Беннеттиталес
| Беннеттиталес | |
|---|---|

| |
| Реставрация представителя Williamsoniaceae Терезы Экблом. | |

| |
| Восстановление жизни "Williamsonia" sewardiana из раннего мела Индии, которая может представлять собой одного из ранних представителей Cycadeoidaceae. | |
| Научная классификация | |
| Королевство: | Растения |
| Клэйд : | Трахеофиты |
| Клэйд : | Сперматофиты |
| Заказ: | † Беннеттиталес Энглер, 1892 г. |
| Семьи | |
Bennettitales (также известные как цикадоиды ) — вымерший порядок семенных растений , впервые появившийся в пермском периоде и вымерший в большинстве районов к концу мелового периода . Bennettitales были одними из наиболее распространенных семенных растений мезозоя и имели морфологию, включая кустарниковые и саговниковые формы. Листва bennettitaleans внешне почти неотличима от листвы саговников, но они отличаются от саговников более сложными цветкообразными репродуктивными органами, по крайней мере некоторые из которых, вероятно, были опылены насекомыми . [ 1 ]
Хотя, конечно, голосеменные sensu lato (шишковидные семенные растения), взаимоотношения беннеттиталовых с другими семенными растениями обсуждаются. Их общее сходство с саговниками противоречит множеству более тонких особенностей их репродуктивной системы и строения листьев. Некоторые авторы связали bennettitaleans с покрытосеменными (цветковыми растениями) и гнетофитами (редкой и необычной группой современных голосеменных растений), образуя более широкую группу, известную как Anthophyta . Молекулярные данные противоречат этому: гнетофиты оказались гораздо более генетически похожими на хвойные деревья . Точная позиция Беннеттиталеса остается неопределенной.
Описание
[ редактировать ]

Bennettitales делятся на два семейства: Cycadeoidaceae и Williamsoniaceae , которые имеют разные особенности роста . Cycadeoidaceae имели толстые саговниковоподобные стволы с биспорангиатными (содержащими как мегаспоры , так и микроспоры ) стробилами (шишками), служащими их репродуктивными структурами. Williamsoniaceae имели либо двуспорангиатные, либо моноспорангиатные шишки, а также отчетливо тонкие и ветвящиеся деревянистые стволы. [ 1 ] Williamsoniaceae росли как древесные кустарники с разнородным ветвлением, похожим на таковое у Banksia . [ 2 ] Было высказано предположение, что Williamsoniaceae представляют собой парафилетическую (не содержащую всех потомков общего предка) совокупность всех Bennettitales, не принадлежащих к Cycadeoidaceae. [ 2 ]
Листва
[ редактировать ]Обычно листья беннеттитала прикрепляются к стеблю спиральным (штопорным) расположением. Некоторые листья (большинство видов Nilssoniopteris и др.) представляют собой узкие одиночные пластинки с гладким («целым») краем. [ 3 ] Большинство морфотипов листьев ( Pterophyllum , Ptilophyllum , Zamites , Otozamites и др.) перистые (перистые), с множеством мелких сегментов листа, прикрепленных к центральному стержню. Другие ( Anomozamites , некоторые виды Nilssoniopteris ) не полностью перистые (пилообразные) и являются переходными между этими двумя концевыми членами. Одна необычная форма листьев, Eoginkgoites , даже приближается к пальчатой форме, похожей на ранние виды гинкго . [ 4 ]
Листва bennettitaleans до такой степени напоминает листву саговников, что листву этих двух групп невозможно надежно различить только на основе общей морфологии. Однако ископаемая листва, сохранившая кутикулу, может быть с уверенностью отнесена к любой группе. Устьица . беннеттиталовых описываются синдетохейльные как Это означает, что основные парные замыкающие клетки развиваются из тех же материнских клеток, что и окружающие их вспомогательные клетки. Это контрастирует с гаплохейлическими устьицами саговников и хвойных деревьев. В гаплохейлических устьицах кольцо вспомогательных клеток не происходит из тех же исходных структур, что и замыкающие клетки. Это фундаментальное различие является основным способом различения листвы беннеттитовой и саговниковой. [ 5 ]
Шишки и семена
[ редактировать ]

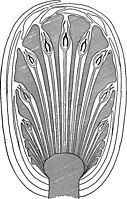
Как и у других голосеменных растений, у беннеттитальных репродуктивные соцветия имеют форму шишек , образующих пыльцу и семяпочки (неоплодотворенные семена). Шишки имеют толстый центральный сосуд, окруженный простыми, спирально расположенными плодородными и неплодородными структурами. Ткань у основания конуса образует слои чешуйчатых или лепесткообразных прицветников, защищающих излучающие внутренние структуры. Некоторые авторы называют беннеттитальные шишки «цветами», хотя они не эквивалентны настоящим цветкам покрытосеменных растений . Пыльца часто заключена в парные синангии (пыльцевые мешочки). Синангии лежат на адаксиальном (внутреннем) крае листообразных структур, несущих пыльцу, известных как микроспорофиллы . Это контрастирует с саговниками, у всех из которых отсутствуют дискретные синангии и они несут пыльцу на абаксиальной (внешней) поверхности микроспорофиллов. [ 6 ]
Многие bennettitaleans являются двуспорангиатными , при этом пыльца и семязачатки располагаются на одной (двуполой или гермафродитной) шишке. Полости, заполненные изогнутыми микроспорофиллами, несущими синангии, окружены тонкими радиальными структурами, включающими толстые неплодородные межсеменные чешуйки и фертильные спорофиллы с семяпочками на концах. Наличие семязачатков на кончиках спорофиллов, а не на кончиках стеблей, является основным отличием шишек беннеттиталовых от гнетофитов. По мере оплодотворения и созревания шишки микроспорофиллы отмирают, а семязачатки превращаются в семена. [ 6 ]
Большинство bennettitaleans семейства Williamsoniaceae вместо этого являются моноспорангиатными , с отдельной пыльцой и образующими семяпочки (однополыми) шишками на одном растении. Шишки, дающие семяпочки (женские) ( Williamsonia и др . ), подобны зрелым двуспорангиатным шишкам, с межсемянными чешуйками и спорофиллами с семязачатками, окруженными прицветниками. Пыльцевые (мужские) шишки ( Weltrichia и др . ), напротив, имеют обнаженную крону из суженных микроспорофиллов с адаксиальными рядами синангиев. Микроспорофиллы могут содержать один линейный ряд парных синангиев или вместо этого синангии, расположенные в виде перистых (перьевых) узоров. [ 6 ]
Семена двудольные (имеют два зародышевых листа ), с центральным зародышем, окруженным тремя слоями: тонким мегагаметофитом, несколько более толстым нуцеллусом и защитным покровом . Верхний кончик семени сужается и открывается тонким и часто расширенным микропиле . Длинное узкое микропиле, выходящее из семени, внешне похоже на состояние живых гнетофитов . После оплодотворения семени микропиле закрывается пробкообразным продолжением нуцеллуса. В отличие от живых голосеменных растений, на кончике нуцеллуса нет пыльцевой камеры (приемника для накопленной пыльцы). Покровы плотные и толстые, со многими слоями дифференцированных клеток. Это контрастирует с тонким двурядным (двухслойным) покровом гнетофитов. У беннеттиталеев также отсутствует еще одна черта, напоминающая гнетофит: оболочка из сросшихся прицветников, окутывающих семя. Большинство покровных клеток не являются необычными по размеру и форме. Однако вблизи микропиле самый внутренний слой интегументных клеток становится радиально ориентированным и удлиненным, частично замыкаясь на микропиле. Нуцеллус и интегумент над поверхностью тела не срослись. халаза (основание семени), в отличие от саговников или гнетофитов, у которых слои на большей части своей высоты слиты. [ 6 ]
Было высказано предположение, что цикадоидные были самоопыляемыми, их стебли и шишки были погребены под землей. [ 1 ] [ 7 ] хотя альтернативно предполагалось, что они были опылены жуками. [ 8 ] Цветкоподобная мужская репродуктивная структура вильямсонии Weltrichia связана с женской репродуктивной структурой Williamsonia , хотя неясно, были ли родительские растения однодомными (мужские и женские репродуктивные структуры присутствовали на одном и том же растении) или двудомными (где каждое растение имеет только одну пол репродуктивного органа). Вельтрихия , вероятно, в основном опылялась ветром , а некоторые виды, возможно, опылялись жуками. [ 9 ]
Несколько групп юрских и раннемеловых насекомых обладали длинным хоботком , и было высказано предположение, что они питались нектаром, вырабатываемым беннеттитовыми репродуктивными структурами, такими как двуполая репродуктивная структура вильямсониевых Williamsoniella , которая имела длинный и узкий центральный сосуд, который, вероятно, в противном случае недоступен. [ 10 ] Было обнаружено, что пыльца bennettitalean раннего мела напрямую ассоциировалась с мухой-носителем хоботка, принадлежащей вымершему семейству Zhangsolvidae , что свидетельствует о том, что это семейство действовало в качестве опылителей для этой группы. [ 11 ] Межсемянные чешуи овулирующих шишек Bennettitales могли стать мясистыми по мере созревания, что потенциально могло сделать их привлекательными для диких животных, служивших распространителями семян. [ 12 ]
Таксономия
[ редактировать ]История открытия
[ редактировать ]Cycadeoideae (первоначально «Cycadeoideae») были названы английским геологом Уильямом Баклендом в 1828 году из ископаемых стволов, найденных в юрских слоях на острове Портленд в Англии, которым Бакленд дал рода название Cycadeoidea . Бакленд предоставил описание семейства и двух видов, но не дал описания рода, в результате чего описание семейства Баклендом было признано недействительным по современным таксономическим стандартам. [ 13 ] В публикациях 1870 года шотландский ботаник Уильям Каррутерс и английский палеоботаник Уильям Кроуфорд Уильямсон описали первые известные репродуктивные органы Bennettitales из юрских отложений Йоркшира и юрско-меловых отложений острова Уайт и острова Портленд. [ 14 ] [ 15 ] [ 16 ] Карутерс был первым, кто осознал, что Bennettitales имеет явные отличия от саговников, и основал трибы «Williamsonieae» и «Bennettiteae». [ 16 ] причем последний назван в честь рода Bennettites, названного Карутерсом в той же публикации , в честь британского ботаника Джона Джозефа Беннета . [ 14 ] [ 17 ] Отряд Bennettitales был создан немецким ботаником Адольфом Энглером в 1892 году, который признал эту группу отдельной от Cycadales. [ 18 ]
Отношения с другими семенными растениями
[ редактировать ]Гипотеза антофитов , выдвинутая Арбером и Паркингом в 1907 году. [ 19 ] предположил, что покрытосеменные произошли от Bennettitales, о чем свидетельствуют древоподобные структуры и рудиментарные цветки . [ 1 ] Однако на основании морфологических данных Bennettitales были классифицированы как монофилетическая группа в сочетании с Gnetales. [ 20 ] исследование 2006 года показало, что Bennettitales, покрытосеменные и Gigantopteridales образуют кладу , основанную на присутствии олеанана . [ 21 ] Молекулярные данные постоянно противоречат гипотезе антофитов, обнаруживая, что покрытосеменные являются сестринской группой всех ныне живущих голосеменных, включая Gnetales. [ 22 ] Некоторые авторы предположили, что из-за сходства семенных оболочек Bennettitales образуют кладу с отрядами голосеменных Gnetales и Erdtmanithecales , получившую название «группа BEG». [ 23 ] Однако это предложение оспаривается другими авторами, которые утверждают, что эти сходства лишь поверхностны и не указывают на близкое родство. [ 24 ] Филогения 2017 года, основанная на молекулярных признаках окаменелой кутикулы, показала, что Bennettitales были более тесно связаны с кладой Ginkgo + Cycads, чем с хвойными деревьями, и были тесно связаны с Nilssonia и Ptilozamites . [ 25 ]
Эволюционная история
[ редактировать ]Самыми старыми подтвержденными окаменелостями bennettitaleans являются листья Nilssoniopteris shanxiensis , вида из верхней части формации Верхнего Шиххоцзе в провинции Шаньси , Китай . [ 5 ] Эти толщи датируются раннекунгурским этапом ранней перми ( Приуралья ), около 281 миллиона лет назад. [ 26 ] Предполагаемые каменноугольно-пермские записи Pterophyllum не имеют убедительного сходства с беннеттиталами или были переинтерпретированы как листва саговников в форме рода Pseudoctenis . [ 27 ] Настоящие пермские находки листьев бенеттиталовой редки; за пределами формации Шиххоцзе они встречаются только в позднепермского (вероятно, чангсинского периода ) возраста формации Умм Ирна в Иордании . [ 5 ] Это образование примечательно ранним появлением другой флоры мезозойского типа, включая самые ранние записи користоспермальной листвы ( Dicroidium ). [ 28 ] Отряд Fredlindiales (содержащий род Fredlindia ) из позднего триаса Гондваны, по-видимому, тесно связан с Bennettitales, но отличается от него в некоторых аспектах своих репродуктивных органов. [ 16 ]
Летопись окаменелостей беннеттиталов вновь появилась в среднем триасе , и к концу периода williamsoniaceans стали распространяться по всему миру. [ 29 ] [ 5 ] Самые старые репродуктивные структуры беннеттиталов - это маленькие «цветки» Уильямсонии из формации Эск среднего триаса в Австралии . [ 16 ] В то время как Williamsoniaceae имели глобальное распространение, Cycadeoidaceae, по-видимому, были в основном ограничены западными частями Лавразии и в основном известны с мелового периода. [ 16 ] Bennettitales были широко распространены и многочисленны в юрском и раннем меловом периоде, однако численность беннеттиталов резко сократилась в позднем меловом периоде, что совпало с появлением цветковых растений , которые в основном вымерли к концу периода, и были обнаружены последние известные останки из Северного полушария. в полярной широте Формация Каканаут на Чукотке , Россия, датируемая маастрихтом , относящаяся к Pterophyllum. [ 30 ] Сообщалось о возможной поздней находке из раннего олигоцена восточной Австралии и Тасмании , относящейся к роду Ptilophyllum , но кутикула не сохранилась, что делает отнесение неубедительным. [ 31 ]
Подгруппы
[ редактировать ]- Cycadolepis (чешуйки или прицветники, не относящиеся к семейству)
- Гаитянский ? (орган пыльцы?)
- Лунция (пыльцевой орган, нерасположенный в семье)
- Легуминантус ? (орган пыльцы?)
- Лейтардтия ? (орган пыльцы?)
- Вестершеймия (овуляционный орган, нерасположенный в семье)
- Антрофиопсис ? [ 32 ] (лист)
- Семейство Цикадоидные.
- Семейство Вильямсониевые.
- Аномозамиты (лист)
- Bennetticarpus (женская семенная шишка)
- Беннеттистемон (мужской пыльцевой орган)
- Бакландия (топоры)
- Эогинггоиты (лист)
- Ишнофитон
- Кимуриелла (целое растение)
- Нильссониоптерис (лист)
- Отозамиты (лист)
- Птерофиллум (лист)
- Птилофиллум (лист)
- Вардеклоефтия (женская семенная шишка)
- Вельтрихия (мужской пыльцевой орган)
- Виландиелла (целое растение)
- Вильямсония (женские семенные шишки)
- Williamsoniella (двуполая репродуктивная структура)
- Замиты (лист, частично)
- Гаитингерия (орган пыльцы)
Bennettitales обычно считается единственным отрядом в классе Bennettitopsida Engler (1897) или Cycadeoideopsida Scott (1923) . Большинство палеоботаников предпочитают использовать эти два семейства, хотя некоторые авторы, такие как Андерсон и Андерсон (2007), классифицируют отряд по большему числу семейств. [ 33 ] Anderson & Anderson также классифицировали отряды Fredlindiales Anderson & Anderson (2003). [ 34 ] и Pentoxylales Pilger & Melchior (1954) в пределах Bennettitopsida. [ 33 ]
Галерея
[ редактировать ]-
 Ископаемый лист Zamites mariposana из юрского периода .
Ископаемый лист Zamites mariposana из юрского периода . -
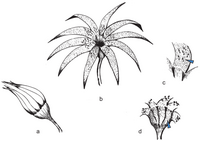 Восстановление Weltrichia magna из юры Мексики.
Восстановление Weltrichia magna из юры Мексики. -
 отозамита Лист
отозамита Лист -
 Лист аномозамитов
Лист аномозамитов -
-
 Ствол цикадоидеи
Ствол цикадоидеи -
 Ископаемое Ptilophyllum grandifolium
Ископаемое Ptilophyllum grandifolium







Ссылки
[ редактировать ]- ^ Jump up to: а б с д Пеньяльвер, Энрике; Арилло, Антонио; Перес-де ла Сурс, Рикардо; Риччио, Марк Л.; Дельклос, Ксавье; Бэррон, Эдвард; Гримальди, Дэвид А. (2015). «Длиннохоботные мухи как опылители меловых голосеменных растений» . Современная биология . 25 (14): 1917–1923. дои : 10.1016/j.cub.2015.05.062 . ПМИД 26166781 . S2CID 13022302 .
- ^ Jump up to: а б Потт, Кристиан; Маклафлин, Стивен (01 июня 2014 г.). «Различный характер роста Williamsoniaceae (Bennettitales): раскрытие экологии ключевой группы мезозойских растений» . Палеобиоразнообразие и палеосреда . 94 (2): 307–325. дои : 10.1007/s12549-014-0157-9 . S2CID 84440045 .
- ^ Рэй, ММ; Ротвелл, GW; Стоки, РА (сентябрь 2014 г.). «Анатомически сохранившиеся раннемеловые беннеттитальные листья: Nilssoniopteris corrugata n. sp. с острова Ванкувер, Канада» . Журнал палеонтологии . 88 (5): 1085–93. дои : 10.1017/S002233600005767X . S2CID 232349931 .
- ^ Потт, Кристиан; Аксмит, Брайан Дж. (февраль 2015 г.). «Williamsonia carolinensis sp. nov. и связанная с ней листва Eoginkgoites из верхнетриасовой формации Пекин, Северная Каролина: значение для ранней эволюции Williamsoniaceae (Bennettitales)» . Международный журнал наук о растениях . 176 (2): 174–185. дои : 10.1086/679471 . S2CID 44559347 .
- ^ Jump up to: а б с д Бломенкемпер, Патрик; Боймер, Роберт; Бэкер, Мальте; Абу Хамад, Абдалла; Ван, Цзюнь; Керп, Ганс; Бомфлер, Бенджамин (2021). «Беннеттитальные листья из перми экваториальной Пангеи — раннее излучение культовой мезозойской группы голосеменных» . Границы в науках о Земле . 9 : 162. Бибкод : 2021FrEaS...9..162B . дои : 10.3389/feart.2021.652699 .
- ^ Jump up to: а б с д Ротвелл, Гар В.; Крепет, Уильям Л.; Стоки, Рут А. (2009). «Жива ли гипотеза антофитов? Новые данные о репродуктивных структурах Bennettitales» . Американский журнал ботаники . 96 (1): 296–322. дои : 10.3732/ajb.0800209 . ПМИД 21628190 .
- ^ Осборн, Дж. М.; Тейлор, МЛ (2010). «Структура пыльцы и копролита Cycadeoidea (Bennettitales): значение для понимания систем опыления и спаривания у мезозойских цикадоидов». Растения в глубоком мезозое: морфологические инновации, филогения и экосистемы . Блумингтон, Индиана: Издательство Университета Индианы. стр. 34–49. ISBN 978-0-253-00199-3 .
- ^ Рамка, Рассвет; Готтсбергер, Герхард (21 апреля 2023 г.). «Различные сексуальные стратегии ископаемых голосеменных растений: новый взгляд на опыление Bennettitales» . Фитон . 62/63.
- ^ Попа, Михай Эмилиан (27 февраля 2019 г.). «Обзор беннеттитального рода Weltrichia» . Журнал палеогеографии . 8 (1): 12. Бибкод : 2019JPalg...8...12P . дои : 10.1186/s42501-019-0023-9 . S2CID 85458053 .
- ^ Храмов, Александр В.; Лукашевич, Елена Дмитриевна (июль 2019 г.). «Юрский двукрылый опылитель с чрезвычайно длинным хоботком». Исследования Гондваны . 71 : 210–215. Бибкод : 2019GondR..71..210K . дои : 10.1016/j.gr.2019.02.004 . S2CID 134847380 .
- ^ Пеньяльвер Э, Арилло А, Перес-де ла Фуэнте Р, Риччио МЛ, Дельклос Х, Баррон Э, Гримальди Д.А. (июль 2015 г.). «Длиннохоботные мухи как опылители меловых голосеменных растений» . Современная биология . 25 (14): 1917–23. дои : 10.1016/j.cub.2015.05.062 . ПМИД 26166781 .
- ^ Фриис, Эльза Мари; Педерсен, Кай Раунсгаард; Крейн, Питер Р., ред. (2011), «Покрытосеменные в контексте: современные и ископаемые семенные растения» , Ранние цветы и эволюция покрытосеменных , Кембридж: Cambridge University Press, стр. 101–140, ISBN 978-0-521-59283-3 , получено 20 марта 2023 г.
- ^ Доуэлд, Александр Б. (3 мая 2016 г.). «Номенклатура Cycadeoidea (ископаемые Spermatophyta: Cycadeoideopsida)» . Таксон . 65 (2): 372–379. дои : 10.12705/652.16 .
- ^ Jump up to: а б Каррутерс, Уильям (май 1870 г.). «XVIII. Об ископаемых цикадовых стеблях из вторичных пород Британии» . Труды Лондонского Линнеевского общества . 26 (4): 675–708. дои : 10.1111/j.1096-3642.1870.tb00201.x .
- ^ Уильямсон, WC (май 1870 г.). «XVII. Вклад в историю Замиа Гигас, Линдл и Хатт. *» . Труды Лондонского Линнеевского общества . 26 (4): 663–674. дои : 10.1111/j.1096-3642.1870.tb00200.x .
- ^ Jump up to: а б с д и Маклафлин, Стивен; Потт, Кристиан; Соббе, Ян Х. (01 марта 2018 г.). «Разнообразие репродуктивных органов австралийских мезозойских беннеттитопсидов» (PDF) . Палеобиоразнообразие и палеосреда . 98 (1): 71–95. дои : 10.1007/s12549-017-0286-z . S2CID 135237376 .
- ^ Бриттен, Джеймс; Боулджер, Джордж Саймондс (1893). Биографический указатель британских и ирландских ботаников . Лондон: Вест, Ньюман и компания. п. 14. дои : 10.5962/bhl.title.56947 .
- ^ Энглер, HGA 1892. Программа лекций по специальной и медико-фармацевтической ботанике . xxiii + 184 стр. Gebr.
- ^ Арбер, EN; Паркин, Дж. (июль 1907 г.). «О происхождении покрытосеменных растений» . Ботанический журнал Линнеевского общества . 38 (263): 29–80. дои : 10.1111/j.1095-8339.1907.tb01074.x .
- ^ Крейн, PR (сентябрь 1985 г.). «Филогенетические взаимоотношения семенных растений». Кладистика . 1 (4): 329–348. дои : 10.1111/j.1096-0031.1985.tb00432.x . ПМИД 34965681 . S2CID 85709030 .
- ^ Тейлор, Дэвид Уиншип; Ли, Хунци; Даль, Джереми; Фаго, Фред Дж.; Зинникер, Дэвид; Молдован, Дж. Майкл (март 2006 г.). «Биогеохимические доказательства присутствия молекулярного ископаемого олеанана покрытосеменных растений в палеозойских и мезозойских не покрытосеменных окаменелостях». Палеобиология . 32 (2): 179–90. doi : 10.1666/0094-8373(2006)32[179:BEFTPO]2.0.CO;2 . S2CID 83801635 .
- ^ Ран, Цзинь-Хуа; Шен, Тин-Тин; Ван, Мин-Мин; Ван, Сяо-Цюань (июнь 2018 г.). «Филогеномика решает глубокую филогению семенных растений и указывает на частичную конвергентную или гомопластическую эволюцию между Gnetales и покрытосеменными» . Слушания. Биологические науки . 285 (1881): 20181012. doi : 10.1098/rspb.2018.1012 . ПМК 6030518 . ПМИД 29925623 .
- ^ Фрис, Эльза Мари; Крейн, Питер Р.; Педерсен, Каге Раунсгаард; Бенгтсон, Стивен; Донохью, Филип Си Джей; Гримм, Гвидо В.; Стампанони, Марко (ноябрь 2007 г.). «Фазово-контрастная рентгеновская микротомография связывает семена мелового периода с Gnetales и Bennettitales» . Природа 450 (7169): 549–52. Бибкод : 2007Nature.450..549F . дои : 10.1038/nature06278 . ПМИД 18033296 . S2CID 1198220 .
- ^ Ротвелл, Гар В.; Стоки, Рут А. (март 2013 г.). «Эволюция и филогения гнетофитов: данные по анатомически сохранившимся семенным конусам Protoephedrites eamesii gen. et sp. nov. и семенам нескольких видов Bennettitalean» . Международный журнал наук о растениях . 174 (3): 511–529. дои : 10.1086/668688 . ISSN 1058-5893 . S2CID 84063572 .
- ^ Вайда, Виви; Пучетайте, Милда; Маклафлин, Стивен; Энгдаль, Андерс; Хеймдал, Джимми; Увдал, Пер (август 2017 г.). «Молекулярные признаки ископаемых листьев предоставляют неожиданные новые доказательства взаимоотношений вымерших растений» . Экология и эволюция природы . 1 (8): 1093–1099. дои : 10.1038/s41559-017-0224-5 . ПМИД 29046567 . S2CID 3604369 .
- ^ Ву, Цюн; Рамезани, Джахандар; Чжан, Хуа; Ван, Цзюнь; Цзэн, Фангуй; Чжан, Ичунь; Лю, Фэн; Чен, Цзюнь; Цай, Яофэн; Хоу, Чжаншуай; Лю, Чао (05 февраля 2021 г.). «Высокоточные U-Pb возрастные ограничения пермского круговорота флоры, изменения палеоклимата и тектоники Северо-Китайского блока» . Геология . 49 (6): 677–681. Бибкод : 2021Geo....49..677W . дои : 10.1130/G48051.1 . S2CID 234064909 .
- ^ Потт, Кристиан; Маклафлин, Стивен; Линдстрем, Анна (2009). «Позднепалеозойская листва из Китая демонстрирует сходство с Cycadales, а не с Bennettitales, что требует переоценки палеозойских видов Pterophyllum» . Acta Palaeontologica Polonica . 55 (1): 157–168. дои : 10.4202/app.2009.0070 . S2CID 52253114 .
- ^ Бломенкемпер, Патрик; Керп, Ганс; Абу Хамад, Абдалла; ДиМишель, Уильям А.; Бомфлер, Бенджамин (декабрь 2018 г.). «Скрытая колыбель эволюции растений в пермских тропических низменностях» . Наука . 362 (6421): 1414–1416. Бибкод : 2018Sci...362.1414B . дои : 10.1126/science.aau4061 . ПМИД 30573628 .
- ^ Кустачер, Эвелин; ван Конийненбург-ван Циттерт, Йоханна Х.А. (2005). «Ладинская флора (средний триас) Доломитовых Альп: палеоэкологические реконструкции и палеоклиматические соображения» (PDF) . Гео.Альп . 2 : 31–51.
- ^ Гниловская Анастасия Александровна; Головнёва Лина Борисовна (февраль 2018 г.). «Позднемеловой Pterophyllum (Bennettitales) на Северо-Востоке России» . Меловые исследования . 82 : 56–63. дои : 10.1016/j.cretres.2017.09.013 .
- ^ Маклафлин, Стивен; Карпентер, Раймонд Дж.; Потт, Кристиан (апрель 2011 г.). «Ptilophyllum muelleri (Ettingsh.) comb. nov. из олигоцена Австралии: последний из Bennettitales?» . Международный журнал наук о растениях . 172 (4): 574–85. дои : 10.1086/658920 . S2CID 52885618 .
- ^ Сюй, Юаньюань; Ван, Юндон; Маклафлин, Стивен (сентябрь 2023 г.). «Насколько схожи жилкование и кутикулярные признаки Glossopteris, Sagenopteris и Anthrophyopsis?» . Обзор палеоботаники и палинологии . 316 : 104934. doi : 10.1016/j.revpalbo.2023.104934 .
- ^ Jump up to: а б Андерсон, Джон М.; Андерсон, Хайди М.; Клил, Крис Дж. (2007). «Краткая история голосеменных растений: классификация, биоразнообразие, фитогеография и экология» (PDF) . Стрелиция . 20 : 1–280.
- ^ Андерсон, Джон М.; Андерсон, Хайди М. (2003). «Расцвет голосеменных растений: систематика и биоразнообразие позднетриасовых молтенских фруктификаций» . Стрелиция . 15 : 1–308.