Кладограмма

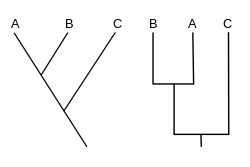
Кладограмма кладистике (от греческого кладо «ветвь» и характер грамма ») - это диаграмма, используемая в для демонстрации отношений между организмами. Кладограмма, однако, не является эволюционным деревом, потому что она не показывает, как предки связаны с потомками, и не показывает, насколько они изменились, так много различных эволюционных деревьев могут соответствовать одной и той же кладограмме. [ 1 ] [ 2 ] [ 3 ] [ 4 ] [ 5 ] Кладограмма использует линии, которые разветвляются в разных направлениях, заканчивающихся кладкой , группой организмов с последним общим предком . Есть много форм кладограмм, но все они имеют линии, которые отражаются от других линий. Линии можно проследить до того места, где они разветвляются. Эти точки разветвления представляют гипотетический предок (а не фактическое сущность), который можно вывести, чтобы показать признаки, разделяемые среди терминальных таксонов над ним. [ 4 ] [ 6 ] Этот гипотетический предок может затем дать подсказки о порядке эволюции различных особенностей, адаптации и других эволюционных повествований о предках. Хотя традиционно такие кладограммы генерировались в основном на основе морфологических признаков, ДНК и РНК данные секвенирования и вычислительная филогенетика в настоящее время часто используются в генерации кладограмм, либо самостоятельно, либо в сочетании с морфологией.
Генерируя кладограмму
[ редактировать ]В этом разделе нужны дополнительные цитаты для проверки . ( Апрель 2016 ) |
Молекулярные и морфологические данные
[ редактировать ]Характеристики, используемые для создания кладограммы, могут быть примерно классифицированы как морфологические (синапсид черепа, теплое кровообращение, нотокорр , одноклеточное и т. Д.), либо молекулярное (ДНК, РНК или другая генетическая информация). [ 7 ] До появления секвенирования ДНК кладистический анализ в основном использовал морфологические данные. Поведенческие данные (для животных) также могут использоваться. [ 8 ]
Поскольку секвенирование ДНК стало дешевле и проще, молекулярная систематика стала более популярным способом вывода филогенетических гипотез. [ 9 ] Использование критерия скупости является лишь одним из нескольких методов для вывода филогения из молекулярных данных. Такие подходы, как максимальная вероятность , которые включают в себя явные модели эволюции последовательностей, представляют собой незеннигенские способы оценки данных последовательности. Другим мощным методом реконструкции филогении является использование геномных ретротранспозонов , которые, как считается, менее подвержены проблеме реверсии , которая извлекает данные последовательности. Как правило, они также имеют низкую частоту гомоплазий, потому что когда -то считалось, что их интеграция в геном была совершенно случайной; Однако это, по крайней мере, иногда не так.

Плезиоморфии и синапоморфии
[ редактировать ]Исследователи должны решить, какие состояния характера являются «наследственными» ( плезиоморфий ), а какие получены ( синапоморфии ), потому что только состояния синапоморфного характера предоставляют доказательства группировки. [ 10 ] Это определение обычно выполняется по сравнению с состояниями персонажа одной или нескольких внешних групп . Государства, разделяемые между внешней группой, и некоторыми членами внутригруппа являются симплсуоморфиями; Состояния, которые присутствуют только в подмножестве внутри группы, являются синапоморфиями. Обратите внимание, что характер, уникальные для одного терминала (аутапоморфии), не дают доказательств группировки. Выбор внешней группы является решающим шагом в кладистическом анализе, потому что разные внешние группы могут производить деревья с глубоко разными топологиями.
Homplays
[ редактировать ]Гомоплазия отличной - это состояние персонажа, которое разделяется двумя или более таксонами из -за какой -то причины, от общего происхождения. [ 11 ] Двумя основными типами гомоплазии являются сходимость (эволюция «того же» характера, по крайней мере, в двух различных линиях) и реверсию (возвращение к состоянию наследственного характера). Персонажи, которые, очевидно, гомопластичны, такие как белый мех в разных линиях арктических млекопитающих, не должны быть включены в качестве характера в филогенетический анализ, поскольку они ничего не способствуют нашему пониманию отношений. Тем не менее, гомоплазия часто не очевидна из проверки самого персонажа (например, в последовательности ДНК), а затем обнаруживается по его несоответствию (непредубежденное распределение) на наиболее важной кладограмме. Обратите внимание, что гомопластические признаки могут содержать филогенетический сигнал . [ 12 ]
Хорошо известным примером гомоплазии, вызванной сходящейся эволюцией, будет характер, «Наличие крыльев». Хотя крылья птиц, летучих мышей и насекомых служат одной и той же функции, каждая из них развивалась независимо, что видно по их анатомии . Если бы птица, летучая мышь и крылатое насекомое оценивалось для персонажа, «присутствие крыльев», в набор данных гомоплазию была бы введена, и это может потенциально запутать анализ, возможно, что приведет к ложной гипотезе отношений. Конечно, единственная причина, по которой гомоплазию в первую очередь узнаваем, заключается в том, что есть другие признаки, которые подразумевают схему отношений, которые раскрывают его гомопластическое распределение.
Что не кладограмма
[ редактировать ]В этом разделе нужны дополнительные цитаты для проверки . ( Январь 2021 г. ) |
Кладограмма - это схематический результат анализа, который группирует таксоны только на основе синапоморфий. Есть много других филогенетических алгоритмов, которые обрабатывают данные несколько по -разному и приводят к филогенетическим деревьям, которые похожи на кладограммы, но не являются кладограммами. Например, фенотические алгоритмы, такие как Upgma и соседние, группировки по общему сходству, и рассматривать как синапоморфии, так и симплсуоморфии как доказательства группировки, полученные диаграммы-это фенограммы, а не кладограммы, аналогично, результаты модели (максимум (максимум. Вероятность или байесовские подходы), которые принимают во внимание как порядок ветвления, так и «длина ветвления», считают как синапоморфии, так и Аутапоморфии. В качестве доказательства или против группировки диаграммы, полученные в результате такого рода анализа, также не являются кладограммами. [ 13 ]
Выбор кладограммы
[ редактировать ]Есть несколько алгоритмов , доступных для определения «лучшей» кладограммы. [ 14 ] Большинство алгоритмов используют метрику , чтобы измерить, насколько согласованная кладограмма кандидата с данными. Большинство алгоритмов кладограммы используют математические методы оптимизации и минимизации.
В целом, алгоритмы генерации кладограмм должны быть реализованы в качестве компьютерных программ, хотя некоторые алгоритмы могут выполняться вручную, когда наборы данных являются скромными (например, лишь несколько видов и пара характеристик).
Некоторые алгоритмы полезны только тогда, когда характерные данные являются молекулярными (ДНК, РНК); Другие алгоритмы полезны только тогда, когда характерные данные являются морфологическими. Другие алгоритмы могут использоваться, когда характерные данные включают как молекулярные, так и морфологические данные.
Алгоритмы для кладограмм или других типов филогенетических деревьев включают в себя наименьшие квадраты , соседние , скупость , максимальное вероятность и байесовский вывод .
Биологи иногда используют термин экономии для определенного вида алгоритма генерации кладограмм, а иногда в качестве зонтичного термина для всех филогенетических алгоритмов. [ 15 ]
Алгоритмы, которые выполняют задачи оптимизации (такие как строительные кладограммы), могут быть чувствительны к порядку, в котором представлены входные данные (список видов и их характеристики). Ввод данных в различных порядках может привести к тому, что один и тот же алгоритм создает различные «лучшие» кладограммы. В этих ситуациях пользователь должен ввести данные в различных заказах и сравнить результаты.
Использование различных алгоритмов на одном наборе данных может иногда давать различные «лучшие» кладограммы, потому что каждый алгоритм может иметь уникальное определение того, что является «лучшим».
Из -за астрономического количества возможных кладограмм алгоритмы не могут гарантировать, что решение является лучшим решением. Неоптимальная кладограмма будет выбран, если программа останется на локальном минимуме, а не на желаемый глобальный минимум. [ 16 ] Чтобы помочь решить эту проблему, многие алгоритмы кладограммы используют моделируемый подход отжига, чтобы увеличить вероятность того, что выбранная кладограмма является оптимальной. [ 17 ]
Базальное положение - это направление основания (или корня) корневого филогенетического дерева или кладограммы. Базальная клада - самая ранняя клада (данного таксономического ранга [A]), чтобы разветвляться в большей кладе.
Статистика
[ редактировать ]Испытание на разницу в длины некогена
[ редактировать ]Испытание на разницу в длины неконгерации (ILD) представляет собой измерение того, как комбинация различных наборов данных (например, морфологические и молекулярные, пластидные и ядерные гены) способствует более длинному дереву. Он измеряется путем первого расчета общей длины дерева каждого раздела и суммирования их. Затем повторяются, делая случайно собранные разделы, состоящие из исходных разделов. Длина суммирована. Значение P 0,01 получается для 100 повторностей, если 99 повторений имеют более длинную комбинированную длину деревьев.
Измерение гомоплазии
[ редактировать ]Некоторые меры пытаются измерить количество гомоплазии в наборе данных со ссылкой на дерево, [ 18 ] хотя не обязательно ясно, какое свойство стремится к количественной оценке количественной оценки [ 19 ]
Индекс согласованности
[ редактировать ]Индекс согласованности (CI) измеряет согласованность дерева на набор данных - мера минимального количества гомоплазии, подразумеваемой деревом. [ 20 ] Он рассчитывается путем подсчета минимального количества изменений в наборе данных и деления его на фактическое количество изменений, необходимых для кладограммы. [ 20 ] Индекс согласованности также можно рассчитать для отдельного персонажа I , обозначенный C i .
Помимо отражения количества гомоплазии, метрика также отражает количество таксонов в наборе данных, [ 21 ] (в меньшей степени) количество символов в наборе данных, [ 22 ] степень, в которой каждый персонаж несет филогенетическую информацию, [ 23 ] и мода, в которой аддитивные персонажи закодированы, что делает его непригодным для цели. [ 24 ]
C I занимает диапазон от 1 до 1 /[N.Taxa / 2] бинарными символами с равномерным распределением состояния; Его минимальное значение больше, когда состояния не распространяются. [ 23 ] [ 18 ] В целом, для бинарного или невоичного персонажа с , c я занимает диапазон от 1 до . [ 23 ]


Индекс удержания
[ редактировать ]Индекс удержания (RI) был предложен в качестве улучшения CI «для определенных применений» [ 25 ] Этот показатель также подразумевает измерение количества гомоплазии, но также измеряет, насколько хорошо синапоморфии объясняют дерево. Он рассчитывается, принимая (максимальное количество изменений на дереве за вычетом количества изменений на дереве) и деление на (максимальное количество изменений на дереве минус минимальное количество изменений в наборе данных).
Измененный индекс согласованности (RC) получается путем умножения CI на RI; По сути, это растягивает диапазон CI, так что его минимальное теоретически достижимое значение пересекается до 0, с максимальным оставшимся на уровне 1. [ 18 ] [ 25 ] Индекс гомоплазии (HI) просто 1 - CI.
Отношение гомоплазии
[ редактировать ]Это измеряет количество гомоплазии, наблюдаемое на дереве относительно максимального количества гомоплазии, которое теоретически может присутствовать - 1 - (наблюдаемый избыток гомоплазии) / (максимальный избыток гомоплазии). [ 22 ] Значение 1 указывает на гомоплазию; 0 представляет столько гомоплазии, сколько было бы в полностью случайном наборе данных, а негативные значения указывают на большую гомоплазию (и, как правило, встречаются только в надуманных примерах). [ 22 ] Ее представлены как лучшая мера гомоплазии в настоящее время. [ 18 ] [ 26 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Mayr, Ernst (2009). "Кладистический анализ или кладистическая классификация?" Полем Журнал зоологической систематики и эволюционных исследований . 12 : 94–128. doi : 10.1111/j.1439-0469.1974.tb00160.x .
- ^ Фут, Майк (весна 1996). «О вероятности предков в ископаемом записи». Палеобиология . 22 (2): 141–51. doi : 10.1017/s0094837300016146 . JSTOR 2401114 . S2CID 89032582 .
- ^ Дейрат, Бенуа (лето 2005). «Отношения предков пожилых людей и реконструкция дерева жизни». Палеобиология . 31 (3): 347–53. doi : 10.1666/0094-8373 (2005) 031 [0347: aratro] 2.0.co; 2 . JSTOR 4096939 . S2CID 54988538 .
- ^ Jump up to: а беременный Посада, Дэвид; Крэндалл, Кит А. (2001). «Внутривидочные генные генеалогии: деревья, привитые в сетях». Тенденции в экологии и эволюции . 16 (1): 37–45. doi : 10.1016/s0169-5347 (00) 02026-7 . PMID 11146143 .
- ^ Podani, János (2013). «Мышление дерева, время и топология: комментарии о интерпретации диаграмм деревьев в эволюционной/филогенетической систематике» (PDF) . Кладистика . 29 (3): 315–327. doi : 10.1111/j.1096-0031.2012.00423.x . PMID 34818822 . S2CID 53357985 . Архивировано (PDF) из оригинала 2017-09-21.
- ^ Шух, Рэндалл Т. (2000). Биологическая систематика: принципы и приложения . Издательство Корнелльского университета. ISBN 978-0-8014-3675-8 . [ страница необходима ]
- ^ Desalle, Rob (2002). Методы в молекулярной систематике и эволюции . Биркхаузер. ISBN 978-3-7643-6257-7 . [ страница необходима ]
- ^ Венцель, Джон В. (1992). «Поведенческая гомология и филогения». Анну. Rev. Ecol. Система 23 : 361–381. doi : 10.1146/annurev.es.23.110192.002045 .
- ^ Хиллис, Дэвид (1996). Молекулярная систематика . Синаур. ISBN 978-0-87893-282-5 . [ страница необходима ]
- ^ Хенниг, Уилли (1966). Филогенетическая систематика . Университет Иллинойса Пресс.
- ^ Западный Эберхард, Мэри Джейн (2003). Пластичность и эволюция развития . Оксфордский Univ. Нажимать. С. 353 –376. ISBN 978-0-19-512235-0 .
- ^ Калерсджо, Мари; Альберт, Виктор А.; Фаррис, Джеймс С. (1999). «Гомоплазия увеличивает филогенетическую структуру». Кладистика . 15 : 91–93. doi : 10.1111/j.1096-0031.1999.tb00400.x . S2CID 85905559 .
- ^ Brower, Andrew VZ (2016). "Что такое кладограмма, а что нет?" Полем Кладистика . 32 (5): 573–576. doi : 10.1111/cla.12144 . PMID 34740305 . S2CID 85725091 .
- ^ Китчинг, Ян (1998). Кладистика: теория и практика анализа скупости . Издательство Оксфордского университета. ISBN 978-0-19-850138-1 . [ страница необходима ]
- ^ Стюарт, Каро-Бет (1993). «Сила и подводные камни скупости». Природа . 361 (6413): 603–7. Bibcode : 1993natur.361..603s . doi : 10.1038/361603a0 . PMID 8437621 . S2CID 4350103 .
- ^ Фоли, Питер (1993). Кладистика: практический курс по систематике . Оксфордский Univ. Нажимать. п. 66 ISBN 978-0-19-857766-9 .
- ^ Никсон, Кевин С. (1999). «Резумия, новый метод для анализа быстрого скупости». Кладистика . 15 (4): 407–414. doi : 10.1111/j.1096-0031.1999.tb00277.x . PMID 34902938 . S2CID 85720264 .
- ^ Jump up to: а беременный в дюймовый пересматривается в Арчи, Джеймс В. (1996). «Меры гомоплазии». В Сандерсоне, Майкл Дж.; Хаффорд, Ларри (ред.). Гомоплазия . С. 153 –188. doi : 10.1016/b978-012618030-5/50008-3 . ISBN 9780126180305 .
- ^ Чанг, Джозеф Т.; Ким, Джунхьонг (1996). «Измерение гомоплазии: стохастический взгляд». Гомоплазия . С. 189–203. doi : 10.1016/b978-012618030-5/50009-5 . ISBN 9780126180305 .
- ^ Jump up to: а беременный Kluge, Ag; Фаррис, JS (1969). «Количественная филетитика и эволюция ануранов». Систематическая зоология . 18 (1): 1–32. doi : 10.2307/2412407 . JSTOR 2412407 .
- ^ Арчи, JW; Felsenstein, J. (1993). «Количество эволюционных шагов на деревьях случайной и минимальной длины для случайных эволюционных данных». Теоретическая биология населения . 43 : 52–79. doi : 10.1006/tpbi.1993.1003 .
- ^ Jump up to: а беременный в Арчи, JW (1989). «Избыточные соотношения гомоплазий: новые индексы для измерения уровней гомоплазии в филогенетической систематике и критика индекса согласованности». Систематическая зоология . 38 (3): 253–269. doi : 10.2307/2992286 . JSTOR 2992286 .
- ^ Jump up to: а беременный в Хоял Катхилл, Дженнифер Ф.; Брэдди, Саймон Дж.; Donoghue, Philip CJ (2010). «Формула для максимально возможных шагов в многосторонних символах: изолирующее влияние параметра матрицы на меры эволюционной конвергенции» . Кладистика . 26 (1): 98–102. doi : 10.1111/j.1096-0031.2009.00270.x . PMID 34875753 . S2CID 53320612 .
- ^ Сандерсон, MJ; Donoghue, MJ (1989). «Модели вариаций в уровнях гомоплазии». Эволюция 43 (8): 1781–1795. doi : 10.2307/2409392 . JSTOR 2409392 . PMID 28564338 .
- ^ Jump up to: а беременный Фаррис, JS (1989). «Индекс удержания и измененный индекс согласованности». Кладистика . 5 (4): 417–419. doi : 10.1111/j.1096-0031.1989.tb00573.x . PMID 34933481 . S2CID 84287895 .
- ^ Хоял Катхилл, Дженнифер (2015). «Размер пространства состояния характера влияет на возникновение и обнаружение гомоплазии: моделирование вероятности несовместимости для неупорядоченных филогенетических признаков». Журнал теоретической биологии . 366 : 24–32. Bibcode : 2015jthbi.366 ... 24 ч . doi : 10.1016/j.jtbi.2014.10.033 . PMID 25451518 .
Внешние ссылки
[ редактировать ] СМИ, связанные с кладограммами в Wikimedia Commons
СМИ, связанные с кладограммами в Wikimedia Commons
| Соответствующие поля | ||
|---|---|---|
| Основные понятия | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Групповые типы | ||
| Номенклатура | ||