Медоносная пчела восточноафриканской низменности
| Медоносная пчела восточноафриканской низменности | |
|---|---|
 | |
| Рабочая пчела (самка) питьевая вода | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Членистоногие |
| Сорт: | Насекомое |
| Заказ: | Перепончатокрылые |
| Семья: | Апиды |
| Род: | Апис |
| Разновидность: | |
| Подвиды: | Являюсь. щиток |
| Трехчленное имя | |
| Apis mellifera scutellata Лепелетье , 1836 г. | |
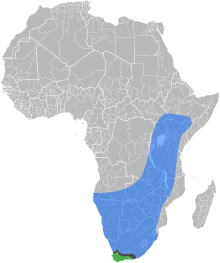 | |
| Естественные ареалы Восточноафриканская равнинная медоносная пчела , Мыс медоносной пчелы и зона контакта, где два подвида перекрываются и гибридизируются | |
Восточноафриканская равнинная медоносная пчела ( Apis mellifera scutellata ) — подвид западной медоносной пчелы . Он произрастает в центральной, южной и восточной Африке, хотя на крайнем юге его заменяет капская медоносная пчела ( Apis mellifera capensis ). [1] Было установлено, что этот подвид представляет собой часть предков африканизированных пчел (также известных как «пчелы-убийцы»), распространившихся по Северной и Южной Америке. [2]
Интродукция капской медоносной пчелы в северную часть Южной Африки представляет угрозу для низинных медоносных пчел Восточной Африки. Если работница из семьи медоносных пчел на мысе входит в гнездо медоносных пчел в низинах Восточной Африки, на нее не нападают, отчасти из-за ее сходства с маткой медоносных пчел в низинах Восточной Африки. Поскольку она способна к партеногенетическому размножению, она может начать откладывать яйца, из которых вылупляются «клоны» ее самой, которые также будут откладывать яйца, вызывая паразитическую A. m. capensis увеличится в количестве. Гибель колонии-хозяина происходит в результате сокращения численности A. m. scutellata , выполняющие обязанности по добыче пищи ( рабочие A. m. capensis значительно недостаточно представлены в составе кормодобывающих сил), смерть королевы и, перед смертью королевы, конкуренция за откладку яиц между A. m. рабочие capensis и королева. Когда колония погибнет, самки capensis начнут искать новую колонию-хозяина. [3]
Характер
[ редактировать ]Один укус низинной восточноафриканской пчелы не более ядовит , чем одиночный укус европейской пчелы, хотя низинные восточноафриканские медоносные пчелы реагируют быстрее, если их потревожить, чем европейские медоносные пчелы. В ответ на угрозу они посылают в три-четыре раза больше рабочих. Они также будут преследовать злоумышленника на большем расстоянии от улья. [ нужна ссылка ] Хотя люди погибли в результате 100–300 укусов пчел, подсчитано, что средняя смертельная доза для взрослого человека составляет 500–1100 укусов пчел. [ нужна ссылка ] Что касается промышленного производства меда, то в естественной среде обитания и неотропиках африканская пчела производит гораздо больше меда, чем ее европейские коллеги. [4] [5] Неясно, связано ли это с превосходной способностью собирать нектар, отсутствием адаптации европейских медоносных пчел к тропической среде. [4] или оба. Создание большего количества стаев и побег (покидание гнезда) также являются примерами адаптивных черт к тропической среде. Во времена длительного голода они мигрировали в район с лучшим источником пищи. [4] стратегия, применяемая также Apis dorsata , вместо того, чтобы ждать лучшего сезона (европейские и восточные пчелы). Отсутствие значительных энергетических потребностей для терморегуляции выводкового гнезда в тропиках соответствует очень быстрому созданию даже самых маленьких африканских колоний, стратегия роения чем выше по численности, тем меньше по размеру.
Появление
[ редактировать ]Внешность восточноафриканской равнинной медоносной пчелы очень похожа на европейскую пчелу. Однако медоносная пчела, обитающая в низинах Восточной Африки, немного мельче. Верхняя часть тела покрыта пухом, а брюшко покрыто черными полосами. [6]
среда обитания
[ редактировать ]Родной ареал Apis mellifera scutellata включает южные и восточные регионы Африки. Этот вид был впервые импортирован через Атлантический океан в Бразилию, а затем распространился в Центральную Америку, Южную Америку и южные районы Соединенных Штатов. Африканизированные медоносные пчелы процветают в тропических регионах и плохо приспособлены к холодным регионам с сильными дождями. [2]
Экономика кормодобывания и привычки пчел
[ редактировать ]Содержание нектара и сбор урожая
[ редактировать ]
Перед медоносными пчелами стоит задача сбалансировать потребление энергии и ее пополнение в поисках нектара. Высокие температуры в грудной клетке, необходимые для полета за кормом, создают терморегуляторный дисбаланс, который медоносные пчелы пытаются облегчить, ориентируясь на определенную вязкость и температуру ресурсов нектара. [7] было показано, На примере Apis mellifera scutellata что при более низких температурах окружающей среды, когда потеря энергии более выражена, медоносные пчелы стремятся к более теплому, менее концентрированному и менее вязкому нектару, что является энергетически выгодным поведением. [7]
Нектар с высокой концентрацией сахара более вязкий и, следовательно, снижает скорость потребления и размер нагрузки медоносных пчел. [7] При более низких температурах окружающей среды сбор небольших концентрированных количеств нектара не позволяет медоносным пчелам поддерживать обмен веществ, необходимый для полета за кормом. Сбор более теплого и менее вязкого нектара выгоден из-за энергии, получаемой за счет тепла. Медоносные пчелы способны стабилизировать температуру своего тела и восполнять энергию, потерянную при полете. [7] В А. М. scutellata , было обнаружено, что масса зоба в основном содержится в брюшке, хотя остается неясным, уравновешивает ли это вышеупомянутую потерю энергии из грудной клетки во время полета. [7]
Похоже, что стоимость сбора менее вязкого нектара заключается в том, что он также менее концентрирован в сахаре и будет представлять собой энергетическую потерю для медоносных пчел. Однако это не так; скорость сбора нектара меньшей вязкости увеличивает количество собираемого в данный момент времени. [8] Относительное преимущество настолько велико, что медоносной пчеле энергетически выгоднее собирать теплый нектар даже при низкой концентрации сахара (10%). [9] Медоносные пчелы получают энергетическое вознаграждение за сбор нектара, температура которого выше температуры окружающей среды, поскольку они компенсируют потери энергии во время кормления и легче получают больше нектара.
Способность шмелей различать теплоту цветов по цвету и нацеливаться на более теплые цветы является одним из известных прецедентов выбора температуры нектара у медоносных пчел. [10]
Значение собирательства
[ редактировать ]Было отмечено, что A. mellifera scutellata имеет более высокие темпы роста, размножения и роения колоний, чем европейские медоносные пчелы ( A. mellifera ligustica и A. mellifera mellifera ), что позволило им стать инвазивным видом. [11] Исследование Фьюэлла и Бертрама было проведено, чтобы понять источник этих различий. Считалось, что различия в стратегии приспособленности объясняются тем фактом, что африканские рабочие пчелы отдают предпочтение пыльце, а не нектару, который является прямым пищевым ресурсом для появляющегося выводка. [12] Считалось, что еще одним важным фактором являются различия между видами в возрастном полиэтизме или распределении различных задач по мере старения медоносной пчелы. [12] Молодые рабочие пчелы сосредоточены на помощи в улье, такой как уход за расплодом, и считалось, что относительно молодые популяции африканских пчел являются одним из объяснений акцента на воспроизводстве и расширении колоний этого вида. Исследование также интересовалось ролью, которую различные социальные среды колоний и различные генетические вариации могут играть в различиях в приспособленности между двумя подвидами. [12]
Поведенческие различия
[ редактировать ]
Основное различие, обнаруженное между африканскими и европейскими медоносными пчелами, заключалось в нескольких поведенческих чертах рабочих пчел, которые были связаны с пищевыми предпочтениями рабочих. [12] Было обнаружено, что рабочие Apis mellifera scutellata сосредоточились на поведении при переработке пыльцы, в то время как европейские рабочие сосредоточились на поведении при переработке нектара. Африканские пчелы также чаще хранили пыльцу, в то время как европейские пчелы хранили мед. Исследование показало, что пищевые предпочтения рабочих определяют, поддерживает ли колония определенный уровень воспроизводства. [12] Например, наличие меньшего количества или относительно более старых рабочих, предпочитающих нектар, означает, что у колонии не будет ресурсов для быстрого и эффективного выкармливания новых выводков. Предпочтения рабочих в еде связаны с генотипическими вариациями [13] в локусах конкретных количественных признаков. [14]
Африканские пчелы — «ранние собиратели»; Являюсь. scutellata начинают добывать пыльцу значительно раньше, чем их европейские собратья A. m. ligustica , и считается, что это связано с тем фактом, что африканские колонии имеют более молодое и асимметричное возрастное распределение по сравнению с ними. [15] Однако это не является прямой причиной различий в стратегиях существования этих двух подвидов. [12]
Компромиссы двух разных стратегий
[ редактировать ]Со временем распределение генотипических признаков пищевых предпочтений рабочих, должно быть, сгруппировалось вокруг тех, которые определяют склонность к использованию ресурсов, которые улучшают приспособленность подвида. Баланс эволюционных издержек и выгод сформировал распределение этих генотипических признаков. Популяция пчел должна найти баланс в распределении ресурсов в сторону роста нынешних членов семьи и воспроизводства. Если на поддержание взрослой семьи тратится слишком много энергии, пчелы потеряют возможность размножаться за счет размножения, но у них будут более старые рабочие, специализирующиеся на ресурсах нектара для получения энергии (меда). Если на размножение тратится слишком много энергии, такая колония будет менее приспособлена к тому, чтобы пережить резкие сезонные изменения, потому что у нее есть более молодые рабочие, которые специализируются на пыльце для кормления выводка, а не на хранении энергии. [12]
Эволюция стратегий истории жизни
[ редактировать ]Эти две стратегии были приняты европейскими и африканскими пчелами соответственно. Европейские пчелы должны пережить зиму — ежегодное событие с предсказуемыми последствиями смертности. Попытка удовлетворить энергетические потребности колонии и размножение может снизить их общую выживаемость зимой, и для них с эволюционной точки зрения более выгодно хранить нектар и мед. [12] [16] Африканские пчелы более уязвимы к менее предсказуемым периодам дефицита или нападения, и поэтому им выгодно производить как можно больше молодняка, увеличивая вероятность того, что некоторые или даже многие из них выживут. [12] [17] Такие обстоятельства благоприятствовали бы рабочим пчелам, которые предпочитали собирать нектар в европейских колониях и пыльцу в африканских колониях, что объясняет, как у A. m. развивались различия в поведении рабочих и возрастном распределении. scutellata и A.m. лигустика . [12] Исследование Фьюэлла и Бертрама важно тем, что оно предлагает правдоподобный метод, с помощью которого характеристики приспособленности подвида могли развиться из небольшого количества поведенческих различий у рабочих пчел.
Паразитирование
[ редактировать ]Apis mellifera capensis (капская медоносная пчела) монополизировала социальный паразитизм хозяев Apis mellifera scutellata в южном регионе Южной Африки. В частности, рабочие A. mellifera capensis производят важнейшие феромоны, достигают репродуктивного статуса и уничтожают A. m. королева щитовиков . Социальный паразитизм у общественных насекомых может включать различные формы эксплуатации, нарушающие нормальное разделение труда в колонии. [18] Недавнее развитие технологии изучения генетического состава колоний показало, что вклад потомства в размножение рабочих паразитов заслуживает более пристального внимания. [19]
В 1990 году в 400 утра. capensis были перемещены в окрестности A.m. подвид щитовки . Десять лет спустя появилась единственная клональная… рабочая линия. [20] было обнаружено, что это разрушительно A. m. scutellata на севере Южной Африки. [21] Монополия этой единственной линии показывает, что они смогли подорвать регулирование королевой воспроизводства и механизмы распознавания рабочих особей. Дитеманн и др. смог доказать, что А.м. capensis рабочие паразиты были способны производить нижнечелюстные феромоны, имитирующие феромоны A. m. королев scutellata в их присутствии. [ нужна ссылка ] Возникающее в результате нарушение разделения труда приводит к дезертирству или гибели паразитированной колонии.
Метод и результаты
[ редактировать ]Хотя многие феромоны способствуют воспроизводству, феромоны, вырабатываемые в нижнечелюстной железе королев, тесно связаны с воспроизводством, и они производятся рабочими, которые размножаются. Феромоны не позволяют другим нападать на них, побуждают рабочих признать их королевой и дают им доступ к более качественной пище. Они также мешают другим работникам стать репродуктивными. [22] Являюсь. capensis рабочие паразиты создают женские клоны и узурпируют A. m. королева щитовиков . Рабочие паразиты и их увеличивающееся число клонов становятся единственными репродуктивными особями в колонии. Разрушение разделения труда приводит к сокращению ресурсов, что в конечном итоге вынуждает колонию покинуть страну или погибнуть. [23]
Эволюция производства феромонов
[ редактировать ]Единая линия паразитирования A. m. capensis , возможно, получил эволюционное преимущество, потому что, по сравнению с другими родственными видами, он не восприимчив к феромонному подавлению репродуктивных рабочих рабочих со стороны королевы-хозяина. Неинвазивные разновидности A. m. capensis производят меньше нижнечелюстного секрета, чем инвазивный штамм. Кроме того, они производят секреты, не столь похожие на секреты A. m. scutellata Queens как у инвазионного штамма. Единственная линия была выбрана из-за ее большей устойчивости и большей способности имитировать и подавлять феромональную регуляцию маток-хозяев. [20]
Феромональные различия
[ редактировать ]Выяснилось, что А.м. Королевы scutellata производят больше феромонов, чем A. m. capensis Queens, предполагая, что качество или содержание феромонов, а не количество, может объяснить, как A. m. рабочие capensis способны игнорировать сигналы королевы-хозяина. Феромональные различия между подвидами — это вопрос, требующий более глубокого исследования, чтобы понять, как такое паразитирование стало возможным. Поскольку нижнечелюстные феромоны были в центре внимания Dietemann et al. исследования вполне вероятно, что разные железы вносят вклад в выработку феромонов, связанных с репродуктивным статусом. [20]
Эволюционные преимущества и недостатки
[ редактировать ]Многогранный аспект общения социальных насекомых позволяет легко захватывать колонии социальных насекомых. [24] Биология и организация потенциальных видов-хозяев, особенно в случае близкородственных видов и подвидов, аналогичны биологии и организации потенциальных видов-паразитаторов, что облегчает их проникновение. С другой стороны, потенциальные паразиты сталкиваются с проблемой обнаружения королевой-хозяином, обычно единственной репродуктивной особью в колонии. Существование А.м. Рабочие паразиты capensis являются примером альтернативной эволюционной стратегии, которая позволяет им повысить свою «прямую приспособленность в иностранных колониях, а не инклюзивную приспособленность в своих натальных гнездах». [20] Рабочие обычно сосредотачивают свои усилия на выращивании и уходе за родственными им личинками, сохраняя таким образом распространение их генов и способствуя их инклюзивной приспособленности. По сравнению с этим паразитическая модель более выгодна, поскольку она позволяет рабочим напрямую воспроизводить потомство, которое более близко к ним родственно и в большем количестве, поэтому они являются компонентом прямой приспособленности. [25]
Инвазионная линия A. m. capensis преуспели либо из-за неспособности распознать хозяина A. m. Королева щитков подает правильный сигнал или сопротивление сигналу. В конечном счете, это интересный пример ранее существовавшей слабости к социальному паразитизму А.М. capensis в A. m. скутеллата . [20] Организмы развивают репродуктивные стратегии, которые обеспечивают выживание и распространение генов организмов. Успешные репродуктивные стратегии справляются с конкретными экономическими ограничениями, с которыми сталкивается организм. Паразитарные отношения между A. m. scutellata и A.m. capensis является примером того, как обычно успешная стратегия химического распознавания и поддержания репродуктивного разделения труда может быть подорвана конкурирующими эксплуататорскими стратегиями. [26]
Эволюция
[ редактировать ]Основная гипотеза агрессивного поведения низинных медоносных пчел Восточной Африки основана на идее о том, что эта раса пчел развивалась в засушливой среде, где пчелам было мало пищи. В этой ситуации отбор отдавал предпочтение более агрессивным семьям, которые защищали свой источник пищи и улей от хищников и грабили пчел из других семей. Такое поведение позволило более агрессивным колониям выжить, тогда как менее агрессивные колонии в конечном итоге были отобраны естественным отбором . [ нужна ссылка ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Руттнер, Ф. 1988: Биогеография и таксономия медоносных пчел. Шпрингер Верлаг, Берлин
- ^ Перейти обратно: а б Мастерсон, Дж. «Apis mellifera scutellata» . Смитсоновская морская станция в Форт-Пирсе . Проверено 8 декабря 2013 г.
- ^ [1] Мориц, RFA (2002) Капская медоносная пчела Apis mellifera capensis От рабочих-несушек к социальным паразитам Apidologie Special Issue 33 (2), 99–244
- ^ Перейти обратно: а б с Риндерер, Т.Э.; Хеллмих, Р.Л. (1981). «Африканская» медоносная пчела. Процессы африканизации (PDF) . ЦРК Пресс. ISBN 978-0-429-30874-1 .
- ^ Флетчер, Дэвид Дж. К. (2009). «Африканские (низированные) пчелы: их история и будущее». Американский пчелиный журнал . 149 (9): 863–870.
- ^ Матерсон, Дж. «Apis mellifera scutellata» . Смитсоновский морской вокзал . Проверено 1 ноября 2013 г.
- ^ Перейти обратно: а б с д и Николсон, Сьюзен; Лео де Веер; Анджела Колер; Кристиан В.В. Пирк (2013). «Медоносные пчелы предпочитают более теплый и менее вязкий нектар, независимо от концентрации сахара» . Труды Королевского общества B: Биологические науки . 280 (1767): 1–8. дои : 10.1098/rspb.2013.1597 . ПМЦ 3735266 . ПМИД 23902913 .
- ^ Шафир С., Афик О (2007). «Влияние температуры окружающей среды на нагрузку урожая медоносной пчелы Apis mellifera (Hymenoptera: Apidae)». Общая энтомология . 29 (2–4): 135–148. дои : 10.1127/entom.gen/29/2007/135 .
- ^ Хейнеман, Эй Джей (1983). «Оптимальная концентрация сахара в цветочных нектарах: зависимость от эффективности потребления сахара и затрат на корм». Экология . 60 (2): 198–213. Бибкод : 1983Oecol..60..198H . дои : 10.1007/bf00379522 . ПМИД 28310487 . S2CID 32118291 .
- ^ А.Г., Дайер; Уитни Х.М.; Арнольд С.Э.Дж.; Гловер Би Джей; Читтка Л (2006). «Пчелы ассоциируют тепло с цветом цветов» . Природа . 442 (7102): 525. дои : 10.1038/442525a . ПМИД 16885975 . S2CID 432440 .
- ^ Уинстон, МЛ; ОР Тейлор; Г. В. Отис (1983). «Некоторые различия между медоносными пчелами умеренной Европы, тропической Африки и Южной Америки». Пчелиный мир . 64 : 12–21. дои : 10.1080/0005772X.1983.11097902 .
- ^ Перейти обратно: а б с д и ж г час я дж Фьюэлл, Дженнифер Х.; Сьюзен М. Бертрам (2002). «Доказательства генетических различий в выполнении рабочих задач африканскими и европейскими медоносными пчелами». Поведенческая экология и социобиология . 52 (4): 318–325. дои : 10.1007/s00265-002-0501-3 . S2CID 22128779 .
- ^ Пейдж, RE; Робинсон Дж. Э. (1991). «Генетика разделения труда в пчелиных семьях». Достижения физиологии насекомых . 23 : 118–169. дои : 10.1016/s0065-2806(08)60093-4 . ISBN 978-0-12-024223-8 .
- ^ Хант, Дж; Страница Р; Фондрк М; Даллум С. (1995). «Основные локусы количественных признаков, влияющие на пищевое поведение медоносных пчел» . Генетика . 141 (4): 1537–1545. дои : 10.1093/генетика/141.4.1537 . ПМК 1206885 . ПМИД 8601492 .
- ^ Гирей, Т; Гузман-Новоа Э; Арон CW; Зелинский Б; Фарбах SE; Робинсон Дж. Э. (2000). «Генетические вариации временного полиэтизма рабочих и защиты колоний у медоносных пчел Apis mellifera » . Поведенческая экология . 11 : 44–55. дои : 10.1093/beheco/11.1.44 .
- ^ Сили, Т.Д. (1978). «Стратегия истории жизни медоносной пчелы Apis mellifera ». Экология . 32 (1): 109–118. Бибкод : 1978Oecol..32..109S . дои : 10.1007/bf00344695 . ПМИД 28308672 . S2CID 12784020 .
- ^ МакНелли, Л; Шнайдер С (1996). «Пространственное распространение и биология гнездования семей африканской медоносной пчелы» . Экологическая энтомология . 25 : 643–652. дои : 10.1093/ee/25.3.643 .
- ^ Шмид-Хемпель, Пол (1998). Паразиты у социальных насекомых . Издательство Принстонского университета. ISBN 0-691-05924-1 .
- ^ Хартель, С; Нойманн П; Раассен Ф.С.; Мориц RFA; Хепберн HR (2006). «Социальный паразитизм рабочих капских пчел в колониях их собственных подвидов ( Apis mellifera capensis Esch.)». Общество насекомых . 53 (2): 183–193. дои : 10.1007/s00040-005-0857-2 . S2CID 44927091 .
- ^ Перейти обратно: а б с д и Дитеманн, Винсент; Йохен Пфлюгфельдер; Стефан Хартель; Питер Нойманн; Робин М. Крю (6 октября 2006 г.). «Социальный паразитизм рабочих пчел ( Apis mellifera capensis Esch.): Доказательства феромонного сопротивления сигналам королевы-хозяина». Поведенческая экология и социобиология . 60 (6): 785–793. дои : 10.1007/s00265-006-0222-0 . JSTOR 25063876 . S2CID 1218222 .
- ^ Хартель, С; Нойманн П; Крайгер П; фон дер Хайде С; Мольцер Г.Дж.; Крю Р.М.; ван Прааг Дж.П.; Мориц RFA (2006). «Уровни заражения роями Apis mellifera scutellata социально-паразитическими рабочими медоносными пчелами мыса ( Apis mellifera capensis Esch.)» . Апидология . 37 : 462–470. дои : 10.1051/apido:2006012 .
- ^ Гувер, СЭР; Привет, ХА; Уинстон МЛ (2006). «Сезонные колебания развития яичников рабочих пчел и влияние питания личинок и взрослых особей». Журнал сравнительной физиологии Б. 176 (1): 55–63. дои : 10.1007/s00360-005-0032-0 . ПМИД 16228242 . S2CID 8253649 .
- ^ Нойманн, П; Мориц RFA (2002). «Феномен капской медоносной пчелы: они наблюдают за эволюцией социального паразита в реальном времени». Поведенческая экология и социобиология . 52 (4): 271–281. дои : 10.1007/s00265-002-0518-7 . S2CID 12117374 .
- ^ Холлдоблер, Б; Уилсон Э.О. (1990). Муравьи . Спрингер.
- ^ Дэвис, Николас Б. (2012). Введение в поведенческую экологию . Западный Суссекс, Великобритания: Blackwell Publishing. стр. 307–33. ISBN 978-1-4051-1416-5 .
- ^ Докинз, Ричард (2006). Эгоистичный ген . Нью-Йорк: Издательство Оксфордского университета. стр. 66–88 . ISBN 978-0-19-929115-1 .