Свободнорадикальное повреждение ДНК
Свободнорадикальное повреждение ДНК может произойти в результате воздействия ионизирующего излучения или радиомиметиков. [ 1 ] соединения. Повреждение ДНК в результате атаки свободных радикалов называется непрямым повреждением ДНК , поскольку образующиеся радикалы могут распространяться по всему телу и поражать другие органы. Злокачественная меланома может быть вызвана непрямым повреждением ДНК, поскольку она обнаруживается в частях тела, не подвергающихся воздействию солнечного света. ДНК уязвима для радикальной атаки из-за очень лабильных атомов водорода , которые можно оторвать, а также из-за преобладания двойных связей в основаниях ДНК , к которым могут легко присоединиться свободные радикалы . [ 2 ]
Повреждения вследствие радиационного воздействия
[ редактировать ]Радиолиз внутриклеточной воды под действием ионизирующего излучения приводит к образованию пероксидов , которые являются относительно стабильными предшественниками гидроксильных радикалов . 60–70% повреждений клеточной ДНК вызваны гидроксильными радикалами. [ 3 ] тем не менее, гидроксильные радикалы настолько реактивны, что могут диффундировать только на один или два диаметра молекулы, прежде чем вступить в реакцию с клеточными компонентами. , гидроксильные радикалы должны образовываться в непосредственной близости от нуклеиновых кислот Таким образом, чтобы вступить в реакцию . Радиолиз воды приводит к образованию пероксидов, которые могут действовать как диффундирующие латентные формы гидроксильных радикалов. Некоторые ионы металлов вблизи ДНК генерируют гидроксильные радикалы из перекиси. [ 4 ]
- Ч 2 О + hν → Ч 2 О + + и −
- Н2О + е − → Н 2 О −
- Н 2 О + → Ч + + ОН ·
- Н 2 О − → ОХ − + Ч ·
- 2 ОН · →H 2 O 2
Считается, что повреждение ДНК свободными радикалами вызывает мутации, которые могут привести к некоторым видам рака.
Реакция Фентона
[ редактировать ]Реакция Фентона приводит к образованию гидроксильных радикалов из перекиси водорода и катализатора железа (II). Железо(III) регенерируется по реакции Габера-Вейсса . Переходные металлы со свободным координационным центром способны восстанавливать пероксиды до гидроксильных радикалов. [ 1 ] Считается, что железо является металлом, ответственным за создание гидроксильных радикалов, поскольку оно существует в самой высокой концентрации среди всех переходных металлов в большинстве живых организмов. [ 5 ] Реакция Фентона возможна, поскольку переходные металлы могут существовать более чем в одной степени окисления, а их валентные электроны могут быть неспаренными, что позволяет им участвовать в одноэлектронных окислительно-восстановительных реакциях.
- Фе 2+ + H 2 O 2 → Fe 3+ + ОН · + ОН −
Создание гидроксильных радикалов в результате катализа железа (II) важно, поскольку железо (II) может быть координировано и, следовательно, находится в непосредственной близости от ДНК. Эта реакция позволяет перекиси водорода, образующейся в результате радиолиза воды, диффундировать к ядру и вступать в реакцию с железом (II) с образованием гидроксильных радикалов, которые, в свою очередь, реагируют с ДНК. Местоположение и связывание железа (II) с ДНК могут играть важную роль в определении субстрата и природы радикальной атаки на ДНК. Реакция Фентона генерирует два типа окислителей: тип I и тип II. Оксиданты I типа умеренно чувствительны к пероксидам и этанолу. [ 5 ] Окислители типа I и типа II преимущественно расщепляют определенные последовательности. [ 5 ]
Радикальная гидроксильная атака
[ редактировать ]
Гидроксильные радикалы могут атаковать остов и основания дезоксирибозы ДНК, потенциально вызывая множество повреждений , которые могут быть цитотоксическими или мутагенными . Клетки разработали сложные и эффективные механизмы восстановления для устранения повреждений. В случае атаки свободных радикалов на ДНК с вырезанием оснований используется механизм репарации . Реакции гидроксильного радикала с основной цепью сахара дезоксирибозы инициируются отрывом водорода от углерода дезоксирибозы, и основным последствием является окончательный разрыв цепи и высвобождение основания. Гидроксильный радикал реагирует с различными атомами водорода дезоксирибозы в порядке 5' H > 4' H > 3' H ≈ 2' H ≈ 1' H. Этот порядок реакционной способности аналогичен воздействию растворителя на водороды дезоксирибозы. [ 6 ]
Гидроксильные радикалы реагируют с основаниями ДНК путем присоединения к богатым электронами пи-связям. Эти пи-связи в основаниях расположены между C5-C6 пиримидинов и N7-C8 у пуринов . [ 7 ] При добавлении гидроксильного радикала может образоваться множество стабильных продуктов. В общем, радикальные гидроксильные атаки на основные фрагменты не вызывают изменения сахаров или разрывов цепей, за исключением случаев, когда модификации лабилизируют N-гликозильную связь, позволяя образовывать неосновные участки, которые подлежат бета-элиминированию.
Абазовые сайты
[ редактировать ]
Отщепление водорода от углерода 1'-дезоксирибозы гидроксильным радикалом приводит к образованию 1'-дезоксирибозильного радикала. Затем радикал может реагировать с молекулярным кислородом, создавая пероксильный радикал, который можно восстановить и дегидратировать с образованием 2'-дезоксирибонолактона и свободного основания. Дезоксирибонолактон обладает мутагенными свойствами и устойчив к репаративным ферментам. Таким образом создается абазовый сайт. [ 8 ]
Радикальное повреждение радиомиметическими соединениями
[ редактировать ]Радикальное повреждение ДНК может также произойти в результате взаимодействия ДНК с некоторыми природными продуктами, известными как радиомиметические соединения, молекулярными соединениями, которые влияют на ДНК аналогично радиационному воздействию. Радиомиметические соединения вызывают двухцепочечные разрывы ДНК посредством высокоспецифичных согласованных атак свободных радикалов на дезоксирибозные фрагменты в обеих цепях ДНК.
Общий механизм
[ редактировать ]Многие радиомиметические соединения представляют собой ендиины , которые подвергаются реакции циклизации Бергмана с образованием 1,4-дидегидробензольного бирадикала. Бирадикал 1,4-дидегидробензола обладает высокой реакционной способностью и отрывает водород от любого возможного донора водорода.
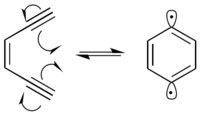
В присутствии ДНК 1,4-дидегидробензольный бирадикал отрывает атомы водорода от основной цепи сахара дезоксирибозы, преимущественно в положениях C-1', C-4' и C-5'. Отщепление водорода вызывает образование радикалов у прореагировавшего углерода. Углеродный радикал реагирует с молекулярным кислородом, что приводит к разрыву цепи ДНК посредством различных механизмов. [ 9 ] 1,4-Дидегидробензол способен позиционироваться таким образом, что может отрывать проксимальные атомы водорода от обеих цепей ДНК. [ 10 ] Это приводит к двухцепочечному разрыву ДНК, который, может привести к клеточному апоптозу если не исправить, .

Энедийны обычно подвергаются циклизации Бергмана при температуре, превышающей 200 ° C. Однако включение ендиина в 10-членный циклический углеводород делает реакцию более термодинамически выгодной за счет освобождения кольцевой деформации реагентов. Это позволяет циклизации Бергмана происходить при 37 ° C, биологической температуре человека. Было обнаружено, что молекулы, которые включают эндиины в эти более крупные кольцевые структуры, чрезвычайно цитотоксичны . [ 11 ]
Натуральные продукты
[ редактировать ]Энедиины присутствуют во многих сложных натуральных продуктах. Первоначально они были обнаружены в начале 1980-х годов во время поиска новых противораковых продуктов, вырабатываемых микроорганизмами. [ 10 ] Калихеамицин был одним из первых обнаруженных подобных продуктов и первоначально был обнаружен в образце почвы, взятом из Керрвилля, штат Техас. Эти соединения синтезируются бактериями в качестве защитных механизмов благодаря их способности расщеплять ДНК посредством образования 1,4-дидегидробензола из ендиинового компонента молекулы.

Калихеамицин и другие родственные соединения имеют несколько общих характеристик. Расширенные структуры, присоединенные к ендиину, позволяют этому соединению специфически связывать ДНК. [ 12 ] в большинстве случаев к малой бороздке двойной спирали. Кроме того, часть молекулы известна как «триггер», который при определенных физиологических условиях активирует эндиин, известный как «боевая часть», и генерируется 1,4-дидегидробензол.
С тех пор были идентифицированы три класса эндиинов: калихеамицин, динемицин и хромопротеинов продукты на основе .
Типы калихеамицина определяются метилтрисульфидной группой, которая участвует в запуске молекулы по следующему механизму. [ 10 ]

Калихеамицин и близкородственный ему эсперамицин использовались в качестве противораковых препаратов из-за их высокой токсичности и специфичности. [ 10 ]
Динемицин и его родственники характеризуются наличием антрахинонового и эндиинового ядра. Антрахиноновый компонент обеспечивает специфическое связывание ДНК на 3'-стороне пуриновых оснований посредством интеркаляции , сайта, отличного от калихеамицина. Его способность расщеплять ДНК значительно увеличивается в присутствии НАДФН и тиоловых соединений. [ 13 ] Это соединение также обнаружило известность в качестве противоопухолевого средства. [ 13 ]

Хромопротеиновые ендиины характеризуются нестабильным хромофорным ендиином, связанным с апопротеином .

Хромофор нереактивен при связывании с апопротеином. После высвобождения он реагирует с образованием 1,4-дидегидробензола и впоследствии расщепляет ДНК.
Противоопухолевая способность
[ редактировать ]Большинство эндиинов, включая перечисленные выше, использовались в качестве мощных противоопухолевых антибиотиков благодаря их способности эффективно расщеплять ДНК. Калихеамицин и эсперамицин являются двумя наиболее часто используемыми типами из-за их высокой специфичности при связывании с ДНК, что сводит к минимуму неблагоприятные побочные реакции. [ 12 ] Было показано, что они особенно полезны для лечения острого миелоидного лейкоза . [ 14 ]
Кроме того, калихеамицин способен расщеплять ДНК в низких концентрациях, оказываясь в 1000 раз более эффективным, чем адриамицин, в борьбе с некоторыми типами опухолей. [ 15 ]
Свободнорадикальный механизм лечения некоторых видов рака выходит за рамки эндиинов. Тирапазамин генерирует свободные радикалы в бескислородных условиях вместо триггерного механизма эндиина. Затем свободный радикал продолжает расщеплять ДНК аналогично 1,4-дидегидробензолу, чтобы лечить раковые клетки. В настоящее время он находится на стадии III испытаний.
Эволюция мейоза
[ редактировать ]Мейоз центральный признак полового размножения эукариот . — Было высказано предположение, что необходимость восстановления окислительного повреждения ДНК, вызванного окислительными свободными радикалами, является основной движущей силой эволюции мейоза. [ 16 ] [ 17 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б Барбусинский К (2009). «Реакция Фентона - споры по поводу химии». Экологическая химия и инженерия . 16 (3).
- ^ Гринберг М.М. (2016). «Реакционная способность радикалов нуклеиновых кислот» . Достижения физико-органической химии . 50 . Эльзевир: 119–202. дои : 10.1016/bs.apoc.2016.02.001 . ISBN 978-0-12-804716-3 . ПМЦ 5435387 . ПМИД 28529390 .
- ^ Уорд Дж. Ф. (1988). Повреждения ДНК, вызванные ионизирующим излучением в клетках млекопитающих: особенности, механизмы образования и возможности восстановления . Прогресс в исследованиях нуклеиновых кислот и молекулярной биологии. Том. 35. С. 95–125. дои : 10.1016/s0079-6603(08)60611-x . ISBN 9780125400350 . ПМИД 3065826 .
- ^ Хенле Э.С., Линн С. (август 1997 г.). «Формирование, предотвращение и восстановление повреждений ДНК, вызванных перекисью железа/водорода» . Журнал биологической химии . 272 (31): 19095–8. дои : 10.1074/jbc.272.31.19095 . ПМИД 9235895 . S2CID 11016259 .
- ^ Jump up to: а б с Погозельский В.К., Туллиус Т.Д. (май 1998 г.). «Окислительное расщепление цепи нуклеиновых кислот: пути, инициируемые отрывом водорода от фрагмента сахара». Химические обзоры . 98 (3): 1089–1108. дои : 10.1021/cr960437i . ПМИД 11848926 .
- ^ Баласубраманян Б., Погозельский В.К., Туллиус Т.Д. (август 1998 г.). «Разрыв цепи ДНК гидроксильным радикалом определяется доступными поверхностями атомов водорода основной цепи ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (17): 9738–43. Бибкод : 1998PNAS...95.9738B . дои : 10.1073/pnas.95.17.9738 . ПМК 21406 . ПМИД 9707545 .
- ^ Стенкен С. (1989). «Пуриновые основания, нуклезиды и нуклеотиды: окислительно-восстановительная химия водных растворов и реакции трансформации их катион-радикалов и аддуктов e- и OH». хим. Преподобный . 89 (3): 503–529. дои : 10.1021/cr00093a003 .
- ^ Ломм Дж., Констан Дж. Ф., Демёнинк М. (1999). «Базовая структура ДНК, реактивность и распознавание». Биополимеры . 52 (2): 65–83. doi : 10.1002/1097-0282(1999)52:2<65::aid-bip1>3.3.co;2-l . ПМИД 10898853 .
- ^ Повирк Л.Ф. (1996). «Повреждение ДНК и мутагенез радиомиметическими ДНК-расщепляющими агентами: блеомицином, неокарциностатином и другими ендиинами». Мутационные исследования/Фундаментальные и молекулярные механизмы мутагенеза . 355 (1–2): 71–89. дои : 10.1016/0027-5107(96)00023-1 . ПМИД 8781578 .
- ^ Jump up to: а б с д Крака Э., Кремер Д. (2000). «Компьютерный дизайн противораковых препаратов. Новая эндийная боеголовка». Дж. Ам. хим. Соц . 122 (34): 8245–8264. дои : 10.1021/ja001017k .
- ^ Чжэнь Ю.С., Мин XY, Ю Б., Отани Т., Сайто Х., Ямада Ю. (август 1989 г.). «Новый макромолекулярный противоопухолевый антибиотик С-1027. III. Противоопухолевая активность» . Журнал антибиотиков . 42 (8): 1294–8. дои : 10.7164/антибиотики.42.1294 . ПМИД 2759910 .
- ^ Jump up to: а б Эллестад, Джорджия (сентябрь 2011 г.). «Структурные и конформационные особенности, имеющие отношение к противоопухолевой активности калихеамицина γ 1I». Хиральность . 23 (8): 660–71. дои : 10.1002/чир.20990 . ПМИД 21800378 .
- ^ Jump up to: а б Сугиура Ю., Шираки Т., Кониси М., Оки Т. (май 1990 г.). «Интеркаляция ДНК и расщепление противоопухолевого антибиотика динемицина, содержащего антрациклиновые и эндииновые ядра» . Труды Национальной академии наук Соединенных Штатов Америки . 87 (10): 3831–5. Бибкод : 1990PNAS...87.3831S . дои : 10.1073/pnas.87.10.3831 . ПМК 53997 . ПМИД 2339123 .
- ^ Сиверс Э.Л., Аппельбаум Ф.Р., Спилбергер Р.Т., Форман С.Дж., Флауэрс Д., Смит Ф.О., Шеннон-Дорси К., Бергер М.С., Бернштейн И.Д. (июнь 1999 г.). «Селективная абляция острого миелолейкоза с использованием химиотерапии, нацеленной на антитела: исследование фазы I иммуноконъюгата калихеамицина против CD33» . Кровь . 93 (11): 3678–84. дои : 10.1182/blood.V93.11.3678 . ПМИД 10339474 .
- ^ Зейн Н., Синха А.М., МакГахрен В.Дж., Эллестад Г.А. (май 1988 г.). «Калихеамицин гамма 1I: противоопухолевый антибиотик, специфически расщепляющий двухцепочечный участок ДНК». Наука . 240 (4856): 1198–201. Бибкод : 1988Sci...240.1198Z . дои : 10.1126/science.3240341 . ПМИД 3240341 .
- ^ Хёрандл Э., Хадачек Ф (декабрь 2013 г.). «Гипотеза инициации окислительного повреждения мейоза» . Репродукция растения . 26 (4): 351–67. дои : 10.1007/s00497-013-0234-7 . ПМЦ 3825497 . ПМИД 23995700 .
- ^ Хёрандл Э., Спейер Д. (февраль 2018 г.). «Как кислород породил эукариотический пол» . Учебник по биологическим наукам . 285 (1872). дои : 10.1098/rspb.2017.2706 . ПМК 5829205 . ПМИД 29436502 .