Популяционная генетика
| Часть серии о |
| Эволюционная биология |
|---|
 |
| Часть серии о |
| Генетическая генеалогия |
|---|
| Концепции |
| Связанные темы |
Популяционная генетика — это раздел генетики , который занимается генетическими различиями внутри и между популяциями и является частью эволюционной биологии . Исследования в этой области биологии изучают такие явления, как адаптация , видообразование и структура популяции . [1]
Популяционная генетика была жизненно важным компонентом возникновения современного эволюционного синтеза . Ее основными основателями были Сьюэлл Райт , Дж.Б.С. Холдейн и Рональд Фишер , которые также заложили основы соответствующей дисциплины количественной генетики . Современная популяционная генетика, традиционно являющаяся высокоматематической дисциплиной, включает теоретические, лабораторные и полевые работы. Популяционно-генетические модели используются как для статистических выводов на основе данных о последовательностях ДНК, так и для доказательства/опровержения концепции. [2]
Что отличает популяционную генетику от более новых, более фенотипических подходов к моделированию эволюции, таких как эволюционная теория игр и адаптивная динамика , так это ее акцент на таких генетических явлениях, как доминирование , эпистаз , степень, в которой генетическая рекомбинация нарушает неравновесие по сцеплению , и случайные явления. мутаций . и дрейфа генетического Это делает его пригодным для сравнения с данными популяционной геномики .
История
[ редактировать ]Популяционная генетика началась как согласование менделевских моделей наследования и биостатистики . Естественный отбор вызовет эволюцию только в том случае, если имеется достаточное количество генетических вариаций в популяции . До открытия менделевской генетики одной из распространенных гипотез была смешанная наследственность . Но при смешанном наследовании генетическая изменчивость будет быстро утрачена, что сделает эволюцию путем естественного или полового отбора неправдоподобной. Принцип Харди-Вайнберга предлагает решение проблемы поддержания изменчивости в популяции с менделевским наследованием. Согласно этому принципу, частоты аллелей (вариаций гена) останутся постоянными при отсутствии отбора, мутаций, миграции и генетического дрейфа. [3]


Следующим ключевым шагом стала работа британского биолога и статистика Рональда Фишера . В серии статей, начавшихся в 1918 году и завершившихся в книге 1930 года «Генетическая теория естественного отбора» , Фишер показал, что непрерывные вариации, измеряемые биометристами, могут быть вызваны совместным действием многих дискретных генов и что естественный отбор может изменить аллель. частоты в популяции, что приводит к эволюции. В серии статей, начавшихся в 1924 году, другой британский генетик, Дж. Б. С. Холдейн , разработал математические расчеты изменения частоты аллелей в одном генном локусе в широком диапазоне условий. Холдейн также применил статистический анализ к реальным примерам естественного отбора, таким как эволюция перцовой моли и промышленный меланизм , и показал, что коэффициенты отбора могут быть больше, чем предполагал Фишер, что приводит к более быстрой адаптивной эволюции в качестве стратегии маскировки после увеличения загрязнения. [4] [5]
Американский биолог Сьюэлл Райт , имевший опыт экспериментов по разведению животных , сосредоточил свое внимание на комбинациях взаимодействующих генов и влиянии инбридинга на небольшие, относительно изолированные популяции, в которых наблюдался генетический дрейф. В 1932 году Райт представил концепцию адаптивного ландшафта и утверждал, что генетический дрейф и инбридинг могут оттолкнуть небольшую изолированную подгруппу населения от пика адаптации, позволяя естественному отбору направить ее к различным пикам адаптации. [ нужна ссылка ]
Работы Фишера, Холдейна и Райта положили начало дисциплине популяционной генетики. Этот интегрированный естественный отбор с менделевской генетикой стал важным первым шагом в разработке единой теории того, как работает эволюция. [4] [5] Джон Мейнард Смит был учеником Холдейна, а У. Д. Гамильтон находился под влиянием работ Фишера. Американец Джордж Р. Прайс работал как с Гамильтоном, так и с Мейнардом Смитом. Американец Ричард Левонтин и японец Мотоо Кимура находились под влиянием Райта и Холдейна. [ нужна ссылка ]
Современный синтез
[ редактировать ]Математика популяционной генетики первоначально была разработана как начало современного синтеза . Такие авторы, как Битти [6] утверждали, что популяционная генетика определяет ядро современного синтеза. В течение первых нескольких десятилетий 20-го века большинство полевых натуралистов продолжали верить, что ламаркизм и ортогенез дают лучшее объяснение сложности, которую они наблюдали в живом мире. [7] В ходе современного синтеза эти идеи были очищены, и остались только эволюционные причины, которые могли быть выражены в математических рамках популяционной генетики. [8] Был достигнут консенсус относительно того, какие эволюционные факторы могут влиять на эволюцию, но не относительно относительной важности различных факторов. [8]
Феодосий Добжанский , постдокторант лаборатории Т. Х. Моргана , находился под влиянием работ по генетическому разнообразию русских генетиков, таких как Сергей Четвериков . » он помог преодолеть разрыв между основами микроэволюции, разработанными популяционными генетиками, и закономерностями макроэволюции , наблюдаемыми полевыми биологами В своей книге 1937 года «Генетика и происхождение видов . Добжанский исследовал генетическое разнообразие диких популяций и показал, что, вопреки предположениям популяционных генетиков, эти популяции обладают большим генетическим разнообразием с заметными различиями между субпопуляциями. Книга также взяла высокоматематическую работу популяционных генетиков и облекла ее в более доступную форму. Через Добжанского влияние популяционной генетики испытало гораздо больше биологов, чем смогли прочитать высокоматематические работы в оригинале. [9]
В Великобритании Э. Б. Форд , пионер экологической генетики , [10] продолжался на протяжении 1930-х и 1940-х годов, чтобы эмпирически продемонстрировать силу отбора, обусловленную экологическими факторами, включая способность поддерживать генетическое разнообразие посредством генетического полиморфизма, такого как группы крови человека . Работа Форда в сотрудничестве с Фишером способствовала смещению акцентов в современном синтезе в сторону естественного отбора как доминирующей силы. [4] [5] [11] [12]
Нейтральная теория и динамика фиксации происхождения
[ редактировать ]Оригинальный современный синтетический взгляд на популяционную генетику предполагает, что мутации предоставляют обильный исходный материал, и фокусируется только на изменении частоты аллелей внутри популяций . [13] Основными процессами, влияющими на частоты аллелей, являются естественный отбор , генетический дрейф , поток генов и рекуррентные мутации . У Фишера и Райта были некоторые фундаментальные разногласия относительно относительной роли отбора и дрейфа. [14] Наличие молекулярных данных обо всех генетических различиях привело к созданию нейтральной теории молекулярной эволюции . С этой точки зрения многие мутации вредны и поэтому никогда не наблюдаются, а большинство остальных нейтральны, т.е. не подвергаются отбору. Поскольку судьба каждой нейтральной мутации оставлена на произвол судьбы (генетический дрейф), направление эволюционных изменений определяется тем, какие мутации происходят, и поэтому не может быть отражено с помощью моделей изменения частоты (существующих) одних только аллелей. [13] [15]
Взгляд на популяционную генетику, основанный на фиксации происхождения, обобщает этот подход за пределы строго нейтральных мутаций и рассматривает скорость, с которой происходит конкретное изменение, как произведение частоты мутаций и вероятности фиксации . [13]
Четыре процесса
[ редактировать ]Выбор
[ редактировать ]Естественный отбор , включающий половой отбор , заключается в том, что некоторые черты повышают шансы организма на выживание и размножение . Популяционная генетика описывает естественный отбор, определяя приспособленность как склонность или вероятность выживания и воспроизводства в определенной среде. Пригодность обычно обозначается символом w =1- s , где s — коэффициент отбора . Естественный отбор действует на фенотипы , поэтому популяционные генетические модели предполагают относительно простые взаимосвязи для прогнозирования фенотипа и, следовательно, приспособленности на основе аллели в одном или небольшом количестве локусов. Таким образом, естественный отбор преобразует различия в приспособленности особей с разными фенотипами в изменения частоты аллелей в популяции на протяжении последующих поколений. [ нужна ссылка ]
До появления популяционной генетики многие биологи сомневались, что небольшие различия в приспособленности достаточны, чтобы иметь большое значение для эволюции. [9] Популяционные генетики частично решили эту проблему, сравнив отбор с генетическим дрейфом . Отбор может преодолеть генетический дрейф, если s больше 1, разделенного на эффективный размер популяции . При выполнении этого критерия вероятность закрепления нового выгодного мутанта примерно равна 2s . [16] [17] Время до фиксации такого аллеля составляет примерно . [18]

Доминирование
[ редактировать ]Доминирование означает, что фенотипический эффект и/или фитнес-эффект одной аллели в локусе зависит от того, какая аллель присутствует во второй копии этого локуса. Рассмотрим три генотипа в одном локусе со следующими значениями приспособленности [19]
| Генотип: | А 1 А 1 | А 1 А 2 | А 2 А 2 |
| Относительная пригодность: | 1 | 1-ч | 1-с |
| Глоссарий популяционной генетики | |
|---|---|
|
s — коэффициент отбора , а h — коэффициент доминирования. Значение h дает следующую информацию:
| ч=0 | А 1 доминантный, А 2 рецессивный |
| ч=1 | А 2 доминантный, А 1 рецессивный |
| 0<ч<1 | неполное доминирование |
| ч<0 | сверхдоминирование |
| ч>1 | Недоминирование |
Эпистаз
[ редактировать ]
Эпистаз означает, что фенотипический эффект и/или приспособленность аллели в одном локусе зависит от того, какие аллели присутствуют в других локусах. Отбор действует не на один локус, а на фенотип, возникающий в результате развития полного генотипа. [20] Однако многие модели популяционной генетики половых видов представляют собой модели «единого локуса», в которых приспособленность особи рассчитывается как произведение вкладов каждого из ее локусов, фактически предполагая отсутствие эпистаза.
На самом деле, генотип и фитнес-ландшафт более сложны. Популяционная генетика должна либо детально смоделировать эту сложность, либо уловить ее с помощью какого-то более простого правила усреднения. Эмпирически полезные мутации, как правило, имеют меньшую выгоду от приспособленности, если их добавить к генетическому фону, который уже имеет высокую приспособленность: это известно как эпистаз с убывающей отдачей. [21] Когда вредные мутации также оказывают меньший эффект на приспособленность на фоне высокой приспособленности, это известно как «синергический эпистаз». Однако эффект вредных мутаций в среднем имеет тенденцию быть очень близким к мультипликативному или даже проявлять противоположную картину, известную как «антагонистический эпистаз». [22]
Синергический эпистаз занимает центральное место в некоторых теориях очистки от мутационной нагрузки. [23] и к эволюции полового размножения .
Мутация
[ редактировать ]
Генетический процесс мутации происходит внутри человека, приводя к наследственным изменениям генетического материала. Этот процесс часто характеризуется описанием начального и конечного состояний или типа изменения, произошедшего на уровне ДНК (например, мутация Т-на-С, делеция 1 п.н.), генов или белков (например, нулевая мутация, мутация с потерей функции) или на более высоком фенотипическом уровне (например, мутация «красных глаз»). Однонуклеотидные изменения часто являются наиболее распространенным типом мутаций, но многие другие типы мутаций возможны и , которые происходят с очень разной частотой, что может проявлять систематическую асимметрию или систематическую ошибку ( мутационная ошибка ).
больших участков ДНК Мутации могут включать в себя дублирование , обычно посредством генетической рекомбинации . [24] Это приводит к изменению количества копий внутри популяции. Дупликации являются основным источником сырья для развития новых генов. [25] Другие типы мутаций иногда создают новые гены из ранее некодирующей ДНК. [26] [27]
При распределении эффектов приспособленности (DFE) для новых мутаций только небольшая часть мутаций приносит пользу. Мутации с грубыми последствиями обычно вредны. Исследования на мухе Drosophila melanogaster показывают, что если мутация изменяет белок, вырабатываемый геном, это, вероятно, будет вредным: около 70 процентов этих мутаций имеют повреждающие эффекты, а остальные либо нейтральны, либо слабо полезны. [28]
Этот биологический процесс мутации представлен в популяционно-генетических моделях одним из двух способов: либо как детерминированное давление рекуррентной мутации на частоты аллелей, либо как источник вариаций. В детерминистской теории эволюция начинается с заранее определенного набора аллелей и продолжается сдвигами непрерывных частот, как будто популяция бесконечна. Возникновение мутаций у людей представлено «силой» или «давлением» мутации на популяционном уровне, т. е. силой бесчисленных событий мутации с масштабированной величиной u, применяемой к сдвигу частот f (A1) к f (A2). . Например, в классической баланса мутации и отбора модели [29] сила мутационного давления поднимает частоту аллели вверх, а отбор против ее вредных эффектов сдвигает частоту вниз, так что в равновесии достигается баланс, определяемый (в простейшем случае) соотношением f = u/s.
Эта концепция мутационного давления в основном полезна для рассмотрения последствий вредных мутаций, таких как мутационная нагрузка и ее влияние на эволюцию частоты мутаций. [30] Трансформация популяций под воздействием мутационного давления маловероятна. Холдейн [31] утверждал, что это потребует высокой частоты мутаций, не встречающей сопротивления со стороны отбора, и Кимура [32] пришли к еще более пессимистическому выводу, что даже это маловероятно, поскольку процесс займет слишком много времени (см. «Эволюция под давлением мутаций »).
Однако эволюция под воздействием мутаций возможна при некоторых обстоятельствах и уже давно рассматривается как возможная причина потери неиспользуемых признаков. [33] Например, пигменты перестают быть полезными, когда животные живут во тьме пещер и имеют тенденцию теряться. [34] Экспериментальный пример включает потерю споруляции в экспериментальных популяциях B. subtilis . Спорообразование — это сложный признак, кодируемый многими локусами, поэтому частота мутаций, приводящая к утрате этого признака, оценивалась как необычайно высокая. . [35] Потеря споруляции в этом случае может происходить путем рекуррентной мутации, не требуя отбора на потерю способности спорообразования. Когда нет отбора на потерю функции, скорость, с которой развивается потеря, больше зависит от частоты мутаций, чем от эффективного размера популяции . [36] что указывает на то, что это вызвано скорее мутацией, чем генетическим дрейфом.

Роль мутации как источника новизны отличается от этих классических моделей мутационного давления. Когда популяционно-генетические модели включают зависящий от скорости процесс внедрения или возникновения мутаций, т. е. процесс, который вводит новые аллели, включая нейтральные и полезные, тогда свойства мутации могут иметь более прямое влияние на скорость и направление эволюции. даже если скорость мутации очень низкая. [37] [38] То есть спектр мутаций может стать очень важным, особенно мутационные ошибки , предсказуемые различия в частоте возникновения разных типов мутаций, поскольку систематическая ошибка при внесении вариаций может внести систематические ошибки в ход эволюции. [39]
Мутация играет ключевую роль в других классических и новейших теориях, включая храповик Мюллера , субфункционализацию Эйгена , концепцию катастрофы ошибок Линча и гипотезу мутационной опасности .
Генетический дрейф
[ редактировать ]Генетический дрейф — это изменение частот аллелей , вызванное случайной выборкой . [40] То есть аллели потомства представляют собой случайную выборку аллелей родителей. [41] Генетический дрейф может привести к полному исчезновению вариантов генов и тем самым уменьшить генетическую изменчивость. В отличие от естественного отбора, который делает варианты генов более распространенными или менее распространенными в зависимости от их репродуктивного успеха, [42] изменения, вызванные генетическим дрейфом, не вызваны воздействием окружающей среды или адаптивными факторами и с одинаковой вероятностью сделают аллель более распространенной или менее распространенной.
Эффект генетического дрейфа сильнее для аллелей, присутствующих в небольшом количестве копий, чем когда аллель присутствует во многих копиях. Популяционная генетика генетического дрейфа описывается либо с помощью ветвящихся процессов , либо с помощью уравнения диффузии, описывающего изменения частоты аллелей. [43] Эти подходы обычно применяются к Райта-Фишера и Морана моделям популяционной генетики . Если предположить, что генетический дрейф является единственной эволюционной силой, действующей на аллель, то после t поколений во многих реплицируемых популяциях, начиная с частот аллелей p и q, дисперсия частот аллелей между этими популяциями равна

Рональд Фишер придерживался мнения, что дрейф генов играет в лучшем случае незначительную роль в эволюции, и эта точка зрения оставалась доминирующей в течение нескольких десятилетий. Ни одна перспектива популяционной генетики никогда не отводила генетическому дрейфу центральную роль сама по себе, но некоторые сделали генетический дрейф важным в сочетании с другой неизбирательной силой. Теория смещающегося баланса Сьюэлла Райта утверждала, что сочетание структуры популяции и генетического дрейфа имеет важное значение. Мотоо Кимуры утверждает Нейтральная теория молекулярной эволюции , что большинство генетических различий внутри популяций и между ними вызвано сочетанием нейтральных мутаций и генетического дрейфа. [45]
Роль генетического дрейфа посредством ошибки выборки в эволюции подверглась критике со стороны Джона Х. Гиллеспи. [46] и придет [47] которые утверждают, что отбор на связанных сайтах является более важной стохастической силой, выполняющей работу, традиционно приписываемую генетическому дрейфу, посредством ошибки выборки. Математические свойства генетического дрейфа отличаются от свойств генетического дрейфа. [48] Направление случайного изменения частоты аллелей автокоррелирует между поколениями. [40]
Поток генов
[ редактировать ]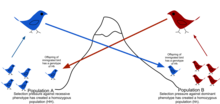
Из-за физических барьеров на пути миграции, а также ограниченной склонности особей к перемещению или распространению ( бродяга ) и склонности оставаться или возвращаться в родное место ( филопатрия ), естественные популяции редко скрещиваются между собой, как можно предположить в теоретических случайных моделях (филопатрия). панмикси ). [49] Обычно существует географический диапазон, в пределах которого люди более тесно связаны друг с другом, чем те, кто случайно выбран из общей популяции. Это описывается как степень генетической структурированности популяции. [50]

Генетическая структуризация может быть вызвана миграцией из-за исторического изменения климата , расширения ареала вида или текущей доступности среды обитания . Потоку генов препятствуют горные хребты, океаны и пустыни или даже созданные человеком структуры, такие как Великая Китайская стена , которая препятствует потоку растительных генов. [51]
Поток генов — это обмен генами между популяциями или видами, разрушающий структуру. Примеры потока генов внутри вида включают миграцию и последующее размножение организмов или обмен пыльцой . Перенос генов между видами включает образование гибридных организмов и горизонтальный перенос генов . Популяционно-генетические модели можно использовать для определения того, какие популяции демонстрируют значительную генетическую изоляцию друг от друга, и для реконструкции их истории. [52]
Изоляция популяции приводит к инбредной депрессии . Миграция в популяцию может привести к появлению новых генетических вариантов. [53] потенциально способствуя эволюционному спасению . Если значительная часть особей или гамет мигрирует, это также может изменить частоты аллелей, например, вызывая миграционную нагрузку . [54]
При наличии потока генов необходимы другие барьеры для гибридизации между двумя расходящимися популяциями скрещивающихся видов, чтобы популяции стали новыми видами .
Горизонтальный перенос генов
[ редактировать ]Горизонтальный перенос генов — это передача генетического материала от одного организма к другому организму, не являющемуся его потомком; это наиболее распространено среди прокариот . [55] В медицине это способствует распространению устойчивости к антибиотикам , поскольку, когда одна бактерия приобретает гены устойчивости, она может быстро передать их другим видам. [56] горизонтальный перенос генов от бактерий к эукариотам, таким как дрожжи Saccharomyces cerevisiae и жук адзуки Callosobruchus chinensis . Также мог иметь место [57] [58] Примером крупномасштабного переноса являются эукариотические бделлоидные коловратки , которые, по-видимому, получили ряд генов от бактерий, грибов и растений. [59] Вирусы также могут переносить ДНК между организмами, что позволяет передавать гены даже между биологическими доменами . [60] Крупномасштабный перенос генов произошел также между предками эукариотических клеток и прокариотами во время приобретения хлоропластов и митохондрий . [61]
Связь
[ редактировать ]Если все гены находятся в равновесии сцепления , эффект аллели в одном локусе может быть усреднен по всему генофонду в других локусах. В действительности один аллель часто обнаруживается в неравновесии по сцеплению с генами в других локусах, особенно с генами, расположенными поблизости в той же хромосоме. Рекомбинация разрушает это неравновесие по сцеплению слишком медленно, чтобы избежать генетического «автостопа» , когда частота аллеля в одном локусе возрастает до высокой частоты, потому что она связана с селекционируемой аллелью в соседнем локусе. Связь также замедляет скорость адаптации, даже в сексуальных популяциях. [62] [63] [64] Эффект неравновесия по сцеплению в замедлении скорости адаптивной эволюции возникает в результате сочетания эффекта Хилла-Робертсона (задержки в объединении полезных мутаций) и фонового отбора (задержки в отделении полезных мутаций от вредных автостопом ).
Сцепление является проблемой для популяционных генетических моделей, которые рассматривают один генный локус за раз. Однако его можно использовать как метод обнаружения действия естественного отбора посредством выборочных проверок .
В крайнем случае бесполой популяции сцепление является полным, и генетические уравнения популяции могут быть выведены и решены в терминах бегущей волны частот генотипов по простому ландшафту приспособленности . [65] Большинство микробов , например бактерий , бесполы. Популяционная генетика их адаптации имеет два контрастных режима. Когда произведение скорости полезных мутаций и размера популяции невелико, бесполые популяции следуют «последовательному режиму» динамики фиксации происхождения, при этом скорость адаптации сильно зависит от этого продукта. Когда продукт намного больше, бесполые популяции следуют режиму «параллельных мутаций», при котором скорость адаптации менее зависит от продукта, характеризующемуся клональной интерференцией и появлением новой полезной мутации до того, как последняя зафиксируется .
Приложения
[ редактировать ]Объяснение уровней генетической изменчивости
[ редактировать ]Нейтральная теория предсказывает, что уровень разнообразия нуклеотидов в популяции будет пропорционален произведению размера популяции и частоты нейтральных мутаций. Тот факт, что уровни генетического разнообразия различаются гораздо меньше, чем размеры популяций, известен как «парадокс изменчивости». [66] Хотя высокий уровень генетического разнообразия был одним из первоначальных аргументов в пользу нейтральной теории, парадокс изменчивости стал одним из самых сильных аргументов против нейтральной теории.
Ясно, что уровни генетического разнообразия сильно различаются внутри вида в зависимости от скорости локальной рекомбинации, как из-за генетического автостопа , так и из-за фонового отбора . Большинство современных решений парадокса вариаций требуют определенного уровня отбора на связанных сайтах. [67] Например, один анализ показывает, что более крупные популяции имеют более селективные зачистки, которые удаляют больше нейтрального генетического разнообразия. [68] Также может способствовать отрицательная корреляция между частотой мутаций и размером популяции. [69]
История жизни влияет на генетическое разнообразие больше, чем история популяции, например, у r-стратегов генетическое разнообразие больше. [67]
Обнаружение выбора
[ редактировать ]Модели популяционной генетики используются для определения того, какие гены подвергаются отбору. Одним из распространенных подходов является поиск областей с высоким неравновесием по сцеплению и низкой генетической вариативностью вдоль хромосомы, чтобы обнаружить недавние селективные проверки .
Второй распространенный подход — это тест Макдональда-Крейтмана , который сравнивает количество вариаций внутри вида ( полиморфизм ) с расхождением между видами (заменами) в двух типах сайтов; один считается нейтральным. Обычно сайты- синонимы считаются нейтральными. [70] Гены, подвергающиеся позитивному отбору, имеют избыток дивергентных сайтов по сравнению с полиморфными сайтами. Тест также можно использовать для получения общегеномной оценки доли замен, фиксируемых положительным отбором, α. [71] [72] Согласно нейтральной теории молекулярной эволюции , это число должно быть близко к нулю. Таким образом, высокие цифры были интерпретированы как общегеномная фальсификация нейтральной теории. [73]
Демографический вывод
[ редактировать ]Самый простой тест структуры популяции у диплоидных видов, размножающихся половым путем, - это проверить, соответствуют ли частоты генотипов пропорциям Харди-Вайнберга как функции частот аллелей. Например, в простейшем случае одного локуса с двумя аллелями, обозначенными A и a на частотах p и q , случайное спаривание предсказывает freq( AA ) = p 2 для АА гомозигот freq( aa ) = q 2 для гомозигот аа и freq( Aa ) = 2 pq для гетерозигот . При отсутствии структуры популяции пропорции Харди-Вайнберга достигаются за 1–2 поколения случайного спаривания. Чаще всего наблюдается избыток гомозигот, что свидетельствует о структуре популяции. Степень этого избытка можно определить количественно как коэффициент инбридинга F.
Особей можно сгруппировать в K субпопуляций. [74] [75] Затем степень структуры популяции можно рассчитать с помощью F ST , который является мерой доли генетической вариативности, которую можно объяснить структурой популяции. Затем генетическую структуру популяции можно связать с географической структурой и генетическую примесь обнаружить .
Теория слияния связывает генетическое разнообразие выборки с демографической историей популяции, из которой она была взята. Обычно он предполагает нейтральность , поэтому для такого анализа отбираются последовательности из более нейтрально развивающихся частей генома. Его можно использовать для вывода о взаимосвязях между видами ( филогенетика ), а также о структуре популяции, демографической истории (например, узких местах популяции , росте популяции ), биологическом расселении , динамике источника-поглотителя. [76] и интрогрессия внутри вида.
Другой подход к демографическому выводу основан на частотном спектре аллелей . [77]
Эволюция генетических систем
[ редактировать ]Предполагая, что существуют локусы, которые контролируют саму генетическую систему, создаются популяционные генетические модели для описания эволюции доминирования и других форм устойчивости , эволюции полового размножения и скорости рекомбинации, эволюции скорости мутаций , эволюции эволюционных конденсаторов. , эволюция дорогостоящих сигнальных признаков , эволюция старения и эволюция сотрудничества . Например, большинство мутаций вредны, поэтому оптимальная частота мутаций для вида может быть компромиссом между ущербом от высокой частоты вредных мутаций и метаболическими затратами на поддержание систем, снижающих скорость мутаций, таких как ферменты репарации ДНК. [78]
Одним из важных аспектов таких моделей является то, что отбор достаточно силен только для того, чтобы удалить вредные мутации и, следовательно, преодолеть мутационную предвзятость в сторону деградации, если коэффициент отбора s больше, чем величина, обратная эффективному размеру популяции . Это известно как барьер дрейфа и связано с почти нейтральной теорией молекулярной эволюции . Теория дрейфового барьера предсказывает, что виды с большими эффективными размерами популяций будут иметь весьма упорядоченные и эффективные генетические системы, в то время как виды с небольшими размерами популяций будут иметь раздутые и сложные геномы, содержащие, например, интроны и мобильные элементы . [79] Однако, как это ни парадоксально, виды с большими размерами популяций могут быть настолько толерантны к последствиям определенных типов ошибок, что у них возникает более высокий уровень ошибок, например, при транскрипции и трансляции , чем у небольших популяций. [80]
См. также
[ редактировать ]- Частотный спектр аллеля
- Ассортативное спаривание
- Балансирующий выбор
- Предвзятость при введении вариаций
- Коалесцентная теория
- Сохраняющая генетика
- Конструктивная нейтральная эволюция
- Выбор направления
- Распределение фитнес-эффектов (DFE)
- Гипотеза дрейфового барьера
- Эффективная численность населения
- Ошибка катастрофы
- Эволюционный ландшафт
- Фитнес-ландшафт
- Фиксация
- Эффект основателя
- Поток генов
- Генофонд
- Генетический дрейф
- Генетический автостоп
- Генетическая вариация
- Дилемма Холдейна
- Равновесие Харди-Вайнберга
- Идеализированное население
- Инбридинг
- Инбредная депрессия
- Модель бесконечных сайтов
- Внутригеномный конфликт
- Родственный отбор
- Неравновесие связей
- Тест Макдональда-Крейтмана
- Мейотический драйв
- трещотка Мюллера
- Баланс мутации и отбора
- Гипотеза мутационной опасности
- Естественный отбор
- Нейтральная теория молекулярной эволюции
- Панмиксия
- Аутбредная депрессия
- Полиморфизм
- Полигенная адаптация
- Численность населения
- Количественная генетика
- Процесс побега (побег по Фишерову)
- Выборочная развертка
- Эгоистичный генетический элемент
- Половой отбор
- Теория смещения баланса
- Субфункциональность
- Суперген (комплекс коадаптированных генов)
Ссылки
[ редактировать ]- ^ «Популяционная генетика – Последние исследования и новости» . www.nature.com . Проверено 29 января 2018 г.
- ^ Серведио, Мария Р .; Брандвейн, Янив; Дхоле, Сумит; Фитцпатрик, Кортни Л.; Гольдберг, Эмма Э.; Стерн, Кейтлин А.; Ван Клив, Джереми; Да, Д. Джастин (9 декабря 2014 г.). «Не просто теория — полезность математических моделей в эволюционной биологии» . ПЛОС Биология . 12 (12): e1002017. дои : 10.1371/journal.pbio.1002017 . ПМК 4260780 . ПМИД 25489940 .
- ^ Юэнс, У.Дж. (2004). Математическая популяционная генетика (2-е изд.). Нью-Йорк: Спрингер. ISBN 978-0-387-20191-7 .
- ^ Перейти обратно: а б с Боулер, Питер Дж. (2003). Эволюция: история идеи (3-е изд.). Беркли: Издательство Калифорнийского университета. стр. 325–339 . ISBN 978-0-520-23693-6 .
- ^ Перейти обратно: а б с Ларсон, Эдвард Дж. (2004). Эволюция: замечательная история научной теории (изд. Современной библиотеки). Нью-Йорк: Современная библиотека. стр. 221–243 . ISBN 978-0-679-64288-6 .
- ^ Битти, Джон (1986). «Синтез и синтетическая теория». Интеграция научных дисциплин . Наука и философия. Том. 2. Спрингер Нидерланды. стр. 125–135. дои : 10.1007/978-94-010-9435-1_7 . ISBN 9789024733422 .
- ^ Майр, Эрнст ; Провайн, Уильям Б., ред. (1998). Эволюционный синтез: взгляды на объединение биологии ([Новое изд.]. Ред.). Кембридж, Массачусетс: Издательство Гарвардского университета. стр. 295–298. ISBN 9780674272262 .
- ^ Перейти обратно: а б Провайн, ВБ (1988). «Прогресс в эволюции и смысл жизни». Эволюционный прогресс . Издательство Чикагского университета. стр. 49–79.
- ^ Перейти обратно: а б Провайн, Уильям Б. (1978). «Роль математических популяционных генетиков в эволюционном синтезе 1930-х и 1940-х годов». Исследования по истории биологии . 2 : 167–192. ПМИД 11610409 .
- ^ Форд, Э.Б. (1975) [1964]. Экологическая генетика (4-е изд.). Лондон: Чепмен и Холл. стр. 1 и далее.
- ^ Майр, Эрнст (1988). К новой философии биологии: наблюдения эволюциониста . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета . п. 402. ИСБН 978-0-674-89665-9 .
- ^ Майр, Эрнст ; Провайн, Уильям Б., ред. (1998). Эволюционный синтез: взгляды на объединение биологии ([Новое изд.]. Ред.). Кембридж, Массачусетс: Издательство Гарвардского университета. стр. 338–341. ISBN 9780674272262 .
- ^ Перейти обратно: а б с МакКэндлиш, Дэвид М.; Штольцфус, Арлин (сентябрь 2014 г.). «Моделирование эволюции с использованием вероятности фиксации: история и последствия». Ежеквартальный обзор биологии . 89 (3): 225–252. дои : 10.1086/677571 . ПМИД 25195318 . S2CID 19619966 .
- ^ Кроу, Джеймс Ф. (2010). «Райт и Фишер об инбридинге и случайном дрейфе» . Генетика . 184 (3): 609–611. дои : 10.1534/genetics.109.110023 . ISSN 0016-6731 . ПМЦ 2845331 . ПМИД 20332416 .
- ^ Касильяс, Соня; Барбадилья, Антонио (2017). «Молекулярная популяционная генетика» . Генетика . 205 (3): 1003–1035. doi : 10.1534/genetics.116.196493 . ПМК 5340319 . ПМИД 28270526 .
- ^ Холдейн, JBS (1927). «Математическая теория естественного и искусственного отбора, Часть V: Отбор и мутация». Математические труды Кембриджского философского общества . 23 (7): 838–844. Бибкод : 1927PCPS...23..838H . дои : 10.1017/S0305004100015644 . S2CID 86716613 .
- ^ Орр, ХА (2010). «Популяционная генетика полезных мутаций» . Философские труды Королевского общества B: Биологические науки . 365 (1544): 1195–1201. дои : 10.1098/rstb.2009.0282 . ПМК 2871816 . ПМИД 20308094 .
- ^ Хермиссон, Дж.; Пеннингс, PS (2005). «Мягкие зачистки: молекулярно-популяционная генетика адаптации к устойчивым генетическим вариациям» . Генетика . 169 (4): 2335–2352. doi : 10.1534/genetics.104.036947 . ПМЦ 1449620 . ПМИД 15716498 .
- ^ Гиллеспи, Джон (2004). Популяционная генетика: краткое руководство (2-е изд.). Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-8008-7 .
- ^ Мико, И. (2008). «Эпистаз: взаимодействие генов и эффекты фенотипа» . Природное образование . 1 (1): 197.
- ^ Бергер, Д.; Постма, Э. (13 октября 2014 г.). «Смещенные оценки эпистаза с убывающей отдачей? Пересмотр эмпирических данных» . Генетика . 198 (4): 1417–1420. дои : 10.1534/genetics.114.169870 . ПМЦ 4256761 . ПМИД 25313131 .
- ^ Куйос, Роджер Д.; Силандер, Олин К.; Бонхёффер, Себастьян (июнь 2007 г.). «Эпистаз между вредными мутациями и эволюцией рекомбинации». Тенденции в экологии и эволюции . 22 (6): 308–315. дои : 10.1016/j.tree.2007.02.014 . ПМИД 17337087 .
- ^ Кроу, Дж. Ф. (5 августа 1997 г.). «Высокий уровень спонтанных мутаций: риск для здоровья?» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (16): 8380–8386. Бибкод : 1997PNAS...94.8380C . дои : 10.1073/pnas.94.16.8380 . ПМК 33757 . ПМИД 9237985 .
- ^ Гастингс, П.Дж.; Лупски-младший; Розенберг, С.М.; Ира, Г. (2009). «Механизмы изменения числа копий гена» . Обзоры природы Генетика . 10 (8): 551–564. дои : 10.1038/nrg2593 . ПМК 2864001 . ПМИД 19597530 .
- ^ М., Лонг; Бетран, Э.; Торнтон, К.; Ван, В. (ноябрь 2003 г.). «Происхождение новых генов: взгляды молодых и старых». Нат. Преподобный Жене . 4 (11): 865–75. дои : 10.1038/nrg1204 . ПМИД 14634634 . S2CID 33999892 .
- ^ Лю, Н.; Окамура, К.; Тайлер, DM; Филлипс; Чанг; Лай (2008). «Эволюция и функциональное разнообразие генов микроРНК животных» . Клеточные исследования . 18 (10): 985–996. дои : 10.1038/cr.2008.278 . ПМК 2712117 . ПМИД 18711447 .
- ^ МакЛисахт, Аойф; Херст, Лоуренс Д. (25 июля 2016 г.). «Открытые вопросы в изучении генов de novo: что, как и почему» . Обзоры природы Генетика . 17 (9): 567–578. дои : 10.1038/nrg.2016.78 . ПМИД 27452112 . S2CID 6033249 .
- ^ Сойер, ЮАР; Парш, Дж.; Чжан, З.; Хартл, Д.Л. (2007). «Распространенность положительного отбора среди замен почти нейтральных аминокислот у дрозофилы» . Труды Национальной академии наук . 104 (16): 6504–6510. Бибкод : 2007PNAS..104.6504S . дои : 10.1073/pnas.0701572104 . ISSN 0027-8424 . ПМЦ 1871816 . ПМИД 17409186 .
- ^ Кроу, Джеймс Ф.; Кимура, Мотоо (1970). Введение в теорию популяционной генетики ([Перепечатка] под ред.). Нью-Джерси: Блэкберн Пресс. ISBN 9781932846126 .
- ^ Линч, Майкл (август 2010 г.). «Эволюция скорости мутаций» . Тенденции в генетике . 26 (8): 345–352. дои : 10.1016/j.tig.2010.05.003 . ПМЦ 2910838 . ПМИД 20594608 .
- ^ Дж. Б. С. Холдейн (1932). Причины эволюции . Лонгманс, Грин и Ко, Нью-Йорк.
- ^ М. Кимура (1980). «Среднее время до фиксации мутантного аллеля в конечной популяции под постоянным давлением мутаций: исследования аналитическими, численными методами и методами псевдовыборки» . Proc Natl Acad Sci США . 77 (1): 522–526. Бибкод : 1980PNAS...77..522K . дои : 10.1073/pnas.77.1.522 . ПМК 348304 . ПМИД 16592764 .
- ^ Холдейн, JBS (1933). «Роль повторяющихся мутаций в эволюции». Американский натуралист . 67 (708): 5–19. дои : 10.1086/280465 . JSTOR 2457127 . S2CID 84059440 .
- ^ Протас, Мередит; Конрад, М.; Гросс, Дж.Б.; Табин, К.; Боровский, Р. (2007). «Регрессивная эволюция мексиканской пещерной тетра Astyanax mexicanus» . Современная биология . 17 (5): 452–454. Бибкод : 2007CBio...17..452P . дои : 10.1016/j.cub.2007.01.051 . ПМК 2570642 . ПМИД 17306543 .
- ^ Х. Моган, Дж. Масел, К.В. Бирки-младший и У.Л. Николсон (2007). «Роль накопления мутаций и отбора в потере споруляции в экспериментальных популяциях Bacillus subtilis» . Генетика . 177 (2): 937–48. doi : 10.1534/genetics.107.075663 . ПМК 2034656 . ПМИД 17720926 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мазель, Дж . ; Кинг, О.Д.; Моэн, Х. (2007). «Потеря адаптивной пластичности в течение длительного периода застоя окружающей среды» . Американский натуралист . 169 (1): 38–46. дои : 10.1086/510212 . ПМК 1766558 . ПМИД 17206583 .
- ^ К. Гомес, Дж. Бертрам и Дж. Масель (2020). «Мутационная предвзятость может повлиять на адаптацию в больших бесполых популяциях, испытывающих клональное вмешательство» . Учеб. Р. Сок. Б. 287 (1937): 20201503. doi : 10.1098/rspb.2020.1503 . ПМЦ 7661309 . ПМИД 33081612 .
- ^ А. В. Кано, Х. Рожонова, А. Штольцфус, Д. М. МакКэндлиш и Дж. Л. Пейн (10 февраля 2022 г.). «Смещение мутаций формирует спектр адаптивных замен» . Proc Natl Acad Sci США . 119 (7). Бибкод : 2022PNAS..11919720C . дои : 10.1073/pnas.2119720119 . ПМЦ 8851560 . ПМИД 35145034 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Штольцфус, А.; Ямпольский, Л.Ю. (2009). «Восхождение на гору Вероятности: мутация как причина неслучайности в эволюции» . Журнал наследственности . 100 (5): 637–647. дои : 10.1093/jhered/esp048 . ПМИД 19625453 .
- ^ Перейти обратно: а б Мазель, Дж. (2011). «Генетический дрейф» . Современная биология . 21 (20): Р837–Р838. Бибкод : 2011CBio...21.R837M . дои : 10.1016/j.cub.2011.08.007 . ПМИД 22032182 .
- ^ Футуйма, Дуглас (1998). Эволюционная биология . Синауэр Ассошиэйтс . п. Глоссарий. ISBN 978-0-87893-189-7 .
- ^ Аверс, Шарлотта (1989). Процесс и закономерности в эволюции . Издательство Оксфордского университета.
- ^ Валь, Л.М. (2011). «Фиксация, когда N и s меняются: классические подходы дают новые элегантные результаты» . Генетика . 188 (4): 783–785. дои : 10.1534/genetics.111.131748 . ПМК 3176088 . ПМИД 21828279 .
- ^ Бартон, Николас Х.; Бриггс, Дерек Э.Г.; Эйзен, Джонатан А.; Гольдштейн, Дэвид Б.; Патель, Нипам Х. (2007). Эволюция . Лабораторный пресс Колд-Спринг-Харбор. п. 417. ИСБН 978-0-87969-684-9 .
- ^ Футуйма, Дуглас (1998). Эволюционная биология . Синауэр Ассошиэйтс . п. 320. ИСБН 978-0-87893-189-7 .
- ^ Гиллеспи, Дж. Х. (2000). «Генетический дрейф в бесконечной популяции: модель псевдоавтостопа» . Генетика . 155 (2): 909–919. дои : 10.1093/генетика/155.2.909 . ПМК 1461093 . ПМИД 10835409 .
- ^ Провайн, Уильям Б. Заблуждение о «случайном генетическом дрейфе» . CreateSpace.
- ^ Неер, Ричард А.; Шрайман, Борис И. (август 2011 г.). «Генетический проект и квазинейтральность в больших факультативно-сексуальных популяциях» . Генетика . 188 (4): 975–996. arXiv : 1108.1635 . дои : 10.1534/genetics.111.128876 . ISSN 0016-6731 . ПМК 3176096 . ПМИД 21625002 .
- ^ Бустон, премьер-министр; Пилкингтон, Дж. Г.; и др. (2007). «Состоят ли группы рыб-клоунов из близких родственников? Анализ изменений микросателлитной ДНК у Amphiprion percula ». Молекулярная экология . 12 (3): 733–742. дои : 10.1046/j.1365-294X.2003.01762.x . ПМИД 12675828 . S2CID 35546810 .
- ^ Репачи, В.; Стоу, Эй Джей; Бриско, Д.А. (2007). «Мелкомасштабная генетическая структура, сооснование и множественное спаривание австралийской аллодапиновой пчелы ( Ramphocinclus brachyurus )». Журнал зоологии . 270 (4): 687–691. дои : 10.1111/j.1469-7998.2006.00191.x .
- ^ Перейти обратно: а б Су, Х.; Цюй, Л.-Дж.; Он, К.; Чжан, З.; Ван, Дж; Чен, З.; Гу, Х. (2003). «Великая Китайская стена: физический барьер для потока генов?». Наследственность . 90 (3): 212–219. дои : 10.1038/sj.hdy.6800237 . ISSN 0018-067X . ПМИД 12634804 . S2CID 13367320 .
- ^ Гравий, С. (2012). «Модели популяционной генетики местного происхождения» . Генетика . 1202 (2): 607–619. arXiv : 1202.4811 . Бибкод : 2012arXiv1202.4811G . дои : 10.1534/genetics.112.139808 . ПМЦ 3374321 . ПМИД 22491189 .
- ^ Морджан, К.; Ризеберг, Л. (2004). «Как виды эволюционируют коллективно: последствия потока генов и отбора для распространения выгодных аллелей» . Молекулярная экология . 13 (6): 1341–56. Бибкод : 2004MolEc..13.1341M . дои : 10.1111/j.1365-294X.2004.02164.x . ПМК 2600545 . ПМИД 15140081 .
- ^ Больник, Дэниел И.; Носил, Патрик (сентябрь 2007 г.). «Естественный отбор в популяциях, подверженных миграционной нагрузке» . Эволюция . 61 (9): 2229–2243. дои : 10.1111/j.1558-5646.2007.00179.x . ПМИД 17767592 . S2CID 25685919 .
- ^ Баучер, Ян; Дуади, Кристоф Дж.; Папке, Р. Тейн; Уолш, Дэвид А.; Будро, Мэри Эллен Р.; Несбё, Камилла Л.; Кейс, Ребекка Дж.; Дулиттл, В. Форд (2003). «Боковой перенос генов и происхождение прокариотических групп». Ежегодный обзор генетики . 37 (1): 283–328. дои : 10.1146/annurev.genet.37.050503.084247 . ISSN 0066-4197 . ПМИД 14616063 .
- ^ Уолш, Т. (2006). «Комбинаторная генетическая эволюция мультирезистентности». Современное мнение в микробиологии . 9 (5): 476–82. дои : 10.1016/j.mib.2006.08.009 . ПМИД 16942901 .
- ^ Кондо, Н.; Никох, Н.; Иджичи, Н.; Шимада, М.; Фукацу, Т. (2002). «Фрагмент генома эндосимбионта Wolbachia, перенесенный на Х-хромосому насекомого-хозяина» . Труды Национальной академии наук . 99 (22): 14280–14285. Бибкод : 2002PNAS...9914280K . дои : 10.1073/pnas.222228199 . ISSN 0027-8424 . ПМЦ 137875 . ПМИД 12386340 .
- ^ Спрэг, Г. (1991). «Генетический обмен между королевствами». Текущее мнение в области генетики и развития . 1 (4): 530–533. дои : 10.1016/S0959-437X(05)80203-5 . ПМИД 1822285 .
- ^ Гладышев Е.А.; Мезельсон, М.; Архипова, ИР (2008). «Массовый горизонтальный перенос генов у бделлоидных коловраток» . Наука . 320 (5880): 1210–1213. Бибкод : 2008Sci...320.1210G . дои : 10.1126/science.1156407 . ISSN 0036-8075 . ПМИД 18511688 . S2CID 11862013 .
- ^ Бальдо, А.; МакКлюр, М. (1 сентября 1999 г.). «Эволюция и горизонтальный перенос генов, кодирующих дУТФазу, у вирусов и их хозяев» . Журнал вирусологии . 73 (9): 7710–7721. doi : 10.1128/JVI.73.9.7710-7721.1999 . ПМЦ 104298 . ПМИД 10438861 .
- ^ Пул, А.; Пенни, Д. (2007). «Оценка гипотез происхождения эукариот». Биоэссе . 29 (1): 74–84. doi : 10.1002/bies.20516 . ПМИД 17187354 .
- ^ Вайсман, Д.Б.; Халлачек, О. (15 января 2014 г.). «Скорость адаптации крупных половых популяций с линейными хромосомами» . Генетика . 196 (4): 1167–1183. дои : 10.1534/genetics.113.160705 . ПМЦ 3982688 . ПМИД 24429280 .
- ^ Вайсман, Дэниел Б.; Бартон, Николас Х.; Маквин, Гил (7 июня 2012 г.). «Пределы скорости адаптивного замещения в сексуальных популяциях» . ПЛОС Генетика . 8 (6): e1002740. дои : 10.1371/journal.pgen.1002740 . ПМЦ 3369949 . ПМИД 22685419 .
- ^ Неер, РА; Шрайман, Б.И.; Фишер, Д.С. (30 ноября 2009 г.). «Скорость адаптации больших сексуальных популяций» . Генетика . 184 (2): 467–481. arXiv : 1108.3464 . дои : 10.1534/genetics.109.109009 . ПМЦ 2828726 . ПМИД 19948891 .
- ^ Десаи, Майкл М.; Фишер, Дэниел С. (2007). «Баланс отбора полезных мутаций и влияние сцепления на положительный отбор» . Генетика . 176 (3): 1759–1798. doi : 10.1534/genetics.106.067678 . ЧВК 1931526 . ПМИД 17483432 .
- ^ Левонтин, RC (1973). Генетическая основа эволюционных изменений ([4-е издание] изд.). Нью-Йорк: Издательство Колумбийского университета. ISBN 978-0231033923 .
- ^ Перейти обратно: а б Эллегрен, Ганс; Галтье, Николя (6 июня 2016 г.). «Детерминанты генетического разнообразия» . Обзоры природы Генетика . 17 (7): 422–433. дои : 10.1038/nrg.2016.58 . ПМИД 27265362 . S2CID 23531428 .
- ^ Корбетт-Детиг, Рассел Б.; Хартл, Дэниел Л.; Сактон, Тимоти Б.; Бартон, Ник Х. (10 апреля 2015 г.). «Естественный отбор ограничивает нейтральное разнообразие широкого спектра видов» . ПЛОС Биология . 13 (4): e1002112. дои : 10.1371/journal.pbio.1002112 . ПМЦ 4393120 . ПМИД 25859758 .
- ^ Сун, В.; Акерман, М.С.; Миллер, Сан-Франциско; Доук, Т.Г.; Линч, М. (17 октября 2012 г.). «Гипотеза дрейфового барьера и эволюция скорости мутаций» (PDF) . Труды Национальной академии наук . 109 (45): 18488–18492. Бибкод : 2012PNAS..10918488S . дои : 10.1073/pnas.1216223109 . ПМЦ 3494944 . ПМИД 23077252 .
- ^ Чарльзуорт, Дж. Эйр-Уокер (2008). «Тест Макдональда-Крейтмана и слегка вредные мутации» . Молекулярная биология и эволюция . 25 (6): 1007–1015. дои : 10.1093/molbev/msn005 . ПМИД 18195052 .
- ^ Эйр-Уокер, А. (2006). «Геномная скорость адаптивной эволюции» (PDF) . Тенденции экологии и эволюции . 21 (10): 569–575. дои : 10.1016/j.tree.2006.06.015 . ПМИД 16820244 .
- ^ Смит, NGC; Эйр-Уокер, А. (2002). «Адаптивная эволюция белков у дрозофилы». Природа . 415 (6875): 1022–1024. Бибкод : 2002Natur.415.1022S . дои : 10.1038/4151022a . ПМИД 11875568 . S2CID 4426258 .
- ^ Хан, М.В. (2008). «К теории отбора молекулярной эволюции» . Эволюция . 62 (2): 255–265. дои : 10.1111/j.1558-5646.2007.00308.x . ПМИД 18302709 . S2CID 5986211 .
- ^ Причард, Дж. К.; Стивенс, М.; Доннелли, П. (июнь 2000 г.). «Вывод о структуре популяции с использованием данных мультилокусного генотипа» . Генетика . 155 (2): 945–959. дои : 10.1093/генетика/155.2.945 . ISSN 0016-6731 . ПМК 1461096 . ПМИД 10835412 .
- ^ Верити, Роберт; Николс, Ричард А. (август 2016 г.). «Оценка количества субпопуляций (K) в структурированных популяциях» . Генетика . 203 (4): 1827–1839. doi : 10.1534/genetics.115.180992 . ISSN 0016-6731 . ПМЦ 4981280 . ПМИД 27317680 .
- ^ Манлик, Оливер; Шабанн, Дельфин; Дэниел, Клэр; Бейдер, Ларс; Аллен, Саймон Дж.; Шервин, Уильям Б. (13 ноября 2018 г.). «Демография и генетика предполагают изменение динамики источника-поглотителя дельфинов, что имеет последствия для сохранения природы». Наука о морских млекопитающих . 35 (3): 732–759. дои : 10.1111/mms.12555 . S2CID 92108810 .
- ^ Гутенкунст, Райан Н.; Эрнандес, Райан Д.; Уильямсон, Скотт Х.; Бустаманте, Карлос Д.; Маквин, Гил (23 октября 2009 г.). «Вывод об общей демографической истории нескольких популяций на основе многомерных данных о частоте SNP» . ПЛОС Генетика . 5 (10): е1000695. arXiv : 0909.0925 . дои : 10.1371/journal.pgen.1000695 . ПМК 2760211 . ПМИД 19851460 .
- ^ Сниеговский, П. (2000). «Эволюция скорости мутаций: отделение причин от последствий». Биоэссе . 22 (12). Джерриш П.; Джонсон Т;. Шейвер А.: 1057–1066. doi : 10.1002/1521-1878(200012)22:12<1057::AID-BIES3>3.0.CO;2-W . ПМИД 11084621 . S2CID 36771934 .
- ^ Линч, Майкл; Конери, Джон С. (2003). «Истоки сложности генома». Наука . 302 (5649): 1401–1404. Бибкод : 2003Sci...302.1401L . CiteSeerX 10.1.1.135.974 . дои : 10.1126/science.1089370 . ПМИД 14631042 . S2CID 11246091 .
- ^ Рэджон, Э.; Мазель, Дж. (3 января 2011 г.). «Эволюция частоты молекулярных ошибок и последствия для эволюционности» . Труды Национальной академии наук . 108 (3): 1082–1087. Бибкод : 2011PNAS..108.1082R . дои : 10.1073/pnas.1012918108 . ПМК 3024668 . ПМИД 21199946 .
Внешние ссылки
[ редактировать ]- Учебные пособия по популяционной генетике (архивировано 23 января 2015 г.)
- Молекулярно-популяционная генетика
- База данных ALLele FREquency в Йельском университете
- EHSTRAFD.org - База данных частот аллелей STR человека Земли (архивировано 13 июля 2009 г.)
- История популяционной генетики
- Как отбор меняет генетический состав популяции , видео лекции Стивена Стернса ( Йельский университет )
- National Geographic : Атлас человеческого пути ( карты миграции людей на основе гаплогрупп )