Область крепления каркаса/матрицы
Термин S/MAR ( область прикрепления каркаса/матрикса ), иначе называемый SAR ( область прикрепления каркаса ) или MAR ( область, связанная с матриксом ), представляет собой последовательности в ДНК эукариотических хромосом, к которым прикрепляется ядерный матрикс . Являясь архитектурными компонентами ДНК, которые организуют геном эукариот в функциональные единицы внутри ядра клетки , S/MAR опосредуют структурную организацию хроматина внутри ядра. Эти элементы составляют опорные точки ДНК для каркаса хроматина и служат для организации хроматина в структурные домены . Исследования отдельных генов привели к выводу, что динамическая и сложная организация хроматина, опосредованная элементами S/MAR, играет важную роль в регуляции экспрессии генов .
Обзор
[ редактировать ]Уже много лет известно, что полимерная сеть, так называемая « ядерная матрица » или «ядерный каркас», является важным компонентом эукариотических ядер. Этот ядерный скелет действует как динамическая опора для многих специализированных событий, связанных со считыванием и распространением генетической информации (см. ниже).
S/MARs сопоставляются с неслучайными участками генома. Они встречаются на флангах транскрибируемых областей, в 5'- интронах , а также в кластерных областях генных точек разрыва (BCR). Будучи точками ассоциации для общих ядерных структурных белков, S/MAR необходимы для аутентичной и эффективной хромосомной репликации и транскрипции , для рекомбинации и конденсации хромосом. S/MAR не имеют очевидной консенсусной последовательности. Хотя элементы-прототипы состоят из богатых АТ участков длиной в несколько сотен пар оснований, общий состав оснований определенно не является основным фактором, определяющим их активность. Вместо этого их функция требует наличия структуры «AT-патчей», которые придают склонность к локальному распариванию цепей при скручивающей нагрузке .
Подходы биоинформатики подтверждают идею о том, что благодаря этим свойствам S/MARs не только отделяют данную транскрипционную единицу (домен хроматина) от ее соседей, но также предоставляют платформы для сборки факторов, обеспечивающих транскрипционные события внутри данного домена. Повышенная склонность к разделению нитей ДНК (так называемый потенциал «стресс-индуцированной дуплексной дестабилизации», SIDD ) может служить образованию вторичных структур, таких как крестоформы или структуры проскальзывания , которые являются узнаваемыми признаками для ряда ферментов ( ДНКазы , топоизомеразы) . , поли(АДФ-рибозил)полимеразы и ферменты аппарата ацетилирования гистонов и метилирования ДНК ). S/MAR были классифицированы как конститутивные (действующие как постоянные границы доменов во всех типах клеток) или факультативные (связанные с типом клеток и активностью) в зависимости от их динамических свойств.
Хотя число S/MAR в геноме человека , по оценкам, приближается к 64 000 (домены хроматина) плюс еще 10 000 (фокусы репликации), в 2007 году лишь незначительная часть (559 для всех эукариот) соответствовала стандартным критериям аннотация в базе данных S/MARt. [ 1 ]
Контекстно-зависимые свойства
[ редактировать ]Современные взгляды на ядерный матрикс рассматривают его как динамическую сущность, которая меняет свои свойства в соответствии с требованиями клеточного ядра — почти так же, как цитоскелет адаптирует свою структуру и функцию к внешним сигналам. Оглядываясь назад, следует отметить, что открытие S/MAR имеет два основных пути:
- описание элементов прикрепления каркаса (SAR) Леммли и его коллег, которые, как считалось, разграничивают границы данного домена хроматина. [ 2 ]
- характеристика матрикс-ассоциированных областей (MAR), первые примеры которых поддерживали энхансер капп-цепи иммуноглобулина в зависимости от его занятости факторами транскрипции [ 3 ]
Последующая работа продемонстрировала как конститутивную (подобную SAR), так и факультативную (подобную MAR) функцию элементов в зависимости от контекста. В то время как было обнаружено, что конститутивные S/MAR связаны с сайтом гиперчувствительности к ДНКазе I во «всех» типах клеток (независимо от того, был ли транскрибирован включенный домен), гиперчувствительность ДНКазы I факультативного типа зависела от транскрипционного статуса. [ 4 ] Основное различие между этими двумя функциональными типами S/MAR заключается в их размере: конститутивные элементы могут простираться на несколько тысяч пар оснований, тогда как факультативные элементы имеют нижний предел размера около 300 пар оснований.
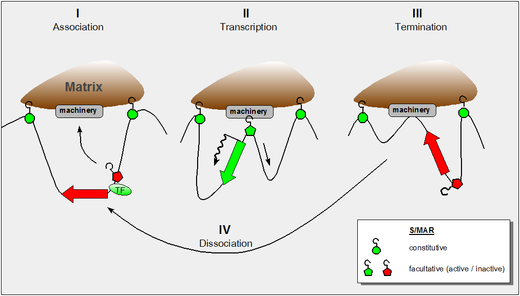
На рисунке показано наше нынешнее понимание этих свойств, и он включает в себя следующие выводы:
- динамические свойства контактов S/MAR-каркас, полученные в результате haloFISH исследований [ 5 ]
- тот факт, что во время транскрипции ДНК наматывается через РНК-полимеразу, которая сама по себе является фиксированным компонентом ядерного матрикса. [ 6 ]
- тот факт, что некоторым внутренним S/MAR домена требуется поддержка соседнего транскрипционного фактора, чтобы стать активными. [ 4 ]
Использование в генной терапии
[ редактировать ]В качестве альтернативы вирусным векторам, которые могут оказывать нежелательное воздействие на организм пациентов, изучаются невирусные методы генной терапии. Один из таких методов использует плазмиды с особыми свойствами — так называемые эписомы . Эписомы обладают способностью делиться вместе с остальной частью эукариотического генома во время митоза. По сравнению со стандартными плазмидами они не подавляются эпигенетически внутри ядра и не разрушаются ферментативно. Эписомы приобретают эту способность благодаря наличию последовательности S/MAR в своей конструкции. [ 7 ]
Дополнительная информация
[ редактировать ]В 2006 году Тетко обнаружил сильную корреляцию внутригенных S/MAR с пространственно-временной экспрессией генов у Arabidopsis thaliana . [ 8 ] В масштабе генома обнаружены выраженные ткане- и органоспецифичные и онтогенетические закономерности экспрессии S/MAR-содержащих генов. Примечательно, что гены транскрипционных факторов содержат значительно более высокую долю S/MAR. Выраженное различие в характеристиках экспрессии S/MAR-содержащих генов подчеркивает их функциональную значимость и важность структурных характеристик хромосом для регуляции генов как у растений, так и у других эукариот.
Ссылки
[ редактировать ]- ^ «СМАРТДБ» . Кафедра биоинформатики . Геттинген, Германия: Геттингенский университет.
- ^ Миркович Дж., Миро М.Э., Леммли, Великобритания (ноябрь 1984 г.). «Организация петли хроматина высшего порядка: специфические места прикрепления ДНК на ядерном каркасе». Клетка . 39 (1): 223–232. дои : 10.1016/0092-8674(84)90208-3 . ПМИД 6091913 . S2CID 24634819 .
- ^ Кокерилл П.Н., Garrard WT (январь 1986 г.). «Закрепление хромосомной петли гена каппа-иммуноглобулина происходит рядом с энхансером в области, содержащей сайты топоизомеразы II». Клетка . 44 (2): 273–282. дои : 10.1016/0092-8674(86)90761-0 . ПМИД 3002631 . S2CID 26202934 .
- ^ Jump up to: а б Клар М., Стелламаннс Э., Ак П., Глюх А., Боде Дж. (декабрь 2005 г.). «Доминантные геномные структуры: обнаружение и потенциальные сигнальные функции в домене бета-интерферона». Джин . 364 : 79–89. дои : 10.1016/j.gene.2005.07.023 . ПМИД 16185826 .
- ^ Хэн Х.Х., Гетце С., Йе С.Дж., Лю Г., Стивенс Дж.Б., Бремер С.В. и др. (март 2004 г.). «Петли хроматина избирательно закрепляются с помощью областей прикрепления каркаса / матрицы». Журнал клеточной науки . 117 (Часть 7): 999–1008. дои : 10.1242/jcs.00976 . ПМИД 14996931 .
- ^ Джексон Д.А., Долле А., Робертсон Дж., Кук PR (август 1992 г.). «Прикрепление петель хроматина к нуклеоскелету». Международные отчеты по клеточной биологии . 16 (8): 687–696. дои : 10.1016/s0309-1651(05)80013-x . ПМИД 1446346 .
- ^ Мулиа Дж.Е., Пикансо-Кастро В., Ставру Э.Ф., Атанассиаду А., Фигейредо М.Л. (октябрь 2021 г.). «Достижения в разработке и применении невирусных эписомальных векторов для генной терапии» . Генная терапия человека . 32 (19–20): 1076–1095. дои : 10.1089/hum.2020.310 . ПМЦ 8819515 . ПМИД 34348480 .
- ^ Тетко И.В., Хаберер Г., Радд С., Мейерс Б., Мьюз Х.В., Майер К.Ф. (март 2006 г.). «Пространственно-временной контроль экспрессии коррелирует с областями прикрепления внутригенного каркасного матрикса (S/MAR) у Arabidopsis thaliana» . PLOS Вычислительная биология . 2 (3): е21. дои : 10.1371/journal.pcbi.0020021 . ПМЦ 1420657 . ПМИД 16604187 .