходкинсон, Т.Р.; Гаррисон, штат Луизиана; Воронцова, М.С. ; Эдвардс, Э.Дж. (2012). «Анатомические факторы и эволюция фотосинтеза C 4 у трав» . Труды Национальной академии наук . 110 (4): 1381–1386. Бибкод : 2013PNAS..110.1381C . дои : 10.1073/pnas.1216777110 . ПМК 3557070 . ПМИД 23267116 .
- ^ Jump up to: а б с д Осборн, CP; Бирлинг, диджей (2006). «Обзор. Зеленая революция природы: замечательный эволюционный рост растений C 4 » . Философские труды Королевского общества Б. 361 (1465): 173–194. дои : 10.1098/rstb.2005.1737 . ПМК 1626541 . ПМИД 16553316 .
- ^ Jump up to: а б Реталлак, GJ (1 августа 1997 г.). «Неогеновое расширение североамериканских прерий». ПАЛЕОС . 12 (4): 380–390. Бибкод : 1997Палай..12..380R . дои : 10.2307/3515337 . JSTOR 3515337 .
- ^ Томассон-младший; Нельсон, Мэн; Закшевский, Р.Дж. (1986). «Ископаемая трава (Gramineae: Chloridoideae) из миоцена с анатомией Кранца». Наука . 233 (4766): 876–878. Бибкод : 1986Sci...233..876T . дои : 10.1126/science.233.4766.876 . ПМИД 17752216 . S2CID 41457488 .
- ^ О'Лири, Мэрион (май 1988 г.). «Изотопы углерода в фотосинтезе». Бионаука . 38 (5): 328–336. дои : 10.2307/1310735 . JSTOR 1310735 .
- ^ Осборн, П.; Фреклтон, П. (февраль 2009 г.). «Давление экологического отбора на фотосинтез C 4 в травах» . Труды: Биологические науки . 276 (1663): 1753–1760. дои : 10.1098/rspb.2008.1762 . ПМЦ 2674487 . ПМИД 19324795 .
- ^ Бонд, Вашингтон; Вудворд, Финляндия; Мидгли, Г.Ф. (2005). «Глобальное распространение экосистем в мире без огня». Новый фитолог . 165 (2): 525–538. дои : 10.1111/j.1469-8137.2004.01252.x . ПМИД 15720663 . S2CID 4954178 .
- ^ При содержании кислорода в атмосфере более 35% распространение огня невозможно остановить. Многие модели предсказывали более высокие значения, и их пришлось пересмотреть, поскольку полного вымирания растительной жизни не произошло.
- ^ Пагани, М.; Зачос, Джей Си; Фриман, К.Х.; Типпл, Б.; Богаты, С. (2005). «Заметное снижение концентрации углекислого газа в атмосфере в палеогене» . Наука . 309 (5734): 600–603. Бибкод : 2005Sci...309..600P . дои : 10.1126/science.1110063 . ПМИД 15961630 . S2CID 20277445 .
- ^ Пиперно, ДР; Сьюс, HD (2005). «Динозавры обедали на траве». Наука . 310 (5751): 1126–1128. дои : 10.1126/science.1121020 . ПМИД 16293745 . S2CID 83493897 .
- ^ Прасад, В.; Стрёмберг, CAE; Алимохаммадиан, Х.; Сахни, А. (2005). «Копролиты динозавров и ранняя эволюция трав и травоядных». Наука . 310 (5751): 1177–1180. Бибкод : 2005Sci...310.1177P . дои : 10.1126/science.1118806 . ПМИД 16293759 . S2CID 1816461 .
- ^ Кили, Дж. Э.; Рундел, PW (2005). «Пожар и расширение лугов C 4 в миоцене ». Экологические письма . 8 (7): 683–690. Бибкод : 2005EcolL...8..683K . дои : 10.1111/j.1461-0248.2005.00767.x .
- ^ Jump up to: а б Реталлак, Дж.Дж. (2001). «Кайнозойское расширение лугов и похолодание климата». Журнал геологии . 109 (4): 407–426. Бибкод : 2001JG....109..407R . дои : 10.1086/320791 . S2CID 15560105 .
- ^ Джин, JP; Он, К; Тан, X; Ли, З; Льв, Л; Чжао, Ю; Луо, Дж; Гао, Г. (июль 2015 г.). «Карта регуляции транскрипции арабидопсиса раскрывает различные функциональные и эволюционные особенности новых факторов транскрипции» . Молекулярная биология и эволюция . 32 (7): 1767–1773. дои : 10.1093/molbev/msv058 . ПМЦ 4476157 . ПМИД 25750178 .
- ^ Пичерский Е.; Банда Д. (2000). «Генетика и биохимия вторичных метаболитов растений: эволюционная перспектива». Тенденции растениеводства . 5 (10): 439–445. дои : 10.1016/S1360-1385(00)01741-6 . ПМИД 11044721 .
- ^ Нина Тайс; Мануэль Лердау (2003). «Эволюция функции вторичных метаболитов растений» . Межд. Дж. Наука о растениях . 164 (С3): С93–С102. дои : 10.1086/374190 . S2CID 85705162 .
- ^ Больманн Дж.; и др. (1998). «Терпеноидсинтазы растений: молекулярный и филогенетический анализ» . Учеб. Натл. акад. наук. США . 95 (8): 4126–4133. Бибкод : 1998PNAS...95.4126B . дои : 10.1073/pnas.95.8.4126 . ПМК 22453 . ПМИД 9539701 .
- ^ Хассани, М.А., Дуран, П. и Хаккард, С. (2018) «Микробные взаимодействия внутри холобионта растений». Микробиом , 6 (1): 58. два : 10.1186/s40168-018-0445-0 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|