Предвзятость при введении вариаций
Предвзятость при введении вариаций («систематическая ошибка прибытия») — это теория в области эволюционной биологии , которая утверждает, что предвзятость при введении наследственной изменчивости отражается на результатах эволюции . Это имеет отношение к темам молекулярной эволюции , эво-дево и самоорганизации . [1] [2] В контексте этой теории «интродукция» («происхождение») — это технический термин, обозначающий события, которые сдвигают частоту аллеля вверх от нуля ( мутация — это генетический процесс, который преобразует одну аллель в другую, тогда как интродукция — это популяционный генетический процесс, который добавляет к набору аллелей в популяции с ненулевыми частотами). Формальные модели демонстрируют, что, когда эволюционный процесс зависит от событий интродукции, мутационные и эволюционные отклонения в генерации вариаций могут влиять на ход эволюции по принципу «первым пришел — первым обслужен», так что эволюция отражает появление более вероятного, а не только более вероятного. выживание слесаря. [3] [4] [5] [6] [7] В то время как обычно предполагается, что мутационные объяснения эволюционных моделей подразумевают или требуют нейтральной эволюции , теория предвзятости прибытия однозначно предсказывает возможность адаптации, основанной на мутациях . [8] Прямые доказательства этой теории получены в лабораторных исследованиях, показывающих, что адаптивные изменения систематически обогащаются мутационно-вероятными типами изменений. [9] [10] [11] [12] Ретроспективный анализ естественных случаев адаптации также подтверждает эту теорию. [11] [13] Эта теория примечательна как пример современного структуралистского мышления, контрастирующего с классической функционалистской точкой зрения, согласно которой ход эволюции определяется естественным отбором (см. [14] ).
История [ править ]
Теория предубеждений в процессе внедрения как причина ориентации или направления эволюции объясняется сближением двух нитей. [15] Первым из теоретической популяционной генетики является явное признание теоретиками (к концу 20-го века) того, что правильная трактовка эволюционной динамики требует зависящего от скорости процесса внедрения (происхождения), отсутствующего в классических трактовках эволюции как процесса. сдвига частот доступных аллелей. [16] [17] [18] [19] Это признание проявляется в появлении моделей происхождения-фиксации , которые изображают эволюцию как двухэтапный процесс возникновения и фиксации (путем дрейфа или отбора ), скорость которого определяется умножением скорости внедрения (основанной на скорости мутации ) на вероятность фиксации (на основе эффекта приспособленности ). Модели фиксации происхождения [19] появились в разгар молекулярной революции , спустя полвека после зарождения теоретической популяционной генетики: вскоре они стали широко применяться в нейтральных моделях скорости и закономерностей молекулярной эволюции ; их использование в моделях молекулярной адаптации было популяризировано в 1990-х годах; к 2014 году они были описаны как основная ветвь формальной теории. [19]
Вторая нить — это долгая история попыток утвердить тезис о том, что мутация и развитие оказывают диспозиционное влияние на эволюцию, предоставляя варианты для последующей функциональной оценки, т. е. действуя таким образом, который логически предшествует отбору. Многие эволюционные мыслители предлагали ту или иную форму этой идеи. В начале 20-го века такие авторы, как Эймер и Коуп, считали, что развитие настолько сильно ограничивает или направляет эволюцию, что эффект отбора имеет второстепенное значение. [20] [21] Ранние генетики, такие как Морган и Паннетт, предположили, что общие параллелизмы (например, меланизм или альбинизм ) могут отражать вероятные мутационные изменения. [22] Expanding on Vavilov's (1922) [23] исследование этой темы, Spurway (1949) [24] писал, что « спектр мутаций группы может быть более важным, чем многие ее морфологические или физиологические особенности».
Аналогичное мышление проявилось при появлении эво-дево , например, Альберч (1980) предполагает, что «в эволюции отбор может определить победителя в данной игре, но развитие определяет игроков неслучайно» (стр. 665). [25] (см. также [26] ). Томсон (1985), [27] обзор нескольких томов, посвященных новому мышлению, основанному на развитии, - книга Раффа и Кауфмана (1983) [28] и тома конференций под редакцией Боннера (1982) [29] и Гудвин и др. (1983) [30] —писал: «Вся суть эволюционистского подхода к эволюции состоит в том, чтобы изучить возможность того, что асимметрия во внесении вариаций на фокальном уровне отдельных фенотипов, возникающая из-за присущих свойств развивающихся систем, представляет собой мощный источник причинно-следственных связей в эволюционных изменениях». (с. 222). Точно так же палеонтологи Элизабет Врба и Найлс Элдридж резюмировали эту новую теорию развития, заявив, что «предвзятость во введении фенотипической изменчивости может быть более важной для направленной фенотипической эволюции, чем сортировка путем отбора». [31]
Однако идея влияния развития на эволюцию была отвергнута Майром и другими, такими как Мейнард Смит («Если мы хотим понять эволюцию, мы должны помнить, что это процесс, который происходит в популяциях, а не у отдельных людей»). [32] и Брюс Уоллес («проблемы, связанные с упорядоченным развитием личности, не связаны с проблемами эволюции организмов во времени»), [33] как несовместимое с общепринятыми концепциями причинности. Этот конфликт между эво-дево и неодарвинизмом является предметом целой книги философа Рона Амундсона. [34] (см. также Шолль и Пильуччи, 2015 г.) [35] ). В теории эволюции как изменения частот генов, которая преобладала в то время, эволюционные причины — это «силы», которые действуют как массовое давление (т. е. совокупные эффекты бесчисленных отдельных событий), смещающие частоты аллелей (см. гл. 4 книги). [36] ), таким образом, развитие не квалифицируется как эволюционная причина. Широко цитируемый комментарий 1985 года об «ограничениях развития». [37] защищали важность влияний на развитие, но не связывали это утверждение с теорией причинно-следственной связи, недостаток, отмеченный критиками, например, Рив и Шерман (1993) защищали адаптационистскую программу (против сторонников развития и критики адаптационизма Гулда знаменитой и Левонтин ), утверждая, что аргумент «ограничений развития» просто повторяет идею о том, что развитие формирует вариации, не объясняя, как такие предпочтения преобладают над давлением отбора. [38] Майр (1994) [39] настаивал на том, что мышление, направленное на развитие, «безнадежно запутано», поскольку развитие является непосредственной причиной, а не эволюционной. Таким образом, теория развития была воспринята в 1980-х и 1990-х годах как спекуляция без строгого обоснования причинно-следственных теорий, и эта позиция сохраняется (например, Lynch , 2007). [40] ).








В ответ на эти упреки специалисты по развитию пришли к выводу, что популяционная генетика не может дать полного объяснения эволюционных причин : [41] вместо этого сухой статистический отчет об изменениях частот генов, полученный в результате популяционной генетики, должен быть дополнен влажным биологическим отчетом об изменениях в генетической организации развития (так называемый «объяснение происхождения» в [42] ). Убеждения о том, что (1) биология развития никогда не была интегрирована в «Современный синтез» и (2) популяционная генетика должна быть дополнена альтернативными объяснениями причин развития, сейчас широко повторяются в литературе эво-дево и явно приводятся в качестве мотивации для реформ через Расширенный эволюционный синтез . [43]
Предложение признать процесс интродукции формально эволюционной причиной. [3] предлагает иное решение этого конфликта. Согласно этому предложению, ключом к пониманию структуралистского тезиса биологов развития была ранее отсутствующая популяционно-генетическая теория последствий ошибок при внедрении . Авторы раскритиковали классические рассуждения, рассматривающие эффективность вариационных тенденций как вопрос эволюции под воздействием мутационного давления , то есть трансформации популяций посредством повторяющихся мутаций. Они утверждали, что если генеративные предубеждения важны, то это не потому, что они превосходят отбор как силы согласно теории сдвига частот генов, а потому, что они действуют до отбора, посредством внедрения. Таким образом, теория предубеждений прибытия предполагает, что генеративные предрасположенности системы генетики развития (т.е. ее тенденции реагировать на генетические пертурбации предпочтительным образом) формируют эволюцию, опосредуя предвзятости при внедрении. Теория, которая применима как к мутационным отклонениям, так и к отклонениям в развитии, рассматривает вопрос о том, как такие предпочтения могут быть эффективными в формировании хода эволюции, даже когда действует сильный отбор.
Систематические доказательства предсказанных последствий интродукционных ошибок впервые начали появляться в результате экспериментальных исследований адаптации бактерий и вирусов. [44] [9] С 2017 года эта поддержка расширилась и теперь включает систематические количественные результаты лабораторной адаптации и аналогичные, но менее обширные результаты ретроспективного анализа естественных адаптаций, прослеживаемых на молекулярном уровне (см. ниже). Эмпирический случай, согласно которому отклонения в адаптации формы мутации считаются установленными для практических целей, таких как эволюционное прогнозирование (например, [45] ).Однако последствия этой теории не были подвергнуты критической проверке в отношении морфологических и поведенческих особенностей животных и растений, которые являются традиционными объектами эволюционных теоретизирований (см. гл. 9 книги). [15] ). Таким образом, значимость теории для молекулярной адаптации установлена, но значение для эво-дево остается неясным. Эта теория иногда ассоциируется с призывами к реформе со стороны сторонников эво-дево (например, [46] [47] ), хотя он еще не появился в учебниках или в широком рассмотрении проблем эволюционной биологии (например, [48] [49] ).
Простая модель [ править ]
Двойная причинно-следственная связь, предложенная этой теорией, была объяснена с помощью аналогии с «Восхождением на гору Вероятности». [50] [2] Представьте себе робота, который поднимается по пересеченной горной местности, совершая стохастический двухэтапный процесс предложения и принятия. На этапе предложения робот тянется конечностями, чтобы попробовать различные варианты захвата, а на этапе принятия робот фиксирует и меняет свое положение. Если на этапе принятия предпочтение отдается более высоким опорам для рук, альпинист поднимется. Но можно также представить предвзятость на этапе предложения, например, робот может попробовать больше захватов слева, чем справа. Тогда двойной процесс предложения-принятия будет демонстрировать как смещение в сторону повышения из-за предвзятости в принятии, так и смещение влево из-за предвзятости в предложении. Если ландшафт неровный, восхождение закончится на местной вершине, которая (из-за предвзятости предложения) будет располагаться левее начальной точки. На идеально гладком ландшафте альпинист просто будет поворачивать влево, пока не достигнет единственной глобальной вершины.В любом случае траектория альпиниста подвержена двойному смещению. Эти два предубеждения не являются конкурирующими факторами за определение частоты аллелей: они действуют на разных этапах и в неидентичных измерениях.

Двойной эффект, предсказанный теорией, первоначально был продемонстрирован на популяционно-генетической модели одношаговой адаптивной ходьбы с двумя вариантами: [3] т. е. альпинист сталкивается с двумя вариантами восходящего выбора: один с более высоким коэффициентом отбора, а другой с более высокой частотой мутаций. Ключевой особенностью модели является то, что ни одна из альтернатив не присутствует в исходной популяции: их необходимо ввести. При моделировании адаптации в рамках этой модели популяция часто достигает фиксации мутационно предпочтительного аллеля, даже если это не самый подходящий вариант. Форма модели не зависит от того, являются ли отклонения мутационными или эволюционными. Последующая теоретическая работа (ниже) обобщила теорию одношаговых прогулок, а также рассмотрела долгосрочные адаптивные прогулки на сложных фитнес-ландшафтах . Общий смысл параллельной эволюции заключается в том, что отклонения во внедрении могут сильно способствовать параллелизму. Общий вывод о направленности и повторяемости адаптивных блужданий заключается в том, что некоторые пути более благоприятны с эволюционной точки зрения из-за того, что они благоприятны с мутационной точки зрения. Общее значение для долгосрочной предсказуемости результатов, например, определенных фенотипов, заключается в том, что некоторые фенотипы более обнаруживаемы, чем другие, из-за мутационных эффектов, и такие эффекты могут сильно формировать распределение развившихся фенотипов.
Применение теории к проблемам эво-дево и самоорганизации формально опирается на концепцию карты генотип-фенотип (ГП). Генетический код , например, представляет собой карту GP, которая вызывает асимметрию в мутационно доступных фенотипах. Рассмотрим эволюцию фенотипа Met (аминокислоты), кодируемого генотипом ATG (кодон). Фенотипический сдвиг от Met к Val требует мутации ATG на GTG; переход от Met к Leu может произойти в результате двух разных мутаций (ATG на CTG или TTG); переход от Met к Ile может произойти в результате трех различных мутаций (в ATT, ATC или ATA). Если каждый тип генетической мутации имеет одинаковую частоту, т. е. без мутационной ошибки как таковой , карта GP индуцирует 3 разные скорости внедрения альтернативных фенотипов Val, Leu и Ile. Из-за этой предвзятости при внедрении предпочтение отдается эволюции от Met до Ile, и это происходит не из-за мутационной предвзятости (в смысле предвзятости, отражающей механизмы мутагенеза), а скорее из-за асимметричного сопоставления фенотипов с мутационно доступными генотипами.
Результаты теоретического моделирования [ править ]
Одношаговые адаптивные прогулки [ править ]
Как отмечалось выше, в простейшем случае эффекта «Вероятное восхождение на гору» можно рассматривать альпиниста, стоящего перед только двумя фиксированными вариантами выбора: вверх и влево или вверх и вправо. Этот случай моделируется с помощью моделирования, [3] и получает более полное лечение [4] В общем, ограничивающее поведение эволюции, когда поступление новых мутаций становится сколь угодно малым, т.е. , называется динамикой «начала-фиксации». . [19] Приближение фиксации начала координат для выбора между левым и правым вариантами и (соответственно) в модели Ямпольского-Штольцфуса определяется следующим образом:



| ( 1 ) |

где (или ) и (или ) — частота мутаций и коэффициент отбора для левой (или правой) альтернативы, и если предположить, что вероятность фиксации . В модели Ямпольского-Штольцфуса это приближение хорошо для .








Для одношаговых блужданий в условиях фиксации начала координат поведение, определяемое уравнением ( 1 ), обобщает 2 на множество альтернатив. Например, Кано и др. (2022) [7] Рассмотрим модельный ген с множеством различных полезных мутаций, и при низком количестве мутаций эффекты предвзятости мутаций пропорциональны спектру адаптивных изменений.
Когда не очень мал, разные полезные аллели могут присутствовать одновременно, конкурируя и замедляя адаптацию; эффект, известный как клональная интерференция . Клональная интерференция снижает эффект предвзятости мутаций в моделях эволюции в конечных генетических пространствах: аллели, которым благоприятствует мутация, по-прежнему имеют тенденцию появляться раньше, но до того, как они достигнут фиксации, возникающие позже более полезные аллели могут превзойти их, усиливая эффект. различий в физической подготовке. В самых экстремальных условиях, когда все возможные полезные аллели надежно присутствуют в большой популяции, наиболее подходящий аллель побеждает детерминированно и нет места для эффекта мутационной предвзятости. [7] [51] Другими словами, когда присутствуют все полезные аллели и отбор определяет победителя, шанс на успех равен 1 для наиболее подходящего аллеля и 0 для всех остальных аллелей. Таким образом, в модели гена с конечным набором полезных мутаций ожидается, что влияние ошибки мутации будет наиболее сильным, когда но упасть как становится большим.


Влияние мутации при различной степени клональной интерференции можно точно оценить количественно с помощью регрессионного метода Кано и др. (2022). [7] Предположим, что ожидаемое количество изменений данного класса мутационных изменений, определяемых начальным и конечным состояниями, прямо пропорционально произведению (1) частоты исходного состояния и (2) скорости мутаций возведен во власть , то есть,


| ( бета1 ) |
![{\displaystyle \mathbb {E} [\mathbf {n} (c,a)]\proto f(c)\mu (c,a)^{\beta }}](https://wikimedia.org/api/rest_v1/media/math/render/svg/355d325e1a0c36f225dc77d52a790b3c41848c7b)
Логарифмирование этого уравнения дает
| ( бета2 ) |
![{\displaystyle \log \mathbb {E} [\mathbf {n} (c,a)]=\alpha +\log f(c)+\beta \log \mu (c,a)}](https://wikimedia.org/api/rest_v1/media/math/render/svg/65973dd2513b9ab8265a8bdf95f1bb38b9a1ba01)
где – логарифм константы пропорциональности. Таким образом, когда неизвестно, его можно оценить как коэффициент регрессии log(counts) на log(expected counts). Моделирование модели гена (рисунок справа от [7] ) показать диапазон от при высоком уровне мутаций в когда запас мутаций низкий. Хотя этот подход был разработан для оценки того, как спектр мутаций влияет на адаптивные миссенс-изменения (определяемые стартовым кодоном и конечной аминокислотой), уравнение отражает общую структуру, применимую к любым классам изменений, определяемым мутациями.



Обратите внимание, что эти соображения применимы к конечным генетическим пространствам. В бесконечном генетическом пространстве клональная интерференция по-прежнему замедляет скорость адаптации из-за конкуренции, но не предотвращает эффект мутационной предвзятости, поскольку среди наиболее подходящего класса аллелей всегда есть мутационно предпочтительные альтернативные аллели. [52] [6]
мутации параллелизм Вклад в
В общем случае, если существует некоторый набор возможных шагов, каждый из которых имеет вероятность , то вероятность параллельности определяется суммированием квадратов: . Это следует из определения дисперсии или коэффициент вариации , что (см. врезку 2 [52] или Ч. 8 из [15] )




| ( 2 ) |

То есть параллелизм увеличивается за счет всего, что уменьшает количество вариантов выбора или увеличивает неоднородность их шансов (измеряемых или ). Этот результат подтверждает интуицию Шулля. [53] что «вера в законы случайности требует напряжения веры в то, что одни и те же изменения должны происходить снова и снова, если возможности безграничны, а вероятности равны» (стр. 448). В той степени, в которой неоднородность отражает гетерогенность мутационных шансов, мутация способствует параллелизму.
В частности, для случая динамики начала-фиксации каждое значение является продуктом термина мутационного происхождения и термина фиксации, так что гетерогенность в любом из них одинаково способствует шансам на параллелизм, и можно разделить эффекты мутации и отбора при учете повторяемости эволюции. [52] В условиях начала-фиксации и при условии , отсюда следует, что [54]

| ( 3 ) |

где и – коэффициенты вариации для векторов коэффициентов отбора и частоты мутаций соответственно. Числовые примеры во вставке 2 [52] предполагают, что мутация иногда больше способствует параллелизму, чем отбору, хотя авторы отмечают, что в знаменателе выше скрыто смешивает эффекты мутации и отбора (поскольку на практике отражает набор путей, которые достаточно благоприятствуются отбором и с достаточной вероятностью наблюдаемых мутаций).



поиска возможность и
Для систематического взгляда на долгосрочные эффекты эволюции в дискретном генотипическом пространстве рассмотрим четыре точки зрения, представленные ниже, сосредоточив внимание на влиянии спектра мутаций (характерного для некоторой развивающейся системы) на различные способы определения шансов эволюции (после лечения). к [52] ):
- точка с доступом к другим близлежащим точкам в пространстве генотипов . От , существует 0 или более шагов или путей вверх, которые различаются мутационной благоприятностью (как функция спектра мутаций ) и преимуществами приспособленности. Эволюционность-из- является функцией этого набора шагов. В простых условиях каждый шаг имеет вероятность а повторяемость эволюции получается путем возведения в квадрат ценности.
- Непустое множество шагов на пути или траверсе возрастающей приспособленности, т. е. адаптивном пути (можно также рассмотреть нейтральный путь или путь неубывающей приспособленности). Каждый путь имеет длину и состав с точки зрения преимуществ приспособленности шагов и мутационной благоприятности шагов. Вероятность того, что эволюция пойдет по заданному пути, должна каким-то образом зависеть от этих свойств по отношению к другим возможным путям.
- Агрегированный набор путей («бассейн притяжения»), которые ведут к заданному пункту назначения, например, к пику или плато приспособленности, или набор шагов в фенотипической сети. Любой пункт назначения можно обнаружить по 0 или более восходящим путям, соединяющим его с нижними точками. Точки в этой коллекции также могут иметь пути к другим пунктам назначения. Для данного пункта назначения возможность развития или находимости зависит от этого набора путей относительно конкурирующих путей. Каждая коллекция имеет некоторый общий размер, т. е. может быть много или мало путей, ведущих к месту назначения.
- , Фитнес-ландшафт который может включать в себя множество вершин и троп . В зависимости от набора вершин и троп ландшафт может быть более или менее пригодным для навигации в том смысле, что у него высока вероятность найти вершину с высокой приспособленностью из случайно выбранной отправной точки. Судоходство по ландшафту будет зависеть от спектра мутаций в зависимости от состава путей в ландшафте .

Доступны теоретические результаты, относящиеся к каждой из этих точек зрения. Например, при моделировании адаптивного блуждания генов, кодирующих белок, в контексте абстрактного ландшафта NK , [8] Эффект смещения мутации GC-AT заключается в изменении состава белковой последовательности способом, качественно соответствующим аналогии с восхождением на гору Вероятность (выше). Каждое адаптивное блуждание начинается со случайной последовательности и заканчивается на некотором локальном пике; направление блуждания и конечный пик зависят от смещения мутации. [8] Например, адаптивное блуждание при смещении мутации в сторону GC приводит к появлению белков, которые содержат больше аминокислот с кодонами, богатыми GC (Gly, Ala, Arg, Pro), и аналогичным образом адаптивное блуждание при смещении AT приводит к появлению белков с большим количеством аминокислоты с АТ-богатыми кодонами (Phe, Tyr, Met, Ile, Asn, Lys). На пересеченной местности первоначальный эффект аналогичен, но адаптивные прогулки короче. [8] То есть смещение мутации накладывает предпочтение (при адаптивном блуждании) на шаги, пути и локальные пики, которые обогащены результатами, которым благоприятствует смещение мутации. Это иллюстрирует концепцию направленного тренда, при котором система совокупно движется в определенном направлении вдоль оси состава.
Влияние систематической ошибки перехода-трансверсии было изучено с использованием эмпирических ландшафтов приспособленности для сайтов связывания транскрипционных факторов. [55] Каждый ландшафт основан на создании тысяч различных 8-нуклеотидных фрагментов и измерении того, насколько хорошо они связываются с определенным фактором транскрипции. К каждому пику на каждом ландшафте можно добраться по некоторому набору путей, состоящих из ступеней, представляющих собой замены нуклеотидов, каждая из которых представляет собой либо переход, либо трансверсию. Среди всех возможных генетических изменений соотношение переходов и трансверсий составляет 1:2. Однако совокупность путей, ведущих к данной вершине (на данном эмпирическом ландшафте), имеет специфический переходно-трансверсионный состав, который может отличаться от 1:2. Аналогичным образом, любая развивающаяся система имеет определенную тенденцию к мутации переход-трансверсия. Чем точнее предвзятость мутации (развивающейся системы) соответствует предвзятости состава (ландшафта), тем больше вероятность того, что развивающаяся система найдет пик. [55] Таким образом, для данной развивающейся системы с ее характерной склонностью к переходу-трансверсии некоторые ландшафты более удобны для навигации, чем другие. Навигация максимизируется, когда предвзятость мутаций развивающейся системы соответствует предвзятости состава ландшафта. [55]
Наконец, вместо того, чтобы организовывать генотипы по приспособленности (в терминах пиков, восходящих путей и совокупности путей, ведущих к пику), мы можем организовать генотипы по фенотипу, используя карту генотип-фенотип . Данный фенотип идентифицирует сеть в пространстве генотипов, включающую все генотипы с этим фенотипом. [56] Эволюционирующая система может нейтрально диффундировать внутри сети генотипов с одним и тем же фенотипом, но предполагается, что конверсии между фенотипами не нейтральны. Каждая фенотипически определенная сеть имеет возможность обнаружения, которая в первом приближении является функцией количества генотипов в сети. [5] [57] Например, если использовать канонический генетический код в качестве карты генотип-фенотип, фенотип лейцина имеет 6 кодонов, тогда как триптофан имеет 1: лейцин более доступен для обнаружения, поскольку существует больше мутационных путей от нелейциновых генотипов. Эту идею можно применить к тому, как складки РНК (рассматриваемые как фенотипы) сопоставляются с последовательностями РНК. Например, эволюционное моделирование показывает, что складки РНК с большим количеством последовательностей легче обнаруживаются, и это связано с тем, что в результате мутации они подвергаются избыточной выборке. [5] Аналогичный момент был высказан в отношении подструктур регулирующих сетей. [58] (см. также [57] ).
Приведенные выше результаты, как и ранее, применимы к конечным пространствам. В бесконечных пространствах набор оставшихся полезных мутаций, которые предстоит изучить, бесконечен и включает в себя бесконечное количество мутационно благоприятных и мутационно неблагоприятных вариантов. Следовательно, эволюция в бесконечных пространствах может продолжаться вечно в направлении, предпочтительном для мутаций, без уменьшения мутационного эффекта, который применяется в краткосрочной перспективе, например, можно рассмотреть уравнение ( 1 ) для такого бесконечного пространства. Модель Гомеса и др. (2020) [6] допускает неограниченную адаптацию за счет двух признаков: один с более высокой частотой полезных мутаций, а другой с более крупными селективными преимуществами. В этой модели смещение мутаций продолжает играть важную роль в долгосрочной эволюции, даже когда предложение мутаций очень велико.
последствия Отличительные
Теорию предубеждений в процессе интродукции как причину ориентации или направления эволюции можно противопоставить другим теориям, которые использовались биологами-эволюционистами для рассуждений о роли изменчивости в эволюции:
- Организмы адаптивно реагируют на условия жизни, и эти реакции передаются по наследству ( ламаркизм ). Обычно считается, что этой теории не хватает механистической основы.
- Вариации поставляют сырье, преобразованное в адаптации путем отбора ( неодарвинизм ). Согласно этой теории, «отбор — единственный фактор, задающий направление эволюции». [59] в то время как вариация является материальной причиной — просто пассивным источником субстанции, а не источником формы, инициативы или направления (обеспечиваемого отбором) — так что законы вариации «не имеют никакого отношения» к структурам, построенным отбором. [60]
- Развитие накладывает предварительные ограничения на форму. В этой народной теории «отбор может определить победителя в данной игре, но развитие определяет игроков не случайным образом». Эта теория появилась в классических аргументах таких авторов, как Эймер и Коуп; [20] он вновь появился в заявлениях сторонников развития в 1980-х годах. [25] [26]
- Массовое преобразование под воздействием мутационного давления трансформирует популяцию. Последствия этого способа причинности были разработаны главным образом Холдейном. [61] и Кимура, [62] которые сочли это неправдоподобным из-за того, что требовалась высокая частота мутаций, не противоречащая отбору
- Величина постоянной изменчивости усиливает или замедляет вызванные отбором сдвиги в количественных признаках. В эволюционной количественной генетике Матрица (постоянная вариация) является источником размерной, но не направленной асимметрии, в зависимости от количества вариаций, доступных по любому данному измерению в пространстве признаков. [63] [64]

По сравнению с этими теориями теория систематических ошибок прибытия имеет особые последствия, некоторые из которых подтверждаются эмпирически, как описано ниже, например, наиболее частый результат адаптивного процесса, такой как возникновение устойчивости к антибиотикам, не обязательно является наиболее полезным, но часто умеренно благоприятный результат, которому способствует высокая частота мутационного происхождения. Точно так же теория подразумевает, что эволюция может иметь направления, которые не являются адаптивными, или тенденции, которые не оптимальны, - вывод одного из комментаторов книги Артура. [46] нашел «тревожным». [65] Эта теория определяется не какой-либо конкретной проблемой, таксоном, уровнем организации или областью исследования, а механизмом, определенным на уровне популяционной генетики, а именно способностью предвзятостей при интродукции накладывать предвзятости на эволюцию. Некоторые последствия заключаются в следующем (см. [2] ).
Эффекты не требуют нейтральности или высокой скорости мутаций . В отличие от теории эволюции под воздействием мутационного давления, исследованной (и отвергнутой) Холдейном и другими, вариационные диспозиции в рамках теории предубеждений прибытия не зависят от нейтральной эволюции и не требуют высоких скоростей мутаций. [3]
Ступенчатые предубеждения могут иметь градуированные эффекты . В отличие от того, что подразумевается под языком «ограничений» или «пределов», используемым в исторических обращениях к внутренним источникам направления эволюции, теория предубеждений прибытия не является детерминистской и не требует абсолютного различия между возможными и невозможными формами. Вместо этого теория является вероятностной, и градуированные смещения могут иметь градуированные эффекты. [3]
Режимозависимость в отношении популяционной генетики . Согласно теории, предвзятость вариаций не имеет гарантированного эффекта, независимого от деталей популяционной генетики. Влияние мутационных ошибок достигает максимума (пропорциональное влияние) в условиях фиксации происхождения и может почти полностью исчезнуть при высоких уровнях предложения мутаций. [7]
Равенство предубеждений фиксации и предубеждений происхождения (при ограничивающих условиях) . В классическом неодарвинистском мышлении отбор управляет и формирует эволюцию, тогда как вариации играют пассивную роль в поставке материалов. Напротив, при ограниченных условиях фиксации происхождения теория смещений прибытия устанавливает условие паритета, так что (например) 2-кратное смещение при фиксации и 2-кратное смещение при внедрении оказывают одинаковое 2-кратное влияние на шансы на эволюцию. [19]
Общность в отношении источников вариационного смещения . В эволюционной литературе мутационные предубеждения, «ограничения» развития и самоорганизация в смысле находимости рассматриваются как отдельные темы. Согласно теории ошибок прибытия, все это проявления одного и того же популяционно-генетического механизма, в котором ошибки при введении вариантов налагают ошибки на эволюцию. Любое краткосрочное смещение является либо мутационным смещением в смысле разницы в скоростях двух полностью определенных генотипических конверсий, либо его можно рассматривать как схему дифференциальной фенотипической агрегации по генотипам (см. [52] ).
Помимо этих прямых последствий, в литературе появились некоторые более сложные или косвенные последствия.
Непричинные ассоциации, вызванные мутацией и отбором. Из-за двойной зависимости от мутации и отбора распределение адаптивных изменений может демонстрировать непричинные связи частоты мутаций и коэффициентов отбора, что в некоторой степени сродни парадоксу Берксона , как это предполагается в гл. 8 оф. [15] и более подробно разработано Гитшлагом и др. (2023). [66]
Условия состава и декомпозиции причин. При ограниченных условиях фиксации происхождения шансы на эволюцию отражают два фактора, умноженные вместе, представляющие собой ошибки при внедрении и ошибки при фиксации, как в уравнении ( 1 ). Таким образом, существуют условия, при которых можно количественно оценить и напрямую сравнить диспозиционные влияния мутации и отбора. Этот подход уже использовался в нескольких эмпирических аргументах, рассматриваемых ниже. [67] [68] [50] (Вставка 2 из [52] ).
Предвзятое истощение спектра полезных мутаций . В любом случае, когда система адаптируется посредством мутации и отбора, существует некоторый набор возможных полезных мутаций, характеризующийся распределением коэффициентов отбора и скоростей мутаций. Поскольку адаптация происходит на основе мутаций, этот спектр возможных полезных мутаций истощается предвзятым образом. Теория этого истощения [69] имеет отношение к экспериментальной работе, показывающей, что «сдвиги в спектре мутаций расширяют доступ к полезным мутациям». [70] То есть экспериментально наблюдаемая благоприятность сдвигов в спектрах мутаций зависит от паттерна предвзятого истощения полезных мутаций, что само по себе является признаком адаптации, обусловленной мутациями.
Доказательства [ править ]

Доказательства теории были недавно обобщены, например, Gomez et al (2020). [6] представить таблицу, в которой перечислены 8 различных исследований, доказывающих влияние предвзятости мутаций на адаптацию, а Ch. 9 мутаций, случайности и эволюции [15] посвящен эмпирическому подтверждению теории (см. также [2] [52] ). Ожидается, что отклонения во внедрении будут влиять на эволюцию, будь то нейтральную или адаптивную, но влияние на нейтральную эволюцию не считается интуитивно неожиданным или противоречивым, и поэтому ему не уделяется много внимания. Вместо этого в отчетах о доказательствах основное внимание уделяется адаптации, обусловленной мутациями, поскольку это подчеркивает, как предсказания теории противоречат классической концепции мутации как слабого давления, легко преодолеваемого отбором, согласно аргументу Фишера и Холдейна о «противоположном давлении». [3]
Прямые доказательства причинно-следственной связи контролируемых условиях в
Прямые доказательства того, что спектр мутаций формирует спектр адаптивных изменений, получены в исследованиях, которые напрямую манипулируют спектром мутаций . В одном исследовании [9] Устойчивость к цефотаксиму вырабатывалась неоднократно с использованием 3 штаммов E. coli с различным спектром мутаций : дикого типа, mutH и mutT . Спектр мутаций устойчивости среди эволюционировавших штаммов показал те же закономерности спонтанных мутаций, что и родительские штаммы. В частности, трансверсии, которым благоприятствует mutT (левый блок столбцов), высоко распространены среди устойчивых изолятов от родителей mutT (синий на прилагаемом рисунке), и аналогичным образом устойчивые штаммы от родителей mutH (красный) имеют тенденцию иметь мутации нуклеотидного перехода, которым благоприятствует mutH (красный цвет ). центральный блок решеток). Таким образом, изменение спектра мутаций соответствующим образом меняет спектр адаптивных изменений.


Еще одно исследование [12] показали, что штамм AR2 P. fluorescens адаптировался к потере подвижности в подавляющем большинстве случаев (> 95% времени) за счет одного специфического изменения - изменения A289C в гене ntrB , тогда как штамм Pf0-2x адаптировался за счет разнообразных изменений в нескольких генах. . Паттерн в производных AR2 был прослежен до мутационной горячей точки. Поскольку поведение «горячей точки» было связано главным образом с синонимичными различиями между двумя штаммами, экспериментаторы смогли использовать генную инженерию, чтобы удалить «горячую точку» из AR2 и добавить ее к Pf0-2x, не меняя кодируемую аминокислотную последовательность. Это изменило качественную картину результатов: модифицированный AR2 (спроектированный для удаления горячей точки) адаптировался посредством различных изменений, в то время как модифицированный Pf0-2x с спроектированной горячей точкой, адаптированный с помощью A289C, менялся в 80% случаев.
Поэтапные эффекты [ править ]

Другой вариант использования имеющихся данных — сосредоточение внимания на идее градуированных эффектов, которая отличает теорию систематических ошибок прибытия от интуитивного понятия «ограничений» или «пределов» возможных форм. В частности, можно отбросить драматические эффекты, связанные с «горячими точками» и мутаторными аллелями, и рассмотреть эффекты обычных количественных ошибок в нуклеотидных мутациях. Ряд исследований установили, что умеренные, многократные отклонения в мутациях могут иметь многократный эффект на эволюцию, а некоторые исследования указывают на примерно пропорциональную связь между частотой мутаций и шансами на адаптивные изменения. [11] [10] [71] [72] [73]
Например, Сакман и др. (2017). [10] изучил параллельную эволюцию четырех родственных бактериофагов. В каждом случае они параллельно адаптировали 20 культур, а затем секвенировали образец адаптированной культуры для выявления причинных мутаций. Результаты показали явное предпочтение нуклеотидных переходов: 29:5 для путей (белые и черные полосы на рисунке справа) и 74:6 для событий. [10]
В исследовании устойчивости к рифампицину у Pseudomonas aeruginosa , MacLean и др. (2010) [74] измерили коэффициенты отбора и частоту эволюции для 35 мутаций устойчивости в гене rpoB (РНК-полимеразы) и сообщили о частоте мутаций для 11 из них. Частота мутаций варьируется в 30-кратном диапазоне. Частота появления устойчивого варианта в наборе из 284 повторных культур сильно и примерно линейно коррелирует со скоростью мутаций (рисунок справа). Это не объясняется корреляцией между коэффициентами отбора и частотой мутаций, которые не коррелируют (см. гл. 9 книги). [15] ).

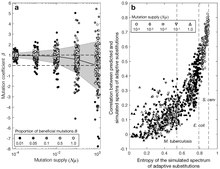
Как объяснялось выше, влияние спектра мутаций на спектр адаптивных изменений можно отразить в одном параметре. , определяемый как коэффициент биномиальной регрессии наблюдаемых значений по ожидаемым значениям из мутационной модели. [7] Основываясь на теоретических соображениях, ожидаемые значения Диапазон значений от 0 (нет влияния) до 1 (пропорциональное влияние). Этот метод был применен Кано и др. [7] к 3 большим наборам данных об адаптивных изменениях, сравнивая модель, основанную на независимых измерениях спектра мутаций , с адаптивными изменениями, ранее выявленными в исследованиях (1) клинической устойчивости микобактерий туберкулеза к антибиотикам , (2) лабораторной адаптации E. coli и ( 3) лабораторная адаптация дрожжей Saccharomyces cerevisiae к стрессу окружающей среды. В каждом случае , что указывает на примерно пропорциональное влияние предвзятости мутации. Авторы сообщают, что это происходит не только из-за влияния предвзятости перехода-трансверсии, поскольку применимо как к систематической ошибке перехода-трансверсии, так и к другим аспектам спектра нуклеотидных мутаций . [7]
Область применения [ править ]
И последнее использование имеющихся данных — рассмотрение диапазона естественных условий, при которых теория может быть применима. Хотя лабораторные исследования могут использоваться для установления причинно-следственной связи и оценки размера эффекта, они не дают прямых указаний о том, где теория применима в природе. Как отмечается в, [52] большинство исследований адаптации не включают генетический анализ, позволяющий выявить конкретные мутации, а в тех редких случаях, когда предпринимаются попытки идентифицировать причинные мутации, результаты обычно указывают лишь на очень небольшое количество изменений, которые подлежат интерпретации. [75] Таким образом, стратегия, которой придерживались в ключевых исследованиях [72] [11] [13] Целью работы было сосредоточиться на проверенных случаях адаптации, в которых предполагаемые функциональные эффекты предполагаемых адаптивных мутаций были проверены с использованием методов генетики.
Пейн и др. [72] искали эффект смещения переходов среди причинных мутаций, вызывающих устойчивость к антибиотикам, в клинически идентифицированных штаммах Mycobacterium Tuberculosis , которые демонстрируют сильную мутационную предвзятость в сторону нуклеотидных переходов. [76] Они сравнили наблюдаемое соотношение переход-трансверсия с нулевым ожиданием 1:2 при отсутствии ошибки мутации. Используя две разные курируемые базы данных, они обнаружили соотношение переход: трансверсия1755:1020 и 1771:900, т.е. обогащение в 3,4 и 3,9 раза больше нуля соответственно. Они также воспользовались особым случаем замены Met-to-Ile, которая может происходить за счет 1 перехода (ATG в ATA) или 2 разных трансверсий (ATG в ATT или ATC). Это соотношение возможностей 1:2 снова представляет собой нулевое ожидание эффектов, независимых от предвзятости мутации. Фактически, мутации в устойчивых изолятах имеют коэффициенты перехода-трансверсии 88:49 и 96:39 (для двух наборов данных), т.е. в 3,6 и 4,9 раза выше нулевых ожиданий. Этот результат не может быть результатом отбора на уровне аминокислот, поскольку все изменения происходят от Met к Ile. Значение этого результата не в том, что смещение мутации работает только тогда, когда варианты выборочно неразличимы: вместо этого урок состоит в том, что смещение в сторону нуклеотидных переходов примерно в 4 раза как для случая Met-to-Ile, так и для случая аминокислот. -изменение замен в целом.
Гораздо более широкий таксономический охват предполагает метаанализ опубликованных исследований параллельной адаптации в природе. [11] В этом исследовании авторы собрали набор данных, охватывающий 10 опубликованных случаев параллельной адаптации, прослеживаемой до молекулярного уровня, включая хорошо известные случаи, связанные с настройкой спектра , устойчивостью к природным токсинам, таким как сердечные гликозиды. [77] и тетродотоксин , [78] [79] брожение передней кишки и так далее. Результаты, показанные ниже (таблица), указывают на соотношение переход-трансверсия 132:99, 2,7-кратное обогащение относительно нулевого ожидания 1:2 (соотношение путей, которое менее чувствительно к экстремальным значениям, составляет 27:28). , 2-кратное обогащение). Таким образом, данное исследование показывает, что склонность к переходам наблюдается в хорошо известных случаях параллельной адаптации у различных таксонов, включая животных и растения.
| Фенотип | Таксон | Цель | Ti считает (пути) | Сумма | Количество ТВ (пути) | ТВ сумма |
|---|---|---|---|---|---|---|
| Устойчивость к инсектицидам | Насекомое | Рдл, Кдр, Туз | 2, 2, 5, 2, 3 (5) | 14 | 9, 2, 4 (3) | 15 |
| Устойчивость к тетродотоксину | Позвоночные животные | На каналы | 2, 6, 3 (3) | 11 | 2, 2, 2, 3, 3 (5) | 12 |
| Устойчивость к гликозидам | многоклеточные животные | 𝑁𝑎+/𝐾+-АТФаза | 4, 4, 2, 2 (4) | 12 | 7, 2, 2, 4 (4) | 15 |
| Устойчивость к гербицидам | Мятликовые | АККейс | 5, 2 (2) | 7 | 7, 2, 4, 5 (4) | 18 |
| Адаптация к высоте | Птицы | 𝛽-гемоглобин | 4, 13 (2) | 17 | 2, 3, 2 (3) | 7 |
| Трихроматическое зрение | Позвоночные животные | Опсины | 2, 5 (2) | 7 | 6, 4, 2 (3) | 12 |
| Эхолокация | Млекопитающие | заем | 2, 2, 2 (3) | 6 | 3, 2 (2) | 5 |
| Рост ритонавира | ВИЧ1 | Протеаза | 25, 7, 9 (3) | 41 | 4 (1) | 4 |
| Ферментация передней кишки | Позвоночные животные | Рибонуклеазы | 2, 4, 4 (3) | 10 | NA (0) | 0 |
| Устойчивость к бензимидазолу | Аскомикота | 𝛽-тубулин | 7 (1) | 7 | 5, 6 (2) | 11 |
| (28) | 132 | (27) | 99 |
Наконец, Шторц и др. [13] проанализировали изменения сродства к гемоглобину, связанные с адаптацией к высоте у птиц. В частности, они изучили эффект смещения CpG — повышенную частоту мутаций в сайтах CpG из-за влияния метилирования цитозина на повреждение и восстановление, широко обнаруженную у млекопитающих и птиц. [80] Они собрали набор данных, состоящий из 35 подобранных пар высокогорных и низкогорных видов птиц. В каждом случае гемоглобины оценивались на предмет функциональных различий, приводящих к более высокому сродству к кислороду у высокогорных видов. Изменения в аффинити, вероятно связанные с адаптацией, включали 10 различных путей, обнаруженных в общей сложности 22 раза. Шесть из 10 путей связаны с мутациями CpG, тогда как только один можно было бы ожидать случайно; и 10 из 22 событий связаны с мутациями CpG, тогда как только 2 можно было бы ожидать случайно (оба различия были значимыми). Это обогащение генетических изменений, вероятных для мутаций, подтверждает теорию ошибок прибытия и предоставляет дополнительные доказательства того, что предсказуемые эффекты ошибок мутаций важны для понимания адаптации в природе.
Контекст эволюционного мышления
Теория смещения прибытия была описана как сквозная теория. [3] [1] [2] [15] потому что он предлагает причинное обоснование (в популяционной генетике) для различных видов ранее существовавших утверждений, для которых причинное обоснование либо неизвестно, либо неверно указано,
- тезис теории развития (выше), согласно которому эволюционные предрасположенности могут возникать в результате того, как развитие формирует вариации, действующие до отбора (например, [26] )
- различные утверждения в литературе по молекулярной эволюции о предвзятых мутационных эффектах (например, при использовании кодонов), которые приписываются неравномерному или направленному «давлению мутаций». [81] но которые не могут быть правдоподобно объяснены как эволюция под воздействием мутационного давления , что вместо этого предполагает необходимость введения теории мутационных предубеждений;
- предположение, возникшее в ходе палеобиологических дебатов 1980-х годов, о том, что в иерархическом расширении эволюционной причинности с уровня популяции на несколько уровней (т. е. популяций, видов, высших таксонов) видообразование является важным источником интродукционных ошибок на уровне высших таксонов. таксоны; [31]
- утверждения о том, что эволюция имеет тенденцию находить фенотипы, чрезмерно представленные в пространстве генотипов, то есть «находимость» или «прибытие частого» [82] эффект, который можно признать как в некоторых аргументах из литературы по молекулярной эволюции, например, King's (1971), так и в [83] объяснение частот аминокислот, а также из литературы по эволюционной самоорганизации , например, аргументов Кауфмана. [84]

Контекст применения теории показан на этом рисунке (справа). Слева показаны детали мутации и развития, ответственные за тенденции возникновения изменчивости (варигенез), т. е. тенденции, предшествовавшие отбору или дрейфу. Справа — наблюдаемые эволюционные закономерности, которые, возможно, можно объяснить этими тенденциями. Зеленая стрелка — это некоторая теория (теория смещения прибытия или некоторая альтернативная теория), которая определяет условия причинно-следственной связи, связывающей вариационные тенденции с эволюционными тенденциями. Применять теорию в этом контексте — значит генерировать эволюционные гипотезы или объяснения, которые апеллируют к внутренним деталям мутации и развития (слева) для объяснения эволюционных закономерностей (справа) через условия причинно-следственной связи, определенные теорией. Например, комментарий Дарвина о том, что законы вариации «не имеют никакого отношения» к структурам, построенным отбором, предполагает, что не существует условий, при которых внутренние детали слева объясняли бы закономерности справа. Все остальные теории предполагают, что вариационные тенденции могут влиять на эволюцию при некоторых условиях. Например, теория Мутационное давление применяется, когда уровень мутаций высок и не встречает сопротивления со стороны отбора, поэтому он имеет ограниченный спектр применения. Теорию эволюционной количественной генетики можно очень широко применить к эволюции количественных признаков, но теория (в том виде, в котором она разработана до сих пор) не предполагает, что мутационные предубеждения будут иметь большое влияние. [85] [86] [87] Напротив, теория смещения прибытия может применяться широко и допускает сильную роль вариационных тенденций в формировании эволюционных тенденций.
Опоздание и неочевидность [ править ]
Хотя сегодня это кажется интуитивно очевидным, формально теория возникла только в 2001 году, например, как отмечалось выше, популяционные генетики не предлагали теорию в 1980-х годах, чтобы ответить на вызов эво-дево, который буквально призывал признать предвзятости при введении вариаций. . Такое позднее появление было объяснено «слепым пятном» из-за множества факторов: [15] включая традицию словесных аргументов, которые минимизируют роль мутации, тенденцию связывать причинно-следственную связь с процессами, которые меняют частоту вариантов, а не с процессами, которые создают варианты, а также формальный аргумент популяционной генетики, который не распространяется на эволюцию от новых мутаций.
В частности, когда Холдейн [61] и Фишер [88] На вопрос, могут ли тенденции мутаций влиять на эволюцию, они сформулировали это как вопрос эффективности мутационного давления (ниже), заключив, что, поскольку частота мутаций низка, мутация является слабой силой, важной только в частном случае аномально высокой мутации. ставки, не встречающие сопротивления со стороны отбора. Их концепция эволюционной причинно-следственной связи была смоделирована на основе отбора, который действует путем изменения частот доступных аллелей, поэтому они относились к повторяющимся мутациям таким же образом. Их вывод верен для случая эволюции от постоянной изменчивости.
В более общем смысле, в рамках теории современного синтеза предполагалось, что эволюция является результатом кратковременного процесса изменения частот доступных аллелей. [89] [90] В этом процессе мутация обычно не имеет значения, за исключением случаев, когда основное внимание уделяется низкочастотным аллелям, поддерживаемым вредным мутационным давлением (см. Популяционная генетика: Мутация ), например, Эдвардс (1977). [91] обратился к теоретической популяционной генетике, вообще не рассматривая мутации; Левонтин (1974) [92] заявил, что «в детерминистской теории практически нет качественных или грубых количественных выводов о генетической структуре популяций, чувствительных к небольшим значениям миграции или каких-либо выводов, зависящих от скорости мутаций» (стр. 267).
Аргумент Холдейна-Фишера о «противостоящем давлении» неоднократно использовался ведущими мыслителями, чтобы отвергнуть структуралистское или интерналистское мышление (примеры в [2] или Ч. 6 из [15] ), например, Фишер (1930) заявил, что «Вся группа теорий, которые приписывают гипотетическим физиологическим механизмам, контролирующим возникновение мутаций, способность направлять ход эволюции, должна быть отброшена, как только смешанная теория наследственности будет доказана». Семьдесят лет спустя Гулд (2002), цитируя Фишера (1930), написал, что «Поскольку ортогенез может работать только тогда, когда давление мутаций становится достаточно высоким, чтобы выступать в качестве агента эволюционных изменений, эмпирические данные о низкой частоте мутаций звучат как приговор. звон интернализма». (стр. 510) [93]

Таким образом, аргументы популяционной генетики использовались для отклонения, а не для поддержки спекулятивных утверждений о роли вариационных тенденций. Ошибка в аргументе Холдейна-Фишера, указанная в: [3] заключается в том, что он рассматривает мутацию только как давление на частоты существующих аллелей, а не как причину возникновения новых аллелей. Когда аллели, имеющие отношение к результату эволюции, изначально отсутствуют, ошибки при интродукции могут привести к сильным ошибкам в результате. Таким образом, позднее появление этой теории проливает свет на то, насколько тесно мышление современного синтеза было связано с предположением о постоянных вариациях. [95] и к теории сил. [15] Эти обязательства продолжают отражаться в современных источниках, например, в официальном документе США, одобренном SSE , SMBE , ASN , ESA и другими соответствующими профессиональными обществами, Futuyma и др. (2001). [94] констатируют как факт, что эволюция меняет частоты генов, определяя основные причины «эволюции» (определенной таким образом) как отбор и дрейф (рисунок).
Однако к концу ХХ века теоретики начали отмечать, что долговременная динамика зависит от событий внедрения мутаций, не освещенных классической теорией. [16] [18] [17] В современной литературе предположение об эволюции в результате постоянной изменчивости лишь изредка делается явным, например: [96] . [97] Чаще всего эволюцию из существующих вариаций рассматривают как вариант, который следует рассматривать вместе с эволюцией из новых мутаций. [98] [19] [99]
для проблем Актуальность современных

Параллелизм и предсказуемость . Применение теории к параллелизму рассмотрено выше. Тенденция к повторению определенных результатов в эволюции является не просто функцией отбора, но также отражает предвзятости при внедрении из-за дифференциальной доступности посредством мутации (или, в случае фенотипов, мутации и измененного развития). Последние обзоры о предсказаниях [52] [45] применить теорию к роли мутационных ошибок в обеспечении повторяемости эволюции.
Разделение причинной ответственности за закономерности на мутацию и отбор . В случае динамики происхождения-фиксации эволюционные предрасположенности можно объяснить сочетанием мутаций и отбора, и в принципе возможно распутать эти вклады, как при анализе регуляторных и структурных эффектов в эволюции. [67] или закономерности замены аминокислот в эволюции белков. [50]
Эво-дево , карты ГП и находимость . Применение теории смещений прибытия к развитию и фенотипам опосредовано концепцией карты генотип-фенотип. Простой пример смещения, вызванного картой GP, показан справа. Эволюционирующая система нейтрально диффундирует внутри генотипической сети своего фенотипа и может время от времени переходить к другому фенотипу. Из стартовой сети генотипов, кодирующих фенотип P0, происходят мутации, ведущие к генотипическим сетям P1 и P2. Однако количество мутаций, ведущих от стартовой сети к P2, в 4 раза выше, что иллюстрирует идею о том, что для данной онтогенетической системы некоторые фенотипы более мутационно доступны. Это не то же самое, что мутационное смещение как таковое (асимметрия, вызванная деталями мутагенеза), но оно может иметь тот же эффект в популяционно-генетической модели. [3] В этом случае, если все мутации происходят с одинаковой скоростью, общая скорость мутационного внедрения P2 в 4 раза выше, чем для P1: это смещение может быть отображено в модели Ямпольского-Штольцфуса и будет иметь те же последствия, что и 4- смещение складчатой мутации.

Для краткосрочной эволюции важно распределение сразу доступных для мутаций фенотипов. Однако в долгосрочной эволюции можно ожидать два разных эффекта, которые можно объяснить рисунком справа после рисунка 4 Фонтаны (2002). [56] Сети показывают генотипы, которые соответствуют трем различным фенотипам: P0, P1 и P2. Со временем система может нейтрально диффундировать среди разных генотипов с одинаковыми фенотипами. Редко может произойти переход от одного фенотипа к другому. В краткосрочной перспективе эволюция зависит только от того, что непосредственно доступно из данной точки генотипического пространства. В среднесрочной перспективе эволюция зависит от доступности альтернативных фенотипических сетей по отношению к исходной сети, например, начиная с P0, P2 в два раза доступнее, чем P1, даже несмотря на то, что P1 и P2 имеют одинаковое количество генотипов. В долгосрочной перспективе значение имеет общая находимость фенотипа среди всех других фенотипов, которая (в первом приближении) зависит от количества генотипов (а точнее, от общей площади поверхности сети). доступный для других фенотипов высокой приспособленности). В этом случае P0 более доступен для обнаружения, чем P1 и P2, поскольку у него вдвое больше генотипов. Вариационная предвзятость в сторону более многочисленных фенотипов названа Динглом и др. (2022) «предвзятостью фенотипа». [82] (см. также [57] ).

Этот эффект находимости является формальной основой для эмпирических и теоретических аргументов в исследованиях находимости мотивов регуляторной сети. [58] или семейства складок РНК. [82] На рисунке справа Дингл и др. (2022) [82] представили доказательства поразительной тенденции наиболее распространенных в природе складок РНК соответствовать складкам, наиболее широко распространенным в пространстве последовательностей.
Роль в широких реформам к призывах
Теория предвзятости прибытия, предложенная в 2001 году, появляется в нескольких последующих призывах к реформе по сравнению с неодарвинистским взглядом на современный синтез. Пер Артур , [46] это часть подхода к эволюции, основанного на развитии, который подчеркивает внутренние организующие эффекты «перепрограммирования развития» на изменчивость. В другом кадре, [15] [2] эффективность предубеждений прибытия подрывает историческую приверженность теоретиков рассматривать эволюцию как процесс смещения частот генов в богатом генофонде, в котором доминируют силы массового действия, и является частью более широкого движения (начавшегося во время молекулярной революции) от неодарвинизм современного синтеза и версия мутационизма, основанная на популяционной генетике. Эта теория также упоминалась в литературе расширенного эволюционного синтеза под заголовком « Ошибка развития» . [47]
от других теорий мутационных Отличие эффектов
Теория предубеждений прибытия фокусируется на своего рода популяционно-генетической причинно-следственной связи, связывающей внутренние генеративные предубеждения, действующие до отбора, с предсказуемыми эволюционными тенденциями. Она отличается от других идей, которым не хватает такого же внимания к причинно-следственной связи, внутренним предубеждениям или процессу внедрения.
Эволюция под воздействием мутаций [ править ]

В классических источниках эволюция под действием «мутационного давления» означает массовую трансформацию популяции путем мутационной конверсии, как у Уилсона и Боссерта (1971, стр. 42). [100] [101] ). Общая оценка этой теории по Холдейну (1932) [102] и Фишер (1930), [88] заключается в том, что эволюция под действием мутационного давления неправдоподобна, поскольку она требует высокой частоты мутаций, не противоречащей отбору. Кимура еще более пессимистично утверждал, что трансформация под воздействием мутационного давления займет так много времени, что ее можно будет игнорировать в практических целях. [62] Тем не менее, более поздние эмпирические и теоретические работы показали, что эта теория может быть ценной в таких случаях, как потеря сложного признака, кодируемого многими локусами, например, потеря споруляции в экспериментальных популяциях B. subtilis , случай, когда частота мутаций для утрата признака оценивалась как необычайно высокая величина, . [103]

Таким образом, теория мутационного давления и теория предвзятости прибытия описывают способы оказания важного влияния на процесс мутации, но они фокусируются на разных способах причинно-следственной связи: влиянии либо на процесс фиксации (давление мутации), либо на процесс внедрения ( смещение прибытия). Эффективность мутационных тенденций этими двумя способами совершенно различна, например, только теория мутационного давления опирается на высокие темпы мутаций, которым не противодействует отбор.
наименьшего сопротивления Эволюция по генетическим линиям
Эволюционная количественная генетика, совокупность теорий, фокусирующаяся на высокополигенных количественных признаках, делает конкретные предсказания о мутационных эффектах, которые имеют некоторую эмпирическую поддержку. В стандартной теории набора количественных признаков постоянная вариация представлена матрица дисперсий и ковариаций, которая зависит (сложным образом) от мутационных входных данных, представленных матрица. Фенотипическая дивергенция будет иметь тенденцию соответствовать (в фенотипическом пространстве) размеру наибольшей изменчивости, , и этот предсказанный эффект изменения стояния наблюдался неоднократно. [104] [105] Этот эффект (более подробно объясненный в разделе «Ошибка развития ») называется адаптацией «по генетическим линиям наименьшего сопротивления» и может быть переформулирован (с вариациями в положительной роли) как адаптация по линиям максимального вариационного топлива. Когда дивергенция также совпадает с Это предполагает, что мутационная изменчивость формирует дивергенцию, но эта косвенная корреляция имеет другие интерпретации и не принимается в качестве убедительного доказательства. [106]



Использование мутационной предвзятости в смысле асимметричного воздействия на средние значения признаков не является частью стандартной схемы. Когда смещение мутации включается в модели одного количественного признака при стабилизирующем отборе, результатом является небольшое отклонение от оптимального значения. [87] [86] [85]
Таким образом, модели эволюционной количественной генетики фокусируются на проблемах другого рода, так что не существует простого перевода между (например) эффектами и влияние предубеждений при внедрении.
Мутационная непредвиденность [ править ]
Эволюционные объяснения часто опирались на парадигму «равновесного объяснения» (гл. 5 книги «Объяснение равновесия»). [36] ) в котором результаты объясняются обращением к выборочно оптимальному, без учета истории или деталей процесса.(как поясняется в [38] )Однако в последние десятилетия внимание сосредоточилось на идее «непредвиденности», то есть идее о том, что результат эволюции не может быть объяснен как предсказуемая или заранее определенная конечная точка детерминированного процесса, а идет по некоторому пути, который невозможно легко предсказать или можно предсказать только зная детали начальных условий и последующей динамики. «Мутационная» непредвиденная ситуация относится к случаям, в которых событие эволюции отчетливо связано с конкретной мутацией или мутационной горячей точкой, например. [107] в том смысле, что эволюционное изменение не произошло бы наблюдаемым образом, если бы отличительная мутация не произошла предполагаемым образом.
Это понятие отличается от теории предубеждений в процессе внедрения, поскольку оно представляет собой объяснительную концепцию (а не механизм), применяемую в идиографических объяснениях, т. е. объясняющих разовые события (символические события), [108] Теория предубеждений в процессе внедрения — это теория общей причинно-следственной связи: результатом успешного применения теории является присвоение не символического объяснения, а общего объяснения, подобного этой схеме, в которой случается чаще, чем вызвано предвзятостью при внедрении из-за более высокой вероятности мутационно-развитой конверсии .


Ограничения развития ( развития предвзятость )
Понятие «ограничение» чревато. [109] Грин и Джонс (2016) [110] утверждают, что биологи-эволюционисты используют его как гибкую объяснительную концепцию, а не как способ ссылки на конкретную причинную теорию, т. е. ограничение — это фактор с некоторым ограничивающим влиянием, которое делает его прогнозирующим, даже если причинная основа этого влияния неясна. .
Простое понятие ограничения развития состоит в том, что некоторые фенотипические формы не наблюдаются из-за невозможности (или, по крайней мере, очень сложно) создать их в процессе развития. [111] , например, многоножки с четным числом сегментов, несущих ноги. [112] То есть ограничение — это объяснение отсутствия фенотипов, основанное на вариационном эффекте (отсутствии) в рамках парадигмы, ориентированной на учет закономерностей существования фенотипов.
Другие ссылки на «ограничение» подразумевают скорее градуированные различия, чем абсолютную разницу между возможными и невозможными формами, например: [37] В то время как эффективность абсолютных предубеждений не требует специальной причинной теории (поскольку невозможная с точки зрения развития форма является эволюционно невозможной формой), идея градуированных предубеждений поднимает вопросы причинности из-за конфликта с классическим «противоположным давлением» Холдейна-Фишера. аргумент , который утверждает, что простые вариационные тенденции неэффективны, поскольку уровень мутаций невелик. Основополагающая статья Мейнарда Смита и др. «Ограничения развития» (1985) [37] отметил эту проблему, не предоставив решения. Сторонников «ограничений» критиковали за неспособность обеспечить механизм. [38] Это вопрос, над которым Ямпольский и Штольцфус [3] стремился исправить.
Тем не менее, теорию систематических ошибок прибытия нелегко сопоставить с концепцией «ограничения», поскольку последнее широко используется как синоним слова «фактор». В литературе эво-дево термин «ограничение» все чаще заменяется ссылками на отклонения в развитии . Однако концепция систематической ошибки развития часто связана с некоторой идеей облегченной изменчивости или эволюционности , тогда как теория систематической ошибки прибытия касается только популяционно-генетических последствий произвольных ошибок в возникновении вариаций.
изменчивость, эволюционность и мутация Облегченная направленная
Теория предубеждений прибытия не требует и не подразумевает облегченных вариаций или направленных мутаций и сама по себе не является теорией эволюции способности к эволюции . Популяционно-генетические модели, используемые для иллюстрации теории, и эмпирические случаи, приведенные в поддержку теории, сосредоточены на эффектах различных форм мутационной предвзятости, где предвзятость всегда связана с каким-то измерением, отличным от приспособленности, например, с переходом. трансверсионная предвзятость, [72] смещение CpG, [13] или асимметрия двух черт с разной изменчивостью. [6] То есть теория не предполагает, что предубеждения полезны в отношении приспособленности, и не предполагает, что мутация каким-то образом способствует адаптации отдельно от эффекта отбора (против [113] ). Фактически, многие модели иллюстрируют эффективность систематических ошибок прибытия, сосредотачиваясь на случае, когда наиболее благоприятные с точки зрения мутаций результаты не являются наиболее подходящими вариантами, как в исходной модели Ямпольского-Штольцфуса, где один вариант имеет более высокую частоту мутаций, но меньшую пригодность. преимущество, а другой имеет более высокую физическую выгоду, но меньшую частоту мутаций. Теория не предполагает ни того, что мутационно благоприятные результаты являются более подходящими, ни того, что они менее подходят.
См. также [ править ]
- Предвзятость развития
- Развиваемость
- Расширенный эволюционный синтез
- Предвзятость мутации
- Популяционная генетика
Ссылки [ править ]
- ↑ Перейти обратно: Перейти обратно: а б А. Штольцфус (2006). «Мутационизм и двойная причинность эволюционных изменений» . Эвол Дев . 8 (3): 304–317. дои : 10.1111/j.1525-142X.2006.00101.x . ПМИД 16686641 . S2CID 10469049 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г А. Штольцфус (2019). «Понимание предвзятости при введении вариаций как эволюционной причины». В Уллере, Т.; Лаланд, КНЦ (ред.). Эволюционная причинность: биологические и философские размышления . Кембридж, Массачусетс: MIT Press.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к Л.Я. Ямпольский и А. Штольцфус (2001). «Предвзятость во введении вариаций как ориентирующий фактор эволюции». Эвол Дев . 3 (2): 73–83. дои : 10.1046/j.1525-142x.2001.003002073.x . ПМИД 11341676 . S2CID 26956345 .
- ↑ Перейти обратно: Перейти обратно: а б А.А. Соарес; Л. Уордил; Л. Б. Клачко; Р. Дикман (2021). «Скрытая роль мутаций в эволюционном процессе». Физика преп . Э. 104 (4): 044413. Бибкод : 2021PhRvE.104d4413S . дои : 10.1103/PhysRevE.104.044413 . ПМИД 34781575 . S2CID 239954422 .
- ↑ Перейти обратно: Перейти обратно: а б с Шапер, С.; Луи, А.А. (2014). «Прибытие частого: как смещение в картах генотип-фенотип может направить популяцию к локальному оптимуму» . ПЛОС ОДИН . 9 (2): e86635. Бибкод : 2014PLoSO...986635S . дои : 10.1371/journal.pone.0086635 . ПМЦ 3914804 . ПМИД 24505262 .
- ↑ Перейти обратно: Перейти обратно: а б с д и К. Гомес; Дж. Бертрам; Дж. Масель (2020). «Мутационная предвзятость может повлиять на адаптацию в больших бесполых популяциях, испытывающих клональное вмешательство» . Учеб. Р. Сок. Б. 287 (1937): 20201503. doi : 10.1098/rspb.2020.1503 . ПМЦ 7661309 . ПМИД 33081612 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к А.В. Кано; Х. Рожонова; А. Штольцфус; Д.М. МакКэндлиш; Дж. Л. Пейн (10 февраля 2022 г.). «Смещение мутаций формирует спектр адаптивных замен» . Proc Natl Acad Sci США . 119 (7). Бибкод : 2022PNAS..11919720C . дои : 10.1073/pnas.2119720119 . ПМЦ 8851560 . ПМИД 35145034 .
- ↑ Перейти обратно: Перейти обратно: а б с д А. Штольцфус (2006). «Адаптация, обусловленная мутациями, в модели белка NK». Мол Биол Эвол . 23 (10): 1852–1862. дои : 10.1093/molbev/msl064 . ПМИД 16857856 .
- ↑ Перейти обратно: Перейти обратно: а б с А. Косе, А. Родригес-Рохас и Х. Бласкес (2015). «Обход генетических ограничений во время эволюции мутатора к устойчивости к антибиотикам». Труды Лондонского королевского общества B: Биологические науки . 282 .
- ↑ Перейти обратно: Перейти обратно: а б с д и А. М. Сакман; Л. В. МакГи; Эй Джей Моррисон; Дж. Пирс; Дж. Анисман; Х. Гамильтон; С. Сандербек; К. Ньюман; ДР Рокита (2017). «Параллельная эволюция, обусловленная мутациями, во время вирусной адаптации» . Мол Биол Эвол . 34 (12): 3243–3253. дои : 10.1093/molbev/msx257 . ПМК 5850295 . ПМИД 29029274 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж А. Штольцфус; Д.М. МакКэндлиш (2017). «Мутационные предубеждения влияют на параллельную адаптацию» . Мол Биол Эвол . 34 (9): 2163–2172. дои : 10.1093/molbev/msx180 . ПМК 5850294 . ПМИД 28645195 .
- ↑ Перейти обратно: Перейти обратно: а б Дж. С. Хортон; Л. М. Фланаган; Р.В. Джексон; Н.К. Священник; ТБ Тейлор (2021). «Мутационная горячая точка, определяющая повторяемую эволюцию, может быть создана и разрушена молчаливыми генетическими изменениями» . Нат Коммун . 12 (1): 6092. Бибкод : 2021NatCo..12.6092H . дои : 10.1038/s41467-021-26286-9 . ПМЦ 8526746 . ПМИД 34667151 .
- ↑ Перейти обратно: Перейти обратно: а б с д Дж. Ф. Шторц; К. Натараджан; А. В. Синьор; К.С. Витт; Д.М. МакКэндлиш; А. Штольцфус (2019). «Роль мутационной предвзятости в адаптивной молекулярной эволюции: выводы из конвергентных изменений в функции белка» . Philos Trans R Soc Lond B Biol Sci . 374 : 20180238.doi : (1777 ) 10.1098/rstb.2018.0238 . ПМК 6560279 . ПМИД 31154983 .
- ^ Р. Новик (2023). Структура и функции . Издательство Кембриджского университета, Кембридж. ISBN 9781009013888 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к А. Штольцфус (2021). Мутация, случайность и эволюция . Оксфорд, Оксфорд.
- ↑ Перейти обратно: Перейти обратно: а б Г. Йедид и Г. Белл (2002). «Макроэволюция, моделируемая с помощью автономно воспроизводящихся компьютерных программ». Природа . 420 (6917): 810–2. Бибкод : 2002Natur.420..810Y . дои : 10.1038/nature01151 . ПМИД 12490948 . S2CID 607532 .
Процесс адаптации происходит в двух временных масштабах. В краткосрочной перспективе естественный отбор просто сортирует вариации, уже присутствующие в популяции, тогда как в долгосрочной перспективе генотипы, совершенно отличные от тех, которые изначально присутствовали, развиваются посредством кумуляции новых мутаций. Первый процесс описывается математической теорией популяционной генетики. Однако эта теория начинается с определения фиксированного набора генотипов и не может обеспечить удовлетворительный анализ второго процесса, поскольку не допускает возникновения какого-либо действительно нового типа.
- ↑ Перейти обратно: Перейти обратно: а б И. Эшель и М.В. Фельдман (2001). «Оптимальность и эволюционная стабильность при краткосрочном и долгосрочном отборе». В С. Х. Орзаке и Э. Собере (ред.). Адаптационизм и оптимальность . Издательство Кембриджского университета, Кембридж. стр. 161–190.
Со времен Фишера неявным рабочим предположением при количественном изучении эволюционной динамики является то, что качественные законы, управляющие долгосрочной эволюцией, могут быть экстраполированы на основе результатов, полученных для краткосрочного процесса. Мы считаем, что эта экстраполяция не точна. Эти два процесса качественно отличаются друг от друга. Одно очевидное отличие состоит в том, что мутационные изменения, которые вызывают переход между одним набором частот генотипа, близким к его краткосрочному равновесию, и следующим набором, являются случайными даже в больших популяциях.
- ↑ Перейти обратно: Перейти обратно: а б Д. Л. Хартл и К. Х. Таубс (1998). «К теории эволюционной адаптации». Генетика . 103 : 525–533. дои : 10.1023/А:1017071901530 . S2CID 2057562 .
Почти каждую теоретическую модель популяционной генетики можно отнести к одному из двух основных типов. В одном типе моделей предполагается, что мутации с оговоренными селективными эффектами присутствуют в популяции в качестве начального условия... Второй основной тип моделей допускает возникновение мутаций в случайные промежутки времени, но предполагается, что мутации избирательно нейтральные или почти нейтральные.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г Д.М. МакКэндлиш; А. Штольцфус (2014). «Моделирование эволюции с использованием вероятности фиксации: история и последствия». Ежеквартальный обзор биологии . 89 (3): 225–252. дои : 10.1086/677571 . ПМИД 25195318 . S2CID 19619966 .
- ↑ Перейти обратно: Перейти обратно: а б И. Попов (2009). «Проблема ограничений вариаций от Дарвина до наших дней». Людус Виталис . 17 : 201–220.
- ^ М. А. Улетт (2014). «Обоснование ортогенеза: популяризация определенно направленной эволюции (1890–1926)». Исследования по истории и философии науки. Часть C: Исследования по истории и философии биологических и биомедицинских наук . 45 : 124–132. дои : 10.1016/j.shpsc.2013.11.009 . ПМИД 24368232 .
- ^ А. Штольцфус; К. Кейбл (2014). «Менделизм-мутационизм: забытый эволюционный синтез» . J Hist Biol . 47 (4): 501–46. дои : 10.1007/s10739-014-9383-2 . ПМИД 24811736 . S2CID 254552058 .
- ^ Н. И. Вавилов (1922). «Закон гомологического ряда в вариациях». Журнал наследственности . 12 : 47–89.
- ^ Х. Спурвей (1949). «Замечания о законе гомологичной вариации Вавилова». Ла Рицерка Научная . 18 :18–24.
Данный вид, семейство, отряд или класс может легче мутировать в сторону определенных фенотипов, чем другие. . . С эволюционной точки зрения спектр мутаций группы может быть более важным, чем многие ее морфологические или физиологические особенности. Они [sic] определяют возможности его эволюции. Большинство из них не эксплуатируются.
- ↑ Перейти обратно: Перейти обратно: а б П. Альберх (1980). «Онтогенез и морфологическое разнообразие» . Американский зоолог . 20 (4): 653–667. дои : 10.1093/icb/20.4.653 .
- ↑ Перейти обратно: Перейти обратно: а б с Г. Остер; П. Альберх (1982). «Эволюция и бифуркация программ развития». Эволюция . 36 (3): 444–459. дои : 10.1111/j.1558-5646.1982.tb05066.x . ПМИД 28568040 . S2CID 773428 .
в эволюции выбор может определить победителя в данной игре, но развитие определяет игроков неслучайно (стр. 665)
- ^ К.С. Томсон (1985). «Обзор эссе: взаимосвязь между развитием и эволюцией». Оксфордские обзоры по эволюционной биологии . 2 : 220–233.
- ^ Р.А. Рафф; Т. К. Кауфман (1983). Эмбрионы, гены и эволюция . Макмиллан, Нью-Йорк.
- ^ Дж. Т. Боннер, изд. (1982). Эволюция и развитие: отчет Далемского семинара по эволюции и развитию, Берлин, 1981 г., 10–15 мая . Спрингер-Верлаг, Нью-Йорк.
- ^ BC Гудвин, Н. Холдер и CC Уайли, изд. (1983). Развитие и эволюция . Издательство Кембриджского университета, Кембридж.
- ↑ Перейти обратно: Перейти обратно: а б Е.С. Врба; Н. Элдридж (1984). «Люди, иерархии и процессы: к более полной эволюционной теории». Палеобиология . 10 (2): 146–171. Бибкод : 1984Pbio...10..146V . дои : 10.1017/S0094837300008149 . S2CID 86346683 .
Биологи развития по-разному подчеркивают: (1) насколько непрямым является генетический контроль на определенных стадиях эпигенеза; (2) система определяет по нисходящей причинно-следственной связи, какие компоненты генома хранятся в неэкспрессируемой форме, а какие выражены в фенотипе; (3) что предвзятость во введении фенотипической изменчивости может быть более важной для направленной фенотипической эволюции, чем сортировка путем отбора.
- ^ Дж. Мейнард Смит (1983). «Развитие и эволюция». В Британской Колумбии Гудвин; Н. Холдер; Си Си Уайли (ред.). Эволюция и развитие: отчет Далемского семинара по эволюции и развитию, Берлин, 1981 г., 10–15 мая . Спрингер-Верлаг, Нью-Йорк.
Это подводит меня к моему последнему пункту. Если мы хотим понять эволюцию, мы должны помнить, что это процесс, происходящий в популяциях, а не у отдельных людей. Отдельные животные могут копать, плавать, лазить или скакать, и они тоже развиваются, но не эволюционируют. Попытка объяснить эволюцию с точки зрения развития индивидуумов означает совершить именно ту ошибку неуместного редукционизма, в которой иногда обвиняют генетиков. 45]
- ^ Б. Уоллес (1986). «Могут ли эмбриологи внести вклад в понимание эволюционных механизмов?». В В. Бектеле (ред.). Интеграция научных дисциплин . Издательство Мартинуса Нийхоффа, Дордрехт. стр. 149–163.
проблемы, связанные с упорядоченным развитием личности, не связаны с проблемами эволюции организмов во времени.
- ^ Р. Амундсон (2005). Меняющаяся роль эмбриона в эволюции . Издательство Кембриджского университета, Кембридж, Великобритания.
Эволюционная нерелевантность биологии развития аргументировалась на философских, методологических, а иногда и исторических основаниях в период с середины двадцатого до конца двадцатого века... Я не знаю, как опровергнуть аргументы нерелевантности неодарвинистов. . Пока неясно, как будет решена эта дилемма.
- ^ Р. Шолль и М. Пильуччи (2015). «Ближайшее и окончательное различие и эволюционная биология развития: причинная нерелевантность против объяснительной абстракции» (PDF) . Биология и философия . 30 (5): 653–670. дои : 10.1007/s10539-014-9427-1 . S2CID 254277341 .
- ↑ Перейти обратно: Перейти обратно: а б Э. Собер (1984). Природа отбора: эволюционная теория в философском фокусе . MIT Press, Кембридж, Массачусетс.
- ↑ Перейти обратно: Перейти обратно: а б с Дж. Мейнард Смит; Р. Буриан; С. Кауфман; П. Альберх; Дж. Кэмпбелл; Б. Гудвин; Р. Ланде; Д. Рауп; Л. Вулперт (1985). «Ограничения развития и эволюция». Кварта. Преподобный биол . 60 : 265–287. дои : 10.1086/414425 . S2CID 85201850 .
Поскольку классическая работа Фишера (1930) и Холдейна (1932) установила слабость направленной мутации по сравнению с отбором, обычно считалось, что направленная предвзятость в изменчивости не приведет к эволюционным изменениям перед лицом противоположного отбора. Эта позиция заслуживает пересмотра.
- ↑ Перейти обратно: Перейти обратно: а б с Х.К. Рив и П.В. Шерман (1993). «Адаптация и цели эволюционных исследований». Ежеквартальный обзор биологии . 68 : 1–32. дои : 10.1086/417909 . S2CID 84402115 .
Здесь могут быть полезны некоторые аналогии. Объясняя, почему в природе преобладают одни черты, а не мыслимые другие, биологи-эволюционисты действуют немного подобно физикам, изучающим стабильные состояния в термодинамических системах, или астрономам, изучающим пространственное расположение звезд и галактик. В каждом случае исследователь изучает продукты исторических процессов. которые разворачиваются на протяжении веков в результате взаимодействия отдельных сущностей. Продукты этих процессов объяснимы, а иногда даже предсказуемы, без детального знания индивидуальной истории каждого объекта. Это происходит потому, что при данном наборе условий большое количество различных индивидуальных историй имеют тенденцию сходиться в небольшом количестве стабильных состояний. Например, наблюдение, что валуны собираются в горных долинах, можно объяснить, не зная точной траектории каждого валуна.
- ^ Э. Майр (1994). «Ответ Джону Битти». Биология и философия . 9 (3): 357–358. дои : 10.1007/BF00857941 . S2CID 84453520 .
Что меня поражает, так это то, насколько в современной литературе это различие [ближайшее-конечное] до сих пор игнорируется или путается. Должно быть, за последние два года я прочитал четыре или пять статей и одну книгу о развитии и эволюции. Развитие, расшифровка генетической программы, очевидно, является вопросом непосредственных причин. Столь же очевидно, что эволюция — это вопрос эволюционных причин. И все же во всех этих статьях и в этой книге оба вида причинности были безнадежно перепутаны.
- ^ М. Линч (2007). «Хрупкость адаптивных гипотез происхождения сложности организма» . Proc Natl Acad Sci США . 104 (Приложение 1): 8597–604. дои : 10.1073/pnas.0702207104 . ПМК 1876435 . ПМИД 17494740 .
- ^ А. С. Уилкинс (1998). «Эволюционная биология развития: куда идет?». Биоэссе . 20 (10): 783–784. doi : 10.1002/(SICI)1521-1878(199810)20:10<783::AID-BIES1>3.0.CO;2-R . S2CID 85320436 .
- ^ Б. Калкотт (2009). «Объяснения происхождения: объяснение того, как меняются биологические механизмы». Британский журнал философии науки . 60 : 51–78. дои : 10.1093/bjps/axn047 . hdl : 1885/50236 .
- ^ Л. Чиу (2022). Расширенный эволюционный синтез. Обзор последних научных исследований . Фонд Джона Темплтона, Вест Коншохокен, Пенсильвания, США. doi : 10.15868/socialsector.40950 . S2CID 252252438 .
- ^ Д.Р. Рокита; П. Джойс; С.Б. Кодл; Х.А. Вичман (2005). «Эмпирическая проверка модели адаптации мутационного ландшафта с использованием одноцепочечного ДНК-вируса». Нат Жене . 37 (4): 441–4. дои : 10.1038/ng1535 . ПМИД 15778707 . S2CID 20296781 .
- ↑ Перейти обратно: Перейти обратно: а б М.Т. Вортель; Д. Агаше; С.Ф. Бэйли; и др. (2023). «На пути к эволюционным предсказаниям: текущие обещания и проблемы» . Эволюционные приложения . 16 (1): 3–21. дои : 10.1111/eva.13513 . ПМК 9850016 . ПМИД 36699126 .
В широком спектре таксонов мутационная ошибка объясняет немаловажную долю случаев параллельной генетической эволюции (Stern & Orgogozo 2008, Bailey et al. 2017, 2018, Stoltzfus & McCandlish 2017). Например, элегантное исследование адаптации птиц к большой высоте обнаружило параллельную эволюцию, отчасти из-за смещения мутаций в сайтах CpG (Storz et al. 2019). Когда из-за существующей мутационной предвзятости выборка весьма полезных мутаций недостаточна, вместо этого могут исправиться другие мутации с меньшим эффектом, но более частые. Такая закономерность наблюдалась в реплицированных эволюционирующих популяциях бактериофагов (Sackman et al. 2017), где мутация с наибольшим эффектом приспособленности не была той, которая чаще всего достигала фиксации, поскольку частота ее мутаций была ниже, чем у других мутаций с меньшей приспособленностью. эффекты.
- ↑ Перейти обратно: Перейти обратно: а б с В. Артур (2004). Предвзятые эмбрионы и эволюция . Издательство Кембриджского университета, Кембридж.
- ↑ Перейти обратно: Перейти обратно: а б Т. Уллер; А.П. Мочек; Р. А. Уотсон; премьер-министр Брейкфилд; К. Н. Лаланд (2018). «Предвзятость развития и эволюция: взгляд на сеть регулирования» . Генетика . 209 (4): 949–966. дои : 10.1534/genetics.118.300995 . ПМК 6063245 . ПМИД 30049818 .
- ^ Дж. Б. Лосос; С. Дж. Арнольд; Г. Бехерано; и др. (2013). «Эволюционная биология XXI века» . ПЛОС Биол . 11 (1): e1001466. дои : 10.1371/journal.pbio.1001466 . ПМЦ 3539946 . ПМИД 23319892 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Джонатан Б. Лосос; Дэвид А. Баум; Дуглас Дж. Футуйма; Хопи Э. Хукстра; Ричард Э. Ленски; Аллен Дж. Мур; Кэтрин Л. Пайчел; Дольф Шлютер; Майкл Дж. Уитлок, ред. (2014). Принстонский путеводитель по эволюции . Издательство Принстонского университета.
- ↑ Перейти обратно: Перейти обратно: а б с А. Штольцфус и Л.Я. Ямпольский (2009). «Восхождение на гору Вероятности: мутация как причина неслучайности в эволюции» . Дж. Херед . 100 (5): 637–47. дои : 10.1093/jhered/esp048 . ПМИД 19625453 .
- ^ С.Ф. Бэйли; Ф. Бланкар; Т. Батайон; Р. Кассен (2017). «Что движет параллельной эволюцией?: Как размер популяции и мутационные вариации способствуют повторной эволюции» . Биоэссе . 39 (1): 1–9. doi : 10.1002/bies.201600176 . ПМИД 27859467 . S2CID 193595 .
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж А.В. Кано; Б. Гитшлаг; Х. Рожонова; А. Штольцфус; Д.М. МакКэндлиш; Дж. Л. Пейн (2023). «Мутационная предвзятость и предсказуемость эволюции» . Учеб. Р. Сок. Б. 378 (1877): 20220055. doi : 10.1098/rstb.2022.0055 . ПМЦ 10067271 . ПМИД 37004719 .
- ^ А. Ф. Шулль (1935). «Вейсман и Геккель: сто лет». Наука . 81 (2106): 443–451. Бибкод : 1935Sci....81..443S . дои : 10.1126/science.81.2106.443 . ПМИД 17818758 .
- ^ Л. М. Шевен, Г. Мартин и Т. Ленорман (2010). «Модель Фишера и геномика адаптации: ограниченная плейотропия, гетерогенная мутация и параллельная эволюция» . Эволюция . 64 (11): 3213–31. дои : 10.1111/j.1558-5646.2010.01058.x . ПМИД 20662921 . S2CID 205782829 .
- ↑ Перейти обратно: Перейти обратно: а б с Кано, Алехандро; Пейн, Джошуа Л. (2020). «Предвзятость мутаций взаимодействует с предвзятостью состава, влияя на адаптивную эволюцию» . PLOS Вычислительная биология . 16 (9): e1008296. Бибкод : 2020PLSCB..16E8296C . дои : 10.1371/journal.pcbi.1008296 . ПМЦ 7571706 . ПМИД 32986712 .
- ↑ Перейти обратно: Перейти обратно: а б В. Фонтана (2002). «Моделирование «эво-дево» с помощью РНК» . Биоэссе . 24 (12): 1164–77. дои : 10.1002/bies.10190 . ПМИД 12447981 .
- ↑ Перейти обратно: Перейти обратно: а б с Дж. Гарсон, Л. Ван и С. Саркар (2003). «Как развитие может направлять эволюцию». Биология и философия . 18 (2): 353–370. дои : 10.1023/А:1023996321257 . S2CID 83110476 .
- ↑ Перейти обратно: Перейти обратно: а б К. Сюн; М. Герштейн; Дж. Масель (2021). «Различия в эволюционной доступности определяют, какой одинаково эффективный регуляторный мотив развивается для генерации импульсов» . Генетика . 219 (3). дои : 10.1093/генетика/iyab140 . ПМЦ 8570775 . ПМИД 34740240 .
- ^ Э. Майр (1980). «Некоторые мысли об истории эволюционного синтеза». В Э. Майре и В. Провайне (ред.). Эволюционный синтез: перспективы унификации биологии . Издательство Гарвардского университета. стр. 1–48.
- ^ К. Дарвин (1868). Изменение животных и растений при одомашнивании . Мюррей, Лондон.
На протяжении всей этой главы и в других местах я говорил об отборе как о первостепенной силе, однако его действие абсолютно зависит от того, что мы по своему невежеству называем спонтанной или случайной изменчивостью. Пусть архитектору придется построить здание из необработанных камней, упавших с пропасти. Форму каждого фрагмента можно назвать случайной; однако форма каждого из них определялась силой гравитации, природой скалы и уклоном пропасти — событиями и обстоятельствами, все из которых зависят от законов природы; но нет никакой связи между этими законами и целью, для которой строитель использует каждый фрагмент. Точно так же вариации каждого существа определяются фиксированными и неизменными законами; но они не имеют никакого отношения к живой структуре, которая медленно выстраивается посредством отбора, будь то естественный или искусственный отбор [курсив добавлен редактором]
- ↑ Перейти обратно: Перейти обратно: а б Дж. Б. С. Холдейн (1927). «Математическая теория естественного и искусственного отбора. V. Отбор и мутация». Учеб. Кэмб. Фил. Соц . 26 : 220–230. дои : 10.1017/S0305004100015644 . S2CID 86716613 .
- ↑ Перейти обратно: Перейти обратно: а б М. Кимура (1980). «Среднее время до фиксации мутантного аллеля в конечной популяции под постоянным давлением мутаций: исследования аналитическими, численными методами и методами псевдовыборки» . Proc Natl Acad Sci США . 77 (1): 522–526. Бибкод : 1980PNAS...77..522K . дои : 10.1073/pnas.77.1.522 . ПМК 348304 . ПМИД 16592764 .
- ^ Р. Ланде (1979). «Количественный генетический анализ многомерной эволюции применительно к мозгу: аллометрия размеров тела». Эволюция . 33 (1Часть 2): 402–416. дои : 10.1111/j.1558-5646.1979.tb04694.x . ПМИД 28568194 . S2CID 17132606 .
- ^ Р. Ланде и С. Дж. Арнольд (1983). «Измерение отбора коррелирующих символов». Эволюция . 37 (6): 1210–1226. дои : 10.2307/2408842 . JSTOR 2408842 . ПМИД 28556011 .
- ^ Дж. Брукфилд (2005). «Откуда берется разнообразие животных?» . Современная биология . 15 (22): Р908–Р910. дои : 10.1016/j.cub.2005.11.007 . S2CID 10714428 .
- ^ Б.Л. Гитшлаг; А.В. Кано; Дж. Л. Пейн; Д.М. МакКэндлиш; А. Штольцфус (2023). «Мутация и отбор вызывают корреляцию между коэффициентами отбора и скоростью мутаций». Американский натуралист . 202 (4): 534–557. дои : 10.1086/726014 . hdl : 20.500.11850/639862 . ПМИД 37792926 . S2CID 256902349 .
- ↑ Перейти обратно: Перейти обратно: а б М. А. Стрейсфельд и М. Д. Раушер (2011). «Популяционная генетика, плейотропия и преимущественная фиксация мутаций в ходе адаптивной эволюции» . Эволюция . 65 (3): 629–42. дои : 10.1111/j.1558-5646.2010.01165.x . ПМИД 21054357 . S2CID 33188741 .
- ^ А. Фарлоу, Э. Медури и К. Шлоттерер (2011). «Репарация двухцепочечных разрывов ДНК и эволюция плотности интронов» . Тенденции в генетике . 27 (1): 1–6. дои : 10.1016/j.tig.2010.10.004 . ПМК 3020277 . ПМИД 21106271 .
- ^ М. Туффаха; С. Варакунан; Д. Кастеллано; РН Гутенкунст; Л. М. Валь (2022). «Изменения в предвзятости мутаций способствуют развитию мутаторов, изменяя распределение эффектов приспособленности». bioRxiv : 2022.09.27.509708. дои : 10.1101/2022.09.27.509708 . S2CID 252684737 .
- ^ М. Сане; Г.Д. Диван; Б.А. Бхат; Л. М. Валь; Д. Агаше (2023). «Сдвиги в спектре мутаций расширяют доступ к полезным мутациям» . Proc Natl Acad Sci США . 120 (22): e2207355120. Бибкод : 2023PNAS..12007355S . дои : 10.1073/pnas.2207355120 . ПМЦ 10235995 . ПМИД 37216547 .
- ^ Дж. Ю. Фам; КБ Огбунугафор; А.Н. Нгуен Ба; Д.Л. Хартл (2019). «Экспериментальная эволюция ширины ниши бактериофага Т4 подчеркивает важность структурных генов» . МикробиологияОткрыть . 9 (2): е968. дои : 10.1002/mbo3.968 . ПМК 7002106 . ПМИД 31778298 .
- ↑ Перейти обратно: Перейти обратно: а б с д Дж. Л. Пейн; Ф. Менардо; А. Траунер; С. Боррель; С.М. Гигли; К. Луазо; С. Ганье; АР Холл (2019). «Смещение при переходе влияет на эволюцию устойчивости к антибиотикам у микобактерий туберкулеза» . ПЛОС Биол . 17 (5): e3000265. дои : 10.1371/journal.pbio.3000265 . ПМК 6532934 . ПМИД 31083647 .
- ^ А. Штольцфус; Д.М. МакКэндлиш (2015). «Адаптация андских крапивников, обусловленная мутациями» . Proc Natl Acad Sci США . 112 (45): 13753–4. Бибкод : 2015PNAS..11213753S . дои : 10.1073/pnas.1518490112 . ПМЦ 4653193 . ПМИД 26489650 .
- ^ Р. К. Маклин; Г.Г. Перрон; А. Гарднер (2010). «Уменьшающаяся отдача от полезных мутаций и повсеместного эпистаза формируют ландшафт устойчивости к рифампицину у Pseudomonas aeruginosa» . Генетика . 186 (4): 1345–54. дои : 10.1534/genetics.110.123083 . ПМК 2998316 . ПМИД 20876562 .
- ^ Д. Кумбс; Дж. В. Б. Мойр; А. М. Пул; Т. Ф. Купер; RCJ Добсон (2019). «Фитнес-задача изучения молекулярной адаптации» . Биохим Соц Транс . 47 (5): 1533–1542. дои : 10.1042/BST20180626 . ПМИД 31642877 . S2CID 204849713 .
- ^ Р. Хершберг и Д.А. Петров (2010). «Доказательства того, что мутация у бактерий повсеместно смещена в сторону АТ» . ПЛОС Генет . 6 (9): e1001115. дои : 10.1371/journal.pgen.1001115 . ПМЦ 2936535 . ПМИД 20838599 .
- ^ Б. Уджвари; Н. Р. Кейсвелл; К. Сунагар; К. Арбакл; В. Вустер; Н. Ло; Д. О'Мили; К. Бекманн; Г. Ф. Кинг; Е. Деплазес; Т. Мэдсен (2015). «Широко распространенное сближение устойчивости к токсинам благодаря предсказуемой молекулярной эволюции» . Proc Natl Acad Sci США . 112 (38): 11911–6. Бибкод : 2015PNAS..11211911U . дои : 10.1073/pnas.1511706112 . ПМЦ 4586833 . ПМИД 26372961 .
- ^ Ч.Р. Фельдман; ЭД Броди младший; ЭД Броди, третий; М. Е. Пфрендер (2012). «Ограничения формируют конвергенцию в устойчивых к тетродотоксину натриевых каналах змей» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (12): 4556–61. дои : 10.1073/pnas.1113468109 . ПМК 3311348 . ПМИД 22392995 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ МС Йост; Д.М. Хиллис; Ю. Лу; Дж. В. Кайл; ХА Фоззард; Его Святейшество Закон (2008). «Устойчивые к токсинам натриевые каналы: параллельная адаптивная эволюция всего семейства генов» . Молекулярная биология и эволюция . 25 (6): 1016–24. дои : 10.1093/molbev/msn025 . ПМЦ 2877999 . ПМИД 18258611 .
- ^ Л. Смедс, А. Кварнстрем и Х. Эллегрен (2016). «Прямая оценка скорости мутации зародышевой линии у птицы» . Геном Рез . 26 (9): 1211–8. дои : 10.1101/гр.204669.116 . ПМК 5052036 . ПМИД 27412854 .
- ^ Н. Суэока (1988). «Направленное мутационное давление и нейтральная молекулярная эволюция» . Proc Natl Acad Sci США . 85 (8): 2653–7. Бибкод : 1988PNAS...85.2653S . дои : 10.1073/pnas.85.8.2653 . ПМК 280056 . ПМИД 3357886 .
- ↑ Перейти обратно: Перейти обратно: а б с д и К. Дингл; Ф. Гаддар; П. Сульк; А. А. Луи (2022). «Смещение фенотипа определяет, как естественные структуры РНК занимают морфопространство всех возможных форм» . Мол Биол Эвол . 39 . дои : 10.1093/molbev/msab280 . ПМЦ 8763027 . ПМИД 34542628 .
- ^ Дж. Л. Кинг (1971). «Влияние генетического кода на эволюцию белка». В Э. Шоффениэльсе (ред.). Биохимическая эволюция и происхождение жизни . Издательство Северной Голландии. стр. 3–13.
Частота каждой аминокислоты пропорциональна вероятности ее появления в результате мутации.
- ^ С.А. Кауфман (1993). Истоки порядка: Самоорганизация и эволюция . Издательство Оксфордского университета, Нью-Йорк.
- ↑ Перейти обратно: Перейти обратно: а б Б. Чарльзуорт (2013). «Стабилизирующий отбор, очищающий отбор и мутационная предвзятость в конечных популяциях» . Генетика . 194 (4): 955–71. дои : 10.1534/генетика.113.151555 . ПМЦ 3730922 . ПМИД 23709636 .
- ↑ Перейти обратно: Перейти обратно: а б XS Чжан и WG Hill (2008). «Повторный обзор аномальных эффектов предвзятой мутации: отклонение среднего от оптимального и очевидный направленный отбор в условиях стабилизирующего отбора» . Генетика . 179 (2): 1135–41. дои : 10.1534/genetics.107.083428 . ПМЦ 2429868 . ПМИД 18558659 .
- ↑ Перейти обратно: Перейти обратно: а б Д. Ваксман и Дж. Р. Пек (2003). «Аномальные эффекты предвзятой мутации» . Генетика . 164 (4): 1615–26. дои : 10.1093/генетика/164.4.1615 . ПМЦ 1462658 . ПМИД 12930765 .
- ↑ Перейти обратно: Перейти обратно: а б Р. А. Фишер (1930). Генетическая теория естественного отбора . Издательство Оксфордского университета, Лондон.
- ^ Т. Добжанский (1937). Генетика и происхождение видов . Издательство Колумбийского университета, Нью-Йорк.
- ^ В. Б. Провайн (1971). Истоки теоретической популяционной генетики . Издательство Чикагского университета, Чикаго.
- ^ AWF Эдвардс (1977). Основы математической генетики . Издательство Кембриджского университета, Нью-Йорк.
Все гены будут считаться стабильными и мутации не будут учитываться [с. 3]
- ^ Р. К. Левонтин (1974). Генетическая основа эволюционных изменений . Издательство Колумбийского университета, Нью-Йорк.
Детерминистская теория критически зависит от параметров отбора. В детерминистской теории практически нет качественных или грубых количественных выводов о генетической структуре популяций, чувствительных к малым значениям миграции или каких-либо выводов, зависящих от скорости мутаций.[p. 267]
- ^ С. Дж. Гулд (2002). Структура эволюционной теории . Издательство Гарвардского университета, Кембридж, Массачусетс.
- ↑ Перейти обратно: Перейти обратно: а б Диджей Футуйма (2001). «Эволюция, наука и общество». Американский натуралист . 158 : S1–S46. дои : 10.1086/509090 . ПМИД 18707297 . S2CID 41606175 .
- ^ Дж. Битти (2022). «Синтез и два сценария». Эволюция . 76 (С1): 6–14. дои : 10.1111/evo.14423 . ПМИД 34951705 . S2CID 245445537 .
- ^ SC Stearns и RF Hoekstra (2005). Эволюция: введение . Издательство Оксфордского университета, Нью-Йорк.
скорость эволюционных изменений не ограничивается появлением новых благоприятных мутаций (с. 99)
- ^ Р. Докинз (2007). «Обзор: Грань эволюции» . Интернэшнл Геральд Трибьюн (29 июня).
Если расчеты Бехи верны, они сразу же смутят поколения математических генетиков, которые неоднократно доказывали, что темпы эволюции не ограничиваются мутациями. В одиночку Бихи сразится с Рональдом Фишером, Сьюэллом Райтом, Дж.Б.С. Холдейном, Теодосием Добжански, Ричардом Левонтином, Джоном Мейнардом Смитом и сотнями их талантливых коллег и интеллектуальных потомков. Несмотря на неудобное существование собак, капусты и голубей, весь корпус математической генетики с 1930 года по сегодняшний день совершенно ошибочен. Майкл Бихи, биохимик из Университета Лихай, от которого отреклись, — единственный, кто правильно подвел итоги. Вы думаете? Лучший способ выяснить это — отправить Бехи математическую статью, скажем, в «Журнал теоретической биологии» или «Американский натуралист», редакторы которого разошлют ее квалифицированным рецензентам.
- ^ Дж. Хермисон и П.С. Пеннингс (2005). «Мягкие зачистки: молекулярно-популяционная генетика адаптации к устойчивым генетическим вариациям» . Генетика . 169 (4): 2335–52. doi : 10.1534/genetics.104.036947 . ПМЦ 1449620 . ПМИД 15716498 .
- ^ Д. Хоул, Б. Морикава и М. Линч (1996). «Сравнение мутационных изменчивостей» . Генетика . 143 (3): 1467–83. дои : 10.1093/генетика/143.3.1467 . ПМК 1207413 . ПМИД 8807316 .
- ^ Э.О. Уилсон и У.Х. Босссерт (1971). Букварь популяционной биологии . Зинауер Сандерленд, Массачусетс.
- ^ Онлайн-демонстрация включена в образовательные инструменты популяционного генетика Джона Макдональда по адресу [1] )
- ^ Дж. Б. С. Холдейн (1932). Причины эволюции . Лонгманс, Грин и Ко, Нью-Йорк.
- ^ Х. Моган; Дж. Масель; CW Бирки-младший; У.Л. Николсон (2007). «Роль накопления мутаций и отбора в потере споруляции в экспериментальных популяциях Bacillus subtilis» . Генетика . 177 (2): 937–48. doi : 10.1534/genetics.107.075663 . ПМК 2034656 . ПМИД 17720926 .
- ^ Д. Шлютер (1996). «Адаптивная радиация по генетическим линиям наименьшего сопротивления». Эволюция . 50 (5): 1766–1774. дои : 10.2307/2410734 . JSTOR 2410734 . ПМИД 28565589 .
- ^ К. Макгиган (2006). «Изучение фенотипической эволюции с использованием многомерной количественной генетики» . Мол Экол . 15 (4): 883–96. дои : 10.1111/j.1365-294X.2006.02809.x . ПМИД 16599954 . S2CID 32969263 .
- ^ Д. Хоул; Г.Х. Болстад; К. ван дер Линде; ТФ Хансен (2017). «Мутация предсказывает 40 миллионов лет эволюции маховых крыльев». Природа . 548 (7668): 447–450. Бибкод : 2017Natur.548..447H . дои : 10.1038/nature23473 . HDL : 11250/2453538 . ПМИД 28792935 . S2CID 90225702 .
- ^ Т. Ленорман, Д. Розе и Ф. Руссе (2009). «Стохастичность в эволюции». Тенденции Эколь Эвол . 24 (3): 157–65. дои : 10.1016/j.tree.2008.09.014 . ПМИД 19178980 .
- ^ Дж. Л. Кинг (2017). «Возвращение к чудесной жизни: случайность и непредвиденные обстоятельства в эдиакарско-кембрийской радиации». В Г. Рэмси и Ч. Пенсе (ред.). Шанс в эволюции . Издательство Чикагского университета.
- ^ Дж. Антонович и П. Х. ван Тиндерен (1991). «Онтоэценофилоограничения? Хаос терминологии ограничений». Тенденции Экол. Эвол . 6 (5): 166–168. дои : 10.1016/0169-5347(91)90059-7 . ПМИД 21232448 .
- ^ С. Грин и Н. Джонс (2016). «Рассуждения на основе ограничений для поиска и объяснения: стратегии понимания изменчивости и закономерностей в биологии» (PDF) . Диалектика . 70 (3): 343–374. дои : 10.1111/1746-8361.12145 .
- ^ М. Е. Олсон (2012). «Ренессанс развития в адаптационизме». Тенденции Эколь Эвол . 27 (5): 278–87. дои : 10.1016/j.tree.2011.12.005 . ПМИД 22326724 .
- ^ В. Артур и М. Фэрроу (1999). «Характер изменения количества сегментов многоножек как пример ограничения развития в эволюции». J Теория Биол . 200 (2): 183–91. Бибкод : 1999JThBi.200..183A . дои : 10.1006/jtbi.1999.0986 . PMID 10504284 .
- ^ Э.И. Свенссон и Д. Бергер (2019). «Роль мутационной предвзятости в адаптивной эволюции». Тенденции Эколь Эвол . 34 (5): 422–434. дои : 10.1016/j.tree.2019.01.015 . ПМИД 31003616 . S2CID 125066709 .