Эфан
| Эфан | |
|---|---|

| |
| Ужасные птицы в Йены ботаническом саду | |
| Научная классификация | |
| Королевство: | Растения |
| Клэйд : | Трахеофиты |
| Клэйд : | покрытосеменные растения |
| Клэйд : | Однодольные |
| Клэйд : | Коммелиниды |
| Заказ: | Пальмоцветные |
| Семья: | Арековые |
| Подсемейство: | Арекоидеи |
| Племя: | Кокосовые |
| Род: | Эфан Уиллд. |
| Типовой вид | |
| Ужасные птицы | |
| Разнообразие | |
| Около 26 видов | |
| |
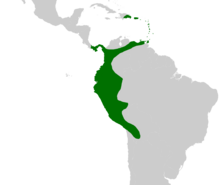
| Родное распространение Аифана | |
| Синонимы [ 1 ] | |
|
Мартинеция ( перепись Кунта , а не Руиса и Павона ) | |
Эфанес — род колючих , пальм произрастающий в тропических регионах Южной и Центральной Америки и Карибского бассейна . [ 2 ] около 26 В этом роде видов (см. Ниже), размеры которых варьируются от подлесковых кустарников с подземными стеблями до деревьев под пологом высотой до 20 метров (66 футов). Большинство из них имеют перисто- сложные листья (листья разделены на листочки, расположенные перьями, попарно вдоль центральной оси); у одного вида листья цельные . Стебли, листья, а иногда и плоды покрыты шипами. Растения цветут неоднократно в течение своей жизни и имеют отдельные мужские и женские цветки, хотя они растут вместе на одном соцветии . Хотя данные о опылителях ограничены, большинство видов, по-видимому, опыляются насекомыми. Плоды едят несколько птиц и млекопитающих, в том числе как минимум два вида амазонских попугаев .
Карл Людвиг Вильденов придумал название Aiphanes в 1801 году. До этого виды, принадлежащие к этому роду, были отнесены к Bactris или Caryota . К этому роду также применялось название Martinezia , и между 1847 и 1932 годами оно обычно использовалось вместо Aiphanes . Макс Буррет возродил название Aiphanes в 1932 году и заложил основу современной концепции рода. Айфан наиболее тесно связан с несколькими другими родами колючих пальм — Acrocomia , Astrocarium , Bactris и Desmoncus . Два вида широко выращиваются в качестве декоративных растений , а плоды, семена или сердцевина пальмы некоторых видов на протяжении тысячелетий употреблялись в пищу коренными народами Америки .
Описание
[ редактировать ]Aiphanes - это род колючих пальм, от деревьев под пологом высотой 20 метров (66 футов) до небольших кустарников с подземными стеблями, растущих в подлеске леса. [ 3 ] Его название сочетает в себе древнегреческое слово ai , что означает «всегда», и phaneros , что означает «очевидный», «видимый» или «заметный». [ 4 ] В своей монографии об этом роде 1996 года ботаники Финн Борхсениус и Родриго Берналь отметили, что «по иронии судьбы, виды Aiphanes обычно очень трудно заметить и найти в густой растительности, и, соответственно, они относятся к числу наиболее плохо собираемых неотропических пальм». [ 4 ]
Стебли
[ редактировать ]
Некоторые виды одноствольные, другие образуют многоствольные ( цеспитозные ) комки. В сочетании с различиями в размерах стеблей это приводит к разнообразию форм роста в этом роде: одиночные (одноствольные) пальмы, которые растут в подпологе леса, одиночные или каустные пальмы, растущие в подлеске леса, и бесстебельные пальмы, у которых нет стеблей. надземный стебель. [ 3 ]
Два вида характеризуются бесстебельным типом роста — A. acaulis и A. spicata . Два других вида — A. ulei и A. weberbaueri — встречаются как в бесстебельных популяциях, так и в популяциях, образующих надземные стебли. Некоторые виды представляют собой одноствольные подлесковые пальмы необычной формы роста. Aiphanes grandis и A. minima — это одноствольные пальмы, высота которых превышает 10 метров (33 фута), а остальные представляют собой многоствольные подлесковые виды. Многоствольные пальмы варьируются от растений с одним главным стеблем и несколькими прикорневыми присосками до цестовидных комков из 20 плотно расположенных стеблей. В пределах одного вида могут существовать различные формы роста, и это, по-видимому, зависит от среды обитания и условий окружающей среды. [ 3 ]
Листья
[ редактировать ]Листья видов Aiphanes обычно перисто разделены: ряды листочков выходят по обе стороны от оси листа в виде перьев или папоротников. Единственным исключением является A. macroloba , у которого листья цельные . Обычно они расположены спирально, но у некоторых видов листья расположены раздельно , что является нормальным для сеянцев пальм, но редко встречается у взрослых особей. Основания старых листьев аккуратно отделяются от стебля, за исключением A. hirsuta subsp. Fosteriorum , у которого часто старые основания листьев прикрепляются к новым частям стебля. [ 5 ]
Листья колючие, но степень колючести варьируется как внутри вида, так и между видами. Листовые влагалища всегда густо колючие, но к концам листьев шипы обычно становятся меньше и реже. [ 5 ]
Шипы
[ редактировать ]
Шипы характерны для Aiphanes и других представителей подтрибы Bactridinae . Они встречаются почти повсеместно на растениях и особенно хорошо развиты на стебле, основаниях листьев и цветоносе . У Aiphanes шипы образуются из внешних тканей растения и не возникают в результате модификации других органов растения. Их длина варьируется от менее 1 миллиметра (0,04 дюйма) до более 25 сантиметров (9,8 дюйма). [ 6 ]
Цветы
[ редактировать ]Виды Aiphanes являются плеонантическими (они цветут неоднократно в течение своей жизни) и однодомными , что означает, что есть отдельные мужские и женские цветки, но отдельные растения несут оба типа цветов. У Aiphanes мужские и женские цветки располагаются вместе на одном соцветии . цветет только одно соцветие Обычно в каждом узле , хотя A. gelatinosa часто цветет группами по три в одном узле. Соцветие обычно состоит из главной оси, состоящей из цветоноса и рахиса . Рахис несет рахиллы, представляющие собой более мелкие ветви, несущие цветы, а цветонос представляет собой основной стебель, соединяющий рахис со стеблем растения. У некоторых видов наблюдается ветвление второго порядка — сами рахиллы ветвистые, на этих ветвях цветут. [ 7 ]
Цветки обычно цветут группами по три — один женский цветок вместе с двумя мужскими цветками. У некоторых видов зарегистрированы группы из четырех цветков (два мужских и два женских). На дальнем конце соцветия, вдали от оси дерева, пары мужских цветков заменяют триады мужских и женских цветков. Окраска цветков малоизвестна. Записывать его необходимо с живых растений, так как сохранившиеся цветы со временем теряют окраску, а записи об этих видах в дикой природе неполны. Мужские цветки, как правило, делятся на две группы: с кремовыми или желтыми цветками и с некоторым количеством фиолетового цвета в цветках. Женские цветки еще менее известны, чем мужские. [ 7 ]
Пыльцевые зерна обычно имеют форму от сферической до эллипсоидной, иногда треугольной, примерно от 20 до 30 микрометров по длинной оси и от 20 до 30 микрометров в диаметре. Обычно они однобороздные , меридионосулькатные , реже трихотомосулькатные . [ 8 ] Борозда представляет собой бороздку, идущую вдоль поверхности пыльцевого зерна и обычно являющуюся местом, где происходит опыление. Пыльца однобороздчатая имеет одну бороздку, идущую вдоль полюса пыльцевого зерна. Меридионосульчатая пыльца имеет бороздку, идущую по экватору пыльцевого зерна. [ 9 ] Пыльца трихотомосулькатной, напротив, имеет три бороздки. [ 10 ] Внешний слой пыльцы в большей или меньшей степени покрыт гребнями, шипами или бородавками. Эта «скульптура», как правило, более выражена у видов, опыляемых мухами, и менее выражена у тех, которые опыляются жуками или пчелами. [ 8 ]
Фрукты
[ редактировать ]Плод вида Aiphanes обычно представляет собой красную шаровидную односемянную костянку . Тонкая кожица (или эпикарпий ), которая может быть как гладкой, так и колючей, покрывает мясистый мезокарпий , который обычно имеет оранжевый и сладкий цвет. Мезокарпий A. horrida имеет одно из самых высоких показателей содержания каротина среди всех растительных продуктов, а также богат белком . Эндокарпий , в котором заключено семя, коричневый или черный и в зрелом состоянии очень твердый. [ 11 ] Семена светло-коричневые, с тонкой семенной оболочкой (или семенником) и белым эндоспермом , сладким и по вкусу напоминающим кокос . [ 11 ]
Кариотип
[ редактировать ]Опубликованные данные о количестве хромосом существуют для двух видов: Aiphanes minima и A. horrida ; Количество гаплоидных хромосом варьируется от 15 до 18. Борхсениус и Бернал сообщают, что трудно получить точное количество хромосом на пальмах и что различия в количестве хромосом могут отражать эти трудности. [ 12 ]
Таксономия
[ редактировать ]| Родство между представителями подтрибы Bactridinae , основанное на пластидной ДНК. филогении [ 13 ] * Aiphanes aculeata — ботанический синоним A. horrida. |

Аифан помещен в подсемейство Arecoideae , трибу Cocoseae и подтрибу Bactridinae вместе с родами Desmoncus , Bactris , Acrocomia и Astrocaryum . [ 14 ]
В своей ревизии рода в 1932 году немецкий ботаник Макс Буррет выделил 32 вида. Семнадцать из них были новыми видами, в основном основанными на коллекциях, собранных немецким ботаником Вильгельмом Кальбрейером в северной Колумбии между 1877 и 1881 годами. Работая с очень узкой концепцией вида и не будучи знаком с вариациями, присутствующими в природных популяциях, Беррет поместил почти каждый экземпляр в отдельный вид. Бомбардировка Берлинского гербария во время Второй мировой войны уничтожила единственные известные коллекции 13 из этих 32 видов, что еще больше усложнило ситуацию. [ 15 ]
Международный кодекс ботанической номенклатуры требует, чтобы каждый вид был представлен типовой коллекцией . [ 16 ] В результате уничтожения коллекций типов Беррета многие виды стали известны только по его оригинальным описаниям, в которых обычно отсутствовали иллюстрации. Другие образцы (называемые неотипами) были предназначены для замены их Родриго Берналем и его коллегами в 1989 году. [ 17 ] или Борхсениусом и Берналом в их монографии об этом роде 1996 года. [ 18 ] Бернал и его коллеги попытались проследить путешествия Калбрейера по северной Колумбии и собрать образцы как можно ближе к месту расположения первоначальных коллекций. [ 17 ]
Беррет разделил Aiphanes на два подрода: Brachyanthera и Macroanthera . Одиннадцать видов были помещены в Macroantera , а остальные были помещены в Brachyantera . [ 15 ] В своей монографии 1996 года Борхсениус и Бернал поставили под сомнение применимость этих подродов. Они признали, что если Macroanthera сократить до трех видов ( A. horrida , A. eggersii и A. minima ), она могла бы сформировать жизнеспособную группу, но это сделало бы Brachyanthera чрезмерно гетерогенной. Следовательно, они отказались от использования Бёрретом подродов. [ 19 ]
За три десятилетия после определения рода Берретом было описано еще 15 видов, в результате чего общее количество видов достигло 47. [ 15 ] Борхсениус и Бернал определили, что многие из этих названий являются синонимами , хотя американский ботаник Джордж Проктор не согласился с их решением объединить A. acanthophylla с A. minima . [ 20 ] Борхсениус и Бернал также описали один новый вид, Aiphanes spicata , в результате чего общее число признанных видов достигло 22. [ 3 ] В двух случаях уничтожение единственных известных коллекций лишило возможности быть абсолютно уверенным в том, что имя является синонимом. [ 21 ] Текущий Всемирный контрольный список избранных семейств растений , который ведет Рафаэль Говертс в Королевском ботаническом саду Кью , признает 26 видов, включая четыре вида, описанные после публикации монографии Борхсениуса и Бернала. [ 22 ]
Беррет разделил Aiphanes на два подрода: Brachyanthera и Macroanthera . Одиннадцать видов были помещены в Macroantera , а остальные — в Brachyantera . [ 15 ] В своей монографии 1996 года Борхсениус и Бернал поставили под сомнение применимость этих подродов. Они признали, что если бы Macroanthera сократилась до трех видов ( A. horrida , A. eggersii и A. minima ), она могла бы сформировать жизнеспособную группу, но это сделало бы Brachyanthera чрезмерно гетерогенной. Следовательно, они отказались от использования Бёрретом подродов. [ 19 ]
История
[ редактировать ]Самое раннее ботаническое описание вида этого рода было сделано французским ботаником Шарлем Плюмье , который описал два вида на основе своих визитов в Вест-Индию между 1689 и 1695 годами. Оба вида Плюмье теперь считаются Aiphanes minima . Тот же вид был описан голландским ботаником Николаусом Йозефом фон Жакеном в 1763 году. Испанский ботаник Хосе Селестино Мутис представил подробное описание A. lindeniana и иллюстрации этого вида и того, что считается A. horrida, в 1779 году. [ 4 ]
В 1791 году Йозеф Гертнер включил вид Aiphanes в свой De Fructibus et Seminibus Plantarum , назвав его Bactris minima . Это самое старое действительно опубликованное название любого представителя рода. Название Aiphanes было придумано немецким ботаником Карлом Людвигом Вильденовым в 1801 году. Он описал единственный вид, A. aculeata , в 1806 году. [ 4 ]
Жакин использовал название Caryota horrida для описания растения, которое принадлежало к тому же виду (и, возможно, было той же особью), что описано Уилденоу. Борхсениус и Берналь ссылаются на дату публикации описания Жакена в 1809 году, в которой приоритет отдается имени Вильденоу. [ 4 ] Однако в более позднем Всемирном контрольном списке (2006 г.) указана дата публикации описания Жакена 1801 год, что делает A. horrida правильным названием этого вида. [ 23 ]
В 1816 году Александр фон Гумбольдт , Эме Бонплан и Карл Сигизмунд Кунт описали Martinezia caryotifolia , добавив еще одно название в список синонимов A. horrida . Поскольку первоначальные диагностические признаки Martinezia не подходили ни к одному из существующих видов, Кунт переопределил их, чтобы они соответствовали M. caryotifolia . Следовательно, Martinezia пришла на смену Aiphanes , и последнее название редко использовалось в период с 1847 по 1932 год. В 1857 году Герман Карстен создал новый род Marara для включения двух колумбийских видов, M. bicuspidata (позже выяснилось, что это синоним A. horrida). ) и M. erinacea (ныне A. erinacea ). Герман Вендланд попытался воскресить Эфана в 1878 году, объединив с ним Мартинецию и Марару , но его предложение было проигнорировано. В 1901 оратор Ф. Кук создал два новых рода — Curima , к которому он отнес A. minima , и Tilmia , к которому отнес A. horrida . [ 24 ] В 1932 году, после публикации вида в Мартинеции , Беррет изменил свое мнение об этом роде и сделал его синонимом Aiphanes . Это привело к нынешнему разграничению рода. [ 15 ]
Разновидность
[ редактировать ]Виды, включенные во Всемирный контрольный список избранных семейств растений : [ 2 ]
- Aiphanes acanthophylla (Mart.) Burret - Пуэрто-Рико, Доминиканская Республика.
- Aiphanes acaulis Галеано и Р.Берналь – Колумбия
- Аргос Аифан Р.Берналь, Борхс. & Хоулс-Гомес – Колумбия [ 25 ]
- Aiphanes bicornis Cerón и Р.Берналь – Эквадор
- Aiphanes buenaventurae Р.Берналь и Борхс. – Валье дель Каука в Колумбии
- Aiphanes chiribogensis Borchs. и Балслев – Эквадор
- Aiphanes deltoidea Burret - Колумбия, Перу, северо-запад Бразилии.
- Айфан Дукей Буррет – Колумбия
- Aiphanes eggersii Burret - Эквадор, Перу.
- Aiphanes erinacea (H.Karst.) H.Wendl. – Колумбия, Эквадор
- Aiphanes gelatinosa HEMoore - Колумбия, Эквадор.
- Aiphanes graminifolia Galeano & R.Bernal – Колумбия
- Айфан Грандис Борхс. и Балслев – Эквадор
- Aiphanes hirsuta Burret – Колумбия, Эквадор, Панама, Коста-Рика.
- Aiphanes horrida (Jacq.) Burret - Тринидад, Колумбия, Венесуэла, Перу, северо-запад Бразилии, Боливия.
- Aiphanes leiostachys Burret - Антиокия в Колумбии.
- Aiphanes lindeniana (H.Wendl.) H.Wendl. -Колумбия
- Aiphanes Linearis Burret - Антиокия и Валле-дель-Каука в Колумбии.
- Aiphanes macroloba Burret – Колумбия, Эквадор.
- Aiphanes minima (Gaertn.) Burret - Сент-Люсия, Барбадос
- Мультиплекс «Айфан» Р.Берналь и Борхс. – Валье-дель-Каука в Колумбии
- Aiphanes parvifolia Burret - Колумбия.
- Aiphanes pilaris Р.Берналь – Колумбия
- Aiphanes simplex Burret - Колумбия.
- Aiphanes spicata Borchs. и Р.Берналь – Перу
- Aiphanes stergiosii S.M.Niño - Штат Португеза на западе Венесуэлы.
- Aiphanes tricuspidata Borchs., M.Ruíz & Bernal – Колумбия, Эквадор
- Aiphanes ulei (Даммер) Буррет - Колумбия, Эквадор, Перу, северо-запад Бразилии.
- Аифан бородавчатый Борхс. и Балслев – Эквадор
- Aiphanes weberbaueri Burret – Эквадор, Перу
Распространение и статус
[ редактировать ]Род Aiphanes распространен от Эспаньолы ( Доминиканская Республика ) и Панамы на севере до Тринидада и Тобаго на востоке, через Колумбию и вниз по Андам до Боливии . В Бразилии встречается только вдоль границы с Перу . Aiphanes обитает преимущественно в Южной Америке: один вид ( A. hirsuta ) встречается в Панаме, а два других ( A. horrida и A. minima ) встречаются в Карибском бассейне. Aiphanes minima , эндемик островного Карибского бассейна, является единственным видом, отсутствующим на материковой части Южной Америки. Хотя A. horrida была зарегистрирована в Гайане и южной Венесуэле, эти сообщения не были подтверждены гербарными ваучерами. [ 26 ]
Aiphanes horrida — наиболее широко распространенный вид. Он простирается от Тринидада до Боливии, но отсутствует в Эквадоре и северном Перу. Другие виды имеют более узкие ареалы: один центр разнообразия находится в западной Колумбии и Эквадоре, а другой второстепенный — на северо-востоке Перу. [ 26 ] 2006 г. В Красный список МСОП включены три вида, которым угрожает разрушение среды обитания: A. grandis , [ 27 ] А. лейостахис [ 28 ] и бородавчатая [ 29 ] — и трое других, считающихся уязвимыми для той же угрозы — A. chiribogensis , [ 30 ] А. Дюк [ 31 ] и А. линдениана . [ 32 ] Родриго Берналь и Глория Галеано расширили этот список в обзоре состояния колумбийских пальм в 2005 году. Они включили два вида в список находящихся под угрозой исчезновения — A. graminifolia , вид, который был впервые описан в 2002 году, и A. leiostachys (который был классифицирован как находящийся под угрозой исчезновения в Красном списке МСОП). Они классифицировали два вида как находящиеся под угрозой исчезновения — A. acaulis и A. parvifolia — и два вида как уязвимые — A. gelatinosa и A. pilaris . Они также классифицировали шесть видов как находящиеся под угрозой исчезновения — A. erinacea , A. hirsuta , A. lindeniana (уязвимые согласно Красному списку МСОП), A. Linearis , A. macroloba и A. simplex . [ 33 ] Угрозы для этих видов не были перечислены, но Йенс-Кристиан Свеннинг сообщил, что A. erinacea находится под угрозой вырубки, учитывая ее ограниченное распространение и плохую способность к регенерации в нарушенных лесах. [ 34 ] В дополнение к этому, A. deltoidea , которая широко распространена в западных тропических лесах Амазонки , присутствует в такой низкой плотности, что Фрэнсис Кан и Фарана Мусса в 1994 году классифицировали ее как редкий вид. [ 35 ]
Среда обитания и экология
[ редактировать ]
Виды Aiphanes – пальмы подлеска и подполога леса. Наиболее широко распространенный вид, A. horrida , встречается как в тропических сухих лесах , так и в более влажных типах леса, но в его распространении имеется пробел, совпадающий с наиболее влажными лесами верхней части бассейна Амазонки . Два других вида, A. minima и A. eggersii , также встречаются в более засушливых условиях; A. eggersii встречается в районах, где выпадает всего 500 мм (20 дюймов) осадков в год. Остальные виды встречаются в горных лесах на больших высотах или во влажных, часто очень влажных, низинных лесах, включая районы, где ежегодно выпадает до 9000 мм (350 дюймов) осадков. [ 26 ]
Записи о посещениях опылителями существуют лишь для нескольких видов, но большинство из них позволяют предположить, что эти виды опыляются насекомыми. Цветки A. chiribogensis выделяют небольшое количество нектара , но не имеют запаха. плодовые мухи ( Drosophilidae ), грибные комары ( Mycetophilidae , Sciaridae ), мошки ( Cecidomyiidae , Ceratopogonidae ) и микроножки ( Lepidoptera При посещении этих цветов были зарегистрированы ), а вот пчелы и журчалки — нет. Aiphanes eggersii Считалось, что дрозофилами ( Drosophilidae ), журчалками (Syrphidae), мокрецами ( Ceratopogonidae ) и листоедами опыляется пчелами и, возможно, ветром. Зарегистрировано посещение цветков A. erinacea (Chrysomelidae) , а пчелами — нет. Сообщается, что Aiphanes horrida опыляется ветром, пчелами ( Meliponidae ), долгоносиками ( Curculionidae ) и клопами ( Hemiptera ). На цветках A. simplex наблюдались мухи и долгоносики . [ 36 ]
Плоды A. horrida богаты витаминами и энергией и могут быть съедены многими животными. масличные птицы Сообщается, что поедают его плоды и разносят семена. Сообщается, что белки также едят плоды, несмотря на колючую природу дерева. [ 36 ] Плоды, цветы и семена A. minima поедаются уязвимой амазонкой Сент-Винсента ( Amazona guildingii ). [ 37 ] а также считается потенциально важным пищевым видом для находящейся под угрозой исчезновения пуэрто-риканской амазонки ( Amazona vittata ). [ 38 ]
Некоторые виды демонстрируют скопленное распространение. Ограничение расселения было использовано для объяснения скопления взрослых особей и ограниченного набора сеянцев как у A. erinacea в Эквадоре, так и у A. erinacea. [ 39 ] и A. minima в Пуэрто-Рико. [ 38 ] Аналогично, редкость A. lindeniana и A. simplex в колумбийских лесах может быть связана с ограниченным производством семян и ограниченной эффективностью распространения семян плодоядными птицами и млекопитающими. [ 40 ]
Использование
[ редактировать ]Виды Aiphanes имеют долгую историю использования человеком. Остатки обуглившихся семян, предположительно принадлежащих A. horrida, были обнаружены на археологических раскопках в Колумбии и датируются примерно 2800 годом до нашей эры ; [ 41 ] семена этого вида до сих пор потребляются и продаются на местных рынках. [ 42 ] Aiphanes horrida также широко выращивается как декоративное растение, как и A. minima . [ 43 ] [ 44 ] Плоды или семена A. deltoidea , [ 45 ] А. Эггерсии , [ 46 ] А. линеарис [ 47 ] и А. минимума [ 48 ] все потребляются локально. Пальмовое сердце A. macroloba потребляется народом Коайкер на северо-западе Южной Америки. [ 49 ] Айфанол , соединение, выделенное из A. horrida , продемонстрировало значительную ингибирующую активность в отношении циклооксигеназ ; [ 50 ] ингибирование этих ферментов может облегчить симптомы воспаления и боли. [ 51 ]
Примечания
[ редактировать ]- ^ Борхсениус и Бернал (1996), стр. 33–34
- ^ Jump up to: а б Контрольный список избранных семейств растений Kew World
- ^ Jump up to: а б с д Борхсениус и Бернал (1996), с. 4
- ^ Jump up to: а б с д и Борхсениус и Бернал (1996), с. 2
- ^ Jump up to: а б Борхсениус и Бернал (1996), стр. 8–9
- ^ Борхсениус и Бернал (1996), стр. 6–8
- ^ Jump up to: а б Борхсениус и Бернал (1996), стр. 11–14
- ^ Jump up to: а б Борхсениус и Бернал (1996), стр. 16–22
- ^ Хоэн, Питер (1999). «Словарь терминологии пыльцы и спор» . Лаборатория палеоботаники и палинологии Мичиганского технологического университета. Архивировано из оригинала 18 мая 2010 года . Проверено 17 марта 2010 г.
- ^ Надот, С.; А. Форчиони; Л. Пенет; Ж. Саннье; А. Рессейр (2006). «Связь между ранним развитием пыльцы и рисунком апертуры у однодольных». Протоплазма . 228 (1–3): 55–64. дои : 10.1007/s00709-006-0164-4 . ПМИД 16937055 . S2CID 28497230 .
- ^ Jump up to: а б Борхсениус и Бернал (1996), стр. 14–15
- ^ Борхсениус и Бернал (1996), с. 26
- ^ Асмуссен, Конни Б.; Джон Дрансфилд; Винни Дейкманн; Андерс С. Барфод; Жан-Кристоф Пино; Уильям Дж. Бейкер (2006). «Новая классификация подсемейства пальмовых (Arecaceae): данные филогении пластидной ДНК» . Ботанический журнал Линнеевского общества . 151 (1): 15–38. дои : 10.1111/j.1095-8339.2006.00521.x .
- ^ Дрансфилд, Джон; Натали В. Уль; Конни Б. Асмуссен; Уильям Дж. Бейкер; Мэдлин М. Харли; Карл Э. Льюис (2005). «Новая филогенетическая классификация семейства пальмовых Arecaceae». Бюллетень Кью . 60 (4). Королевский ботанический сад, Кью: 559–69. JSTOR 25070242 .
- ^ Jump up to: а б с д и Борхсениус и Бернал (1996), с. 3
- ^ «Статья 7» . Международный кодекс ботанической номенклатуры (Венский кодекс) . Проверено 20 марта 2010 г.
- ^ Jump up to: а б Берналь, Родриго Г.; Глория Галеано-Гарсес; Эндрю Хендерсон (1989). «Неотипизация колумбийских пальм, собранных В. Калбрейером». Таксон . 38 (1). Международная ассоциация таксономии растений (IAPT): 98–107. дои : 10.2307/1220905 . JSTOR 1220905 .
- ^ Борхсениус и Бернал (1996)
- ^ Jump up to: а б Борхсениус и Бернал (1996), с. 33
- ^ Джордж Р. Проктор, в Acevedo-Rodríguez & Strong (2005), стр. 138–139.
- ^ Борхсениус и Бернал (1996), с. 87
- ^ Говертс, Р. ; Дж. Хендерсон; СФ Зона; Д.Р. Ходел; А. Хендерсон (2006). «В поисках Эфана » . Всемирный контрольный список арековых . Попечительский совет Королевского ботанического сада Кью . Проверено 5 января 2010 г.
- ^ « Эйфан Хоррида » . Королевский ботанический сад, Кью : Всемирный контрольный список избранных семейств растений . Проверено 4 января 2010 г.
- ^ Борхсениус и Бернал (1996), стр. 2–3.
- ^ « Аифан Аргос » . Растения мира онлайн . Королевский ботанический сад, Кью . Проверено 2 марта 2023 г.
- ^ Jump up to: а б с Борхсениус и Бернал (1996), стр. 26–30
- ^ Монтуфар, Р.; Питман, Н. (2003). « Айфан Грандис » . Красный список исчезающих видов МСОП . 2003 : e.T43757A10826566. doi : 10.2305/IUCN.UK.2003.RLTS.T43757A10826566.en .
- ^ Бернал, Р. (1998). « Аифан лейостахис » . Красный список исчезающих видов МСОП . 1998 : e.T38942A10158301. doi : 10.2305/IUCN.UK.1998.RLTS.T38942A10158301.en .
- ^ Монтуфар, Р.; Питман, Н. (2003). « Аифан бородавчатый » . Красный список исчезающих видов МСОП . 2003 : e.T38069A10095781. doi : 10.2305/IUCN.UK.2003.RLTS.T38069A10095781.en .
- ^ Хихон-Велес, Н.; Куврёр, TLP (2024). « Аифан чирибогенсис » . Красный список исчезающих видов МСОП . 2024 : e.T38752A230955824 . Проверено 30 июня 2024 г.
- ^ Лопес-Гальего, К.; Моралес М., Пенсильвания (2023). « Айфан Дукей » . Красный список исчезающих видов МСОП . 2023 : e.T38941A67530144. doi : 10.2305/IUCN.UK.2023-1.RLTS.T38941A67530144.es .
- ^ Бернал, Р. (1998). « Аифан Линдениана » . Красный список исчезающих видов МСОП . 1998 : e.T38943A10158352. doi : 10.2305/IUCN.UK.1998.RLTS.T38943A10158352.en .
- ^ Берналь, Родриго; Глория Галеано (2006). «Угроза колумбийским пальмам (Arecaceae): изменения за 18 лет». Ботанический журнал Линнеевского общества . 151 (1): 151–163. дои : 10.1111/j.1095-8339.2006.00530.x .
- ^ Свеннинг, Йенс-Кристиан (1998). «Влияние землепользования на местное распространение видов пальм во фрагменте тропического леса Анд на северо-западе Эквадора». Биоразнообразие и сохранение . 7 (12): 1529–1537. Бибкод : 1998BiCon...7.1529S . дои : 10.1023/A:1008831600795 . S2CID 34175807 .
- ^ Кан, Фрэнсис; Фарана Мусса (1994). «Разнообразие и статус сохранения перуанских пальм» (PDF) . Биоразнообразие и сохранение . 3 (3): 227–241. Бибкод : 1994BiCon...3..227K . дои : 10.1007/BF00055940 . S2CID 32827272 .
- ^ Jump up to: а б Борхсениус и Бернал (1996), стр. 30–32
- ^ Кулзак-Уилсон, Листра (2005). План сохранения вида попугая Сент-Винсента Amazona guildingii . Пуэрто-де-ла-Крус, Тенерифе: Фонд Лоро-парка.
- ^ Jump up to: а б Инман, Фейт М.; Томас Р. Вентворт; Марта Грум; Кэвелл Брауни ; Расс Ли (2007). «Использование искусственных просветов в пологе для восстановления среды обитания пуэрто-риканского попугая ( Amazona vittata ) на тропических лесных плантациях». Лесная экология и управление . 243 (2–3): 169–177. Бибкод : 2007ForEM.243..169I . дои : 10.1016/j.foreco.2007.02.003 .
- ^ Свеннинг, Ж.-К. (2001). «Неоднородность окружающей среды, ограничение пополнения и мезомасштабное распространение пальм в тропических горных дождевых лесах (Макипукуна, Эквадор)». Журнал тропической экологии . 17 (1): 97–113. дои : 10.1017/S0266467401001067 . S2CID 83543567 .
- ^ Корреа-Гомес, Диего Ф.; Орландо Варгас-Риос (2009). «Восстановление пальм в естественных лесах и плантациях заповедника фауны и флоры Отун-Кимбайя (Рисаральда, Колумбия)» . Кальдасия (на испанском языке). 31 (2): 195–212. Архивировано из оригинала 27 сентября 2011 г.
- ^ Моркоте-Риос, Гаспар; Родриго Берналь (2001). «Остатки пальм (Palmae) на археологических памятниках Нового Света: обзор». Ботанический обзор . 67 (3). Издательство Нью-Йоркского ботанического сада: 309–350. Бибкод : 2001BotRv..67..309M . дои : 10.1007/BF02858098 . JSTOR 4354394 . S2CID 46582757 .
- ^ Борхсениус и Бернал (1996), с. 47
- ^ Хендерсон, Эндрю ; Глория Галеано ; Родриго Берналь (1995). Полевой путеводитель по пальмам Америки . Принстон, Нью-Джерси: Издательство Принстонского университета. стр. 100-1 171–174. ISBN 0-691-08537-4 .
- ^ Риффл, Роберт Ли (2008). Путеводитель по прессам для древесины по пальмам . Лесной пресс. стр. 30–32. ISBN 978-0-88192-776-4 .
- ^ Борхсениус и Бернал (1996), с. 52
- ^ Борхсениус и Бернал (1996), с. 54
- ^ Борхсениус и Бернал (1996), с. 69
- ^ Борхсениус и Бернал (1996), с. 72
- ^ Борхсениус и Бернал (1996), с. 71
- ^ Банвелл, Мартин Г.; Анна Безос; Сатиш Чанд; Герд Даннхардт; Вернер Кифер; Ульрике Нове; Кристофер Р. Пэриш; Дж. Пол Сэвидж; Хольгер Ульбрих (2003). «Конвергентный синтез и предварительные биологические оценки стильбенолигнана (±)-айфанола и различных его соединений». Органическая и биомолекулярная химия . 1 (14): 2427–2429. дои : 10.1039/b305106d . ПМИД 12956057 .
- ^ Гудселл, Дэвид С. (1 мая 2001 г.). «Циклооксигеназа» . Банк данных белков RCSB. Архивировано из оригинала 23 ноября 2009 г. Проверено 17 января 2010 г.
Ссылки
[ редактировать ]- Асеведо-Родригес, Педро; Марк Т. Стронг (2005). Однодольные и голосеменные растения Пуэрто-Рико и Виргинских островов . Материалы из Национального гербария США. Том. 52. Вашингтон, округ Колумбия: Смитсоновский институт. стр. 1–415.
- Борхсениус, Финн; Родриго Берналь (декабрь 1996 г.). « Аифан (Пальмы)». Флора Неотропика . 70 . Пресса Нью-Йоркского ботанического сада от имени Организации флоры неотропики: 1–94. JSTOR 4393869 .
