Гипотеза двух потоков
Гипотеза двух потоков представляет собой модель нейронной обработки зрения и слуха . [1] Гипотеза, первоначально описанная в статье Дэвида Милнера и Мелвина А. Гудейла в 1992 году, утверждает, что люди обладают двумя различными зрительными системами. [2] Недавно появились доказательства существования двух различных слуховых систем . Когда визуальная информация выходит из затылочной доли , а звук — из фонологической сети, она следует двумя основными путями, или «потоками». Вентральный поток (также известный как «путь что») ведет к височной доле, которая отвечает за идентификацию и распознавание объектов, а также зрительную идентификацию . Дорсальный поток (или «путь пути») ведет к теменной доле, которая участвует в обработке пространственного положения объекта относительно зрителя и повторении речи.
История
[ редактировать ]
Несколько исследователей ранее предлагали аналогичные идеи. Сами авторы отдают должное работе Вейскранца и по слепому зрению предыдущим нейробиологическим исследованиям зрения. Шнайдер впервые предположил существование двух зрительных систем для локализации и идентификации в 1969 году. [3] Ингл описал две независимые зрительные системы лягушек в 1973 году. [4] Эттлингер проанализировал существующие нейропсихологические доказательства различия в 1990 году. [5] Более того, Тревартен еще в 1968 году предложил объяснение двух отдельных механизмов зрения у обезьян. [6]
В 1982 году Унгерлейдер и Мишкин в своих исследованиях поражений обезьян выделили дорсальный и вентральный потоки, обрабатывающие пространственные и зрительные характеристики соответственно, предложив оригинальное различие «где и что». [7] Хотя эта концепция была заменена концепцией Милнера и Гудейла, она остается влиятельной. [8]
Одним из чрезвычайно влиятельных источников информации, которая легла в основу модели, была экспериментальная работа по изучению существующих способностей пациента с диагнозом DF, страдающего зрением. Первый и наиболее влиятельный отчет был сделан Гудейлом и его коллегами в 1991 году. [9] и работы о ней все еще публикуются два десятилетия спустя. [10] Это стало предметом некоторой критики модели из-за того, что она воспринималась как чрезмерная зависимость от результатов одного случая.
Две зрительные системы
[ редактировать ]Гудейл и Милнер [2] собрали множество анатомических, нейропсихологических, электрофизиологических и поведенческих данных для своей модели. Согласно их данным, вентральный «перцептивный» поток вычисляет подробную карту мира на основе зрительной информации, которую затем можно использовать для когнитивных операций, а дорсальный «действительный» поток преобразует поступающую визуальную информацию в необходимую эгоцентрическую (головоцентрированную). ) система координат для квалифицированного планирования движений .Модель также утверждает, что визуальное восприятие кодирует пространственные свойства объектов, такие как размер и расположение, относительно других объектов в поле зрения; другими словами, он использует относительные показатели и системы отсчета, основанные на сценах. С другой стороны, планирование и координация визуальных действий используют абсолютные показатели, определяемые с помощью эгоцентрических систем отсчета, вычисляя фактические свойства объектов относительно наблюдателя. Таким образом, было показано, что хватательные движения, направленные на объекты, включенные в сцены с неоднозначным контрастом размеров, избегают эффектов этих иллюзий, поскольку в восприятии иллюзии участвуют разные системы отсчета и метрики, а не в выполнении акта хватания. [11]
Норман [12] предложил аналогичную двухпроцессную модель зрения и описал восемь основных отличий между двумя системами, согласующихся с другими двухсистемными моделями.
| Фактор | Вентральная система (какая) | Дорсальная система (как) |
|---|---|---|
| Функция | Распознавание/идентификация | Визуально управляемое поведение |
| Чувствительность | Высокие пространственные частоты – подробности | Высокие временные частоты – движение |
| Память | Долговременно хранимые представления | Только очень кратковременное хранение |
| Скорость | Относительно медленно | Относительно быстро |
| Сознание | Обычно высокий | Обычно низкий |
| Система отсчета | Аллоцентрический или объектно-центрированный | Эгоцентричный или ориентированный на зрителя |
| Визуальный ввод | Преимущественно фовеальный или парафовеальный | Через сетчатку |
| Монокулярное зрение | Как правило, достаточно небольшие эффекты | Часто большие эффекты, например параллакс движения. |
Спинной ручей
[ редактировать ]Спинной поток предлагается участвовать в руководстве действиями и распознавании местоположения объектов в пространстве. Дорсальный поток проходит от первичной зрительной коры к задней теменной коре . Первоначально его называли путем «где», поскольку считалось, что спинной поток обрабатывает информацию, касающуюся пространственных свойств объекта. [13] Однако более позднее исследование, проведенное на известном нейропсихологическом пациенте, пациенте Д.Ф., показало, что дорсальный поток отвечает за обработку зрительной информации, необходимой для построения представлений объектов, которыми человек хочет манипулировать. Эти открытия привели к тому, что название спинного потока было обновлено до пути «как». [14] [15] Дорсальный поток взаимосвязан с параллельным вентральным потоком («поток «что»), который течет вниз от V1 в височную долю .
Общие характеристики
[ редактировать ]Дорсальный поток участвует в пространственном осознании и руководстве действиями (например, дотягиванием). При этом он имеет две различные функциональные характеристики: он содержит подробную карту поля зрения, а также хорошо обнаруживает и анализирует движения.
Дорсальный поток начинается с чисто зрительных функций в затылочной доле, а затем постепенно переходит к пространственному осознанию и заканчивается в теменной доле.
Задняя теменная кора необходима для «восприятия и интерпретации пространственных отношений, точного изображения тела и обучения задачам, связанным с координацией тела в пространстве». [16]
Он содержит индивидуально функционирующие дольки. Латеральная внутритеменная борозда (LIP) содержит нейроны, которые вызывают повышенную активацию, когда внимание переключается на стимул или саккады животного в сторону зрительного стимула, а также вентральная внутритеменная борозда (VIP), где интегрируются визуальная и соматосенсорная информация.
Последствия повреждений или поражений
[ редактировать ]Повреждение задней теменной коры вызывает ряд пространственных расстройств, в том числе:
- Симультанагнозия : пациент может описывать только отдельные объекты без способности воспринимать их как компонент набора деталей или объектов в контексте (как в сценарии, например, лес за деревьями).
- Оптическая атаксия : пациент не может использовать зрительно-пространственную информацию для управления движениями рук.
- Гемипространственная игнорация : когда пациент не осознает противоположную половину пространства (то есть он не осознает вещей в левом поле зрения и сосредотачивается только на объектах в правом поле зрения; или кажется, что он не осознает вещей в одном поле зрения). взгляд, когда они воспринимают их в другом). Например, человек с этим расстройством может нарисовать часы, а затем отметить все двенадцать цифр на одной стороне циферблата и считать рисунок завершенным.
- Акинетопсия : неспособность воспринимать движение.
- Апраксия : неспособность совершать произвольные или произвольные движения при отсутствии мышечных нарушений.
Вентральный поток
[ редактировать ]Вентральный поток связан с распознаванием объектов и представлением формы. Также называемый потоком «что», он имеет прочные связи с медиальной височной долей (которая связана с долговременной памятью ), лимбической системой (которая контролирует эмоции) и дорсальным потоком (который занимается расположением объектов и их движением). ).
Вентральный поток получает основной входной сигнал из ( в отличие от магноцеллюлярного ) слоя латерального коленчатого ядра таламуса парвоцеллюлярного . Эти нейроны проецируются на подслои V1 4Cβ, 4A, 3B и 2/3a. [17] последовательно. Отсюда вентральный путь проходит через V2 и V4 к областям нижней височной доли : PIT (задняя нижневисочная), CIT (центральная нижневисочная) и AIT (передняя нижневисочная). Каждая визуальная область содержит полное представление визуального пространства. То есть он содержит нейроны , рецептивные поля которых вместе представляют все поле зрения. Зрительная информация поступает в вентральный поток через первичную зрительную кору и последовательно проходит через остальные области.
Двигаясь по потоку от V1 к AIT, рецептивные поля увеличивают свой размер, латентность и сложность настройки. Например, недавние исследования показали, что область V4 отвечает за восприятие цвета у человека, а область V8 (VO1) — за восприятие формы, а область VO2, расположенная между этими областями и парагиппокампальной корой, интегрирует информацию. о цвете и форме раздражителей в целостный образ. [18]
Все области вентрального потока находятся под влиянием экстраретинальных факторов в дополнение к природе стимула в их рецептивном поле. Эти факторы включают внимание , рабочую память и значимость стимула . Таким образом, вентральный поток не просто дает описание элементов визуального мира — он также играет решающую роль в оценке значения этих элементов.
Повреждение вентрального потока может привести к невозможности распознавать лица или интерпретировать выражение лица. [19]
Две слуховые системы
[ редактировать ]Вентральный поток
[ редактировать ]
Наряду с зрительно- вентральным путем, важным для обработки зрительной информации, существует также вентральный слуховой путь, выходящий из первичной слуховой коры. [20] В этом пути фонемы обрабатываются после слогов и звуков окружающей среды. [21] Затем информация присоединяется к зрительному вентральному потоку в средней височной извилине и височном полюсе. Здесь слуховые объекты преобразуются в аудиовизуальные концепции. [22]
Спинной ручей
[ редактировать ]Функция слухового дорсального пути заключается в сопоставлении слуховых сенсорных представлений с артикуляционно-моторными представлениями. Хикок и Поппель утверждают, что слуховой дорсальный путь необходим, потому что «научение говорить — это, по сути, задача двигательного обучения. Основными входными данными для этого являются сенсорные ощущения, в частности речь. Таким образом, должен существовать нейронный механизм, который одновременно кодирует и поддерживает примеры». звуков речи и может использовать эти сенсорные следы для настройки речевых жестов, чтобы звуки точно воспроизводились». [23]
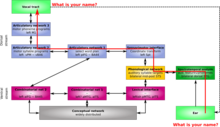
В отличие от слуховой обработки вентрального потока, информация поступает из первичной слуховой коры в заднюю верхнюю височную извилину и заднюю верхнюю височную борозду. Отсюда информация перемещается к началу дорсального пути, который находится на границе височной и теменной долей вблизи сильвиевой щели. Первый этап дорсального пути начинается в сенсомоторном интерфейсе, расположенном в левой сильвиевой теменно-височной области (Spt) (внутри сильвиевой щели на теменно-височной границе). СПТ важен для восприятия и воспроизведения звуков. Это очевидно, поскольку его способность приобретать новый словарный запас нарушается из-за поражений и слуховой обратной связи при воспроизведении речи, артикуляционного снижения при поздней глухоте и нефонологических остатков афазии Вернике; недостаточный самоконтроль. Это также важно для основных нейронных механизмов фонологической кратковременной памяти. Без Spt овладение языком ухудшается. Затем информация поступает в артикуляционную сеть, которая делится на две отдельные части. Артикуляционная сеть 1, обрабатывающая двигательные слоговые программы, расположена в левом заднем отделе. нижняя височная извилина и зона Бродмана 44 (pIFG-BA44). Артикуляционная сеть 2 предназначена для программ моторных фонем и расположена в левом M1-vBA6. [24]
Проводниковая афазия влияет на способность субъекта воспроизводить речь (обычно путем повторения), но не влияет на способность субъекта понимать устную речь. Это показывает, что проводящая афазия должна отражать поражение не слухового вентрального пути, а слухового дорсального пути. Бухсбаум и др. [25] обнаружили, что проводниковая афазия может быть результатом повреждения, особенно повреждений, Spt (сильвиева теменная височная мышца). Об этом свидетельствует участие Spt в приобретении нового словарного запаса, поскольку, хотя эксперименты показали, что большинство людей с афазией проводимости могут повторять высокочастотные простые слова, их способность повторять низкочастотные сложные слова нарушена. Spt отвечает за соединение двигательной и слуховой систем, делая слуховой код доступным для моторной коры. Похоже, что моторная кора воссоздает высокочастотные простые слова (например, чашка ), чтобы более быстро и эффективно получить к ним доступ, в то время как низкочастотные, сложные слова (например, сильвианская теменная височная ) требуют более активного онлайн-регулирования со стороны Spt. Это объясняет, почему страдающие афазией проводимости испытывают особые трудности с произнесением низкочастотных слов, что требует более практического процесса произнесения речи. «Функционально проводниковая афазия характеризуется как дефицит способности кодировать фонологическую информацию для производства», а именно из-за нарушения моторно-слухового интерфейса. [26] Проводниковая афазия более конкретно связана с повреждением дугообразного пучка , который жизненно важен как для речи, так и для понимания языка, поскольку дугообразный пучок образует связь между зонами Брока и Вернике. [26]
Критика
[ редактировать ]Нововведение Гудейла и Милнера заключалось в том, чтобы сместить акцент с акцента на входных различиях, таких как местоположение объекта и его свойства, на акцент на функциональной значимости видения для поведения, восприятия или действия. Однако современные взгляды, основанные на эмпирических исследованиях последних двух десятилетий, предлагают более сложную картину, чем простое разделение функций на два потока. [27] Недавние экспериментальные работы, например, поставили под сомнение эти выводы и предположили, что кажущаяся диссоциация между влиянием иллюзий на восприятие и действие обусловлена различиями во внимании, требованиями к выполнению задач и другими факторами. [28] [29] Однако есть и другие эмпирические данные, которые нельзя так легко отвергнуть, которые убедительно подтверждают идею о том, что на умелые действия, такие как хватание, графические иллюзии не влияют. [30] [31] [32] [33]
Более того, недавние нейропсихологические исследования поставили под сомнение обоснованность разделения двух потоков, которое послужило краеугольным камнем доказательств модели. Диссоциация между зрительной агнозией и зрительной атаксией была оспорена несколькими исследователями как не такая сильная, как первоначально предполагалось; Гессе и его коллеги продемонстрировали нарушения дорсального потока у пациента с ДФ; [34] Химмельбах и его коллеги переоценили способности Д.Ф. и применили более строгий статистический анализ, показавший, что диссоциация была не такой сильной, как казалось на первый взгляд. [10]
Обзор накопленных данных в пользу модели в 2009 году пришел к выводу, что, хотя дух модели был подтвержден, независимость двух потоков была переоценена. [35] Сами Гудейл и Милнер предложили аналогию с телепомощью, одной из наиболее эффективных схем, разработанных для дистанционного управления роботами, работающими во враждебных средах. С этой точки зрения дорсальный поток рассматривается как полуавтономная функция, которая действует под руководством исполнительных функций, которые сами получают информацию от обработки вентрального потока. [36]
Таким образом, новая перспектива в нейропсихологии и нейрофизиологии заключается в том, что, хотя двухсистемная структура была необходимым достижением для стимулирования изучения очень сложных и дифференцированных функций двух нервных путей; реальность, скорее всего, предполагает значительное взаимодействие между видением для действия и видением для восприятия. Роберт Макинтош и Томас Шенк резюмируют эту позицию следующим образом:
Нам следует рассматривать модель не как формальную гипотезу, а как набор эвристик, направляющих эксперимент и теорию. Различные информационные потребности для визуального распознавания и руководства действиями по-прежнему предлагают убедительное объяснение широкой относительной специализации дорсальных и вентральных потоков. Однако для прогресса в этой области нам, возможно, придется отказаться от идеи, что эти потоки работают в значительной степени независимо друг от друга, и обратиться к динамическим деталям того, как многие зрительные области мозга организуются от задачи к задаче в новые функциональные сети. [35] : 62 [ нужна проверка ]
См. также
[ редактировать ]- Обработка речи в мозгу
- Видение для восприятия и видение для действия
- Зрительная память
- Зрительная кора
- Визуальная система
- Магноцеллюлярная клетка
- Тектопульвинарный путь
- Слуховая система
- Распознавание объектов (когнитивная наука)
- Медиальная верхняя височная область
- Дугообразный пучок
- Затылочная доля
- Латеральное подушкообразное ядро
- Сердцевина подушки
- Фантомный контур
- Зонтик от солнца
- Конструктивная апраксия
- Латеральное коленчатое ядро
- Слепое зрение
- Кониоцеллюлярная клетка
Ссылки
[ редактировать ]- ^ Айсенк М.В., Кин М.Т. (2010). Когнитивная психология: Справочник для студентов . Хоув, Великобритания: Psychology Press.
- ^ Jump up to: а б Гудейл М.А., Милнер А.Д. (1992). «Отдельные визуальные пути восприятия и действия». Тенденции нейробиологии . 15 (1): 20–5. дои : 10.1016/0166-2236(92)90344-8 . ПМИД 1374953 . S2CID 793980 .
- ^ Шнайдер, GE. (февраль 1969 г.). «Две зрительные системы». Наука . 163 (3870): 895–902. Бибкод : 1969Sci...163..895S . дои : 10.1126/science.163.3870.895 . ПМИД 5763873 .
- ^ Ингл, Д. (сентябрь 1973 г.). «Две зрительные системы лягушки». Наука . 181 (4104): 1053–5. Бибкод : 1973Sci...181.1053I . дои : 10.1126/science.181.4104.1053 . ПМИД 4542178 . S2CID 42609212 .
- ^ Эттлингер Г. (1990). « «Объектное зрение» и «пространственное зрение»: нейропсихологические доказательства различия» . Кортекс . 26 (3): 319–41. дои : 10.1016/s0010-9452(13)80084-6 . ПМИД 2123426 . S2CID 4488069 .
- ^ Тревартен, CB. (1968). «Два механизма зрения у приматов». Психол Форш . 31 (4): 299–348. дои : 10.1007/bf00422717 . ПМИД 4973634 . S2CID 27943693 .
- ^ Мишкин М, Унгерлейдер Л.Г. (1982). «Вклад стриарных входов в зрительно-пространственные функции теменно-преокципитальной коры у обезьян». Поведение мозга Res . 6 (1): 57–77. дои : 10.1016/0166-4328(82)90081-X . ПМИД 7126325 . S2CID 33359587 .
- ^ Шенк, Томас; Макинтош, Роберт Д. (2010). «Есть ли у нас независимые визуальные потоки для восприятия и действия?». Когнитивная нейронаука . 1 (1): 52–62. дои : 10.1080/17588920903388950 . ISSN 1758-8928 . ПМИД 24168245 . S2CID 3560697 .
- ^ Гудейл, Массачусетс; Милнер, AD; Якобсон, Л.С.; Кэри, ДП. (январь 1991 г.). «Нейрологическая диссоциация между восприятием объектов и их схватыванием». Природа . 349 (6305): 154–6. Бибкод : 1991Natur.349..154G . дои : 10.1038/349154a0 . ПМИД 1986306 . S2CID 4238254 .
- ^ Jump up to: а б Химмельбах, М.; Беме, Р.; Карнат, ХО. (январь 2012 г.). «20 лет спустя: второй взгляд на двигательное поведение DF». Нейропсихология . 50 (1): 139–44. doi : 10.1016/j.neuropsychologia.2011.11.011 . ПМИД 22154499 . S2CID 7513099 .
- ^ Аглиоти С., ДеСуза Дж. Ф., Гудейл М. А. (1995). «Иллюзия контраста размеров обманывает глаз, но не руку» . Курс. Биол . 5 (6): 679–85. дои : 10.1016/S0960-9822(95)00133-3 . ПМИД 7552179 . S2CID 206111613 .
- ^ Норман Дж. (2002). «Две зрительные системы и две теории восприятия: попытка примирить конструктивистский и экологический подходы» . Наука о поведении мозга . 25 (1): 73–144. дои : 10.1017/s0140525x0200002x . ПМИД 12625088 .
- ^ Мишкин, Мортимер; Унгерлейдер, Лесли Г.; Мако, Кэтлин А. (1 января 1983 г.). «Объектное зрение и пространственное зрение: два корковых пути» . Тенденции в нейронауках . 6 : 414–417. дои : 10.1016/0166-2236(83)90190-X . ISSN 0166-2236 . S2CID 15565609 .
- ^ Гудейл, Массачусетс; Милнер, AD; Якобсон, Л.С.; Кэри, ДП (январь 1991 г.). «Нейрологическая диссоциация между восприятием объектов и их схватыванием» . Природа . 349 (6305): 154–156. Бибкод : 1991Natur.349..154G . дои : 10.1038/349154a0 . ISSN 1476-4687 . ПМИД 1986306 . S2CID 4238254 .
- ^ Милнер, AD; Гудейл, Массачусетс (2006). Визуальный мозг в действии (2-е изд.). ОУП Оксфорд. ISBN 978-0198524724 .
- ^ Марк Ф. Медведь; Барри Коннорс; Майкл Парадизо (2007). Нейронаука: исследование мозга . Хагерстаун, Мэриленд: Липпинкотт Уильямс и Уилкинс. ISBN 978-0-7817-6003-4 .
- ^ Ламме, Виктор А.Ф.; Супер, Ганс; Спекрейсе, Хенк (1998). «Обработка прямой, горизонтальной и обратной связи в зрительной коре». Современное мнение в нейробиологии . 8 (4): 529–535. дои : 10.1016/S0959-4388(98)80042-1 . ПМИД 9751656 . S2CID 1499090 .
- ^ Козловский Станислав; Рогачев, Антон (2021). «Как взаимодействуют области вентрального зрительного потока, когда мы запоминаем информацию о цвете и форме» . Достижения в области интеллектуальных систем и вычислений . 1358 г. (95–100). Спрингер-Природа: 95–100. дои : 10.1007/978-3-030-71637-0_10 . ISBN 978-3-030-71636-3 . ISSN 2194-5357 . S2CID 234902744 .
- ^ «Ухудшение зрения вследствие поражения головного мозга» . www.ssc.education.ed.ac.uk .
- ^ Хикок, Грегори; Поппель, Дэвид (13 апреля 2007 г.). «Корковая организация обработки речи». Обзоры природы Неврология . 8 (5): 393–402. дои : 10.1038/nrn2113 . ISSN 1471-003X . ПМИД 17431404 . S2CID 6199399 .
- ^ ДеВитт, И.; Раушекер, JP (1 февраля 2012 г.). «Распознавание фонем и слов в слуховом вентральном потоке» . Труды Национальной академии наук . 109 (8): Е505–Е514. Бибкод : 2012PNAS..109E.505D . дои : 10.1073/pnas.1113427109 . ISSN 0027-8424 . ПМК 3286918 . ПМИД 22308358 .
- ^ Гоу, Дэвид В. (июнь 2012 г.). «Корковая организация лексических знаний: двойная лексическая модель обработки разговорной речи» . Мозг и язык . 121 (3): 273–288. дои : 10.1016/j.bandl.2012.03.005 . ISSN 0093-934X . ПМЦ 3348354 . ПМИД 22498237 .
- ^ Хикок, Грегори (2012). «Корковая организация обработки речи: управление обратной связью и прогнозирующее кодирование в контексте двухпоточной модели» . Журнал коммуникативных расстройств . 45 (6): 393–402. дои : 10.1016/j.jcomdis.2012.06.004 . ПМЦ 3468690 . ПМИД 22766458 .
- ^ Говард, Гарри. «Спинной поток» . Мозг и язык . Архивировано из оригинала 10 мая 2017 г. Проверено 5 декабря 2015 г.
- ^ Бухсбаум, Брэдли Р.; Бальдо, Джулиана; Окада, Кайоко; Берман, Карен Ф.; Дронкерс, Нина; Д'Эспозито, Марк; Хикок, Грегори (декабрь 2011 г.). «Кондуктивная афазия, сенсомоторная интеграция и фонологическая кратковременная память – совокупный анализ поражения и данные фМРТ» . Мозг и язык . 119 (3): 119–128. дои : 10.1016/j.bandl.2010.12.001 . ISSN 0093-934X . ПМК 3090694 . ПМИД 21256582 .
- ^ Jump up to: а б Говард, Гарри. «Сенсомоторный интерфейс» . Мозг и язык .
- ^ Милнер, AD; Гудейл, Массачусетс. (февраль 2008 г.). «Пересмотр двух зрительных систем». Нейропсихология . 46 (3): 774–85. doi : 10.1016/j.neuropsychologia.2007.10.005 . ПМИД 18037456 . S2CID 1475566 .
- ^ Франц В.Х., Гегенфуртнер К.Р., Бюльтхофф Х.Х., Фале М. (2000). «Захват зрительных иллюзий: нет доказательств диссоциации между восприятием и действием». Психол. Наука . 11 (1): 20–5. дои : 10.1111/1467-9280.00209 . ПМИД 11228838 . S2CID 6461170 .
- ^ Франц В.Х., Шарновски Ф., Гегенфуртнер К.Р. (2005). «Эффект иллюзии при хватании постоянен во времени, а не динамичен». J Exp Psychol Hum Percept Performance . 31 (6): 1359–78. дои : 10.1037/0096-1523.31.6.1359 . ПМИД 16366795 .
- ^ Ганель Т., Гудейл Массачусетс (2003). «Визуальный контроль действия, а не восприятия, требует аналитической обработки формы объекта». Природа . 426 (6967): 664–7. Бибкод : 2003Natur.426..664G . дои : 10.1038/nature02156 . PMID 14668865 . S2CID 4314969 .
- ^ Ганель Т., Танзер М., Гудейл М.А. (2008). «Двойная диссоциация между действием и восприятием в контексте зрительных иллюзий: противоположные эффекты реального и иллюзорного размера». Психол. Наука . 19 (3): 221–5. дои : 10.1111/j.1467-9280.2008.02071.x . ПМИД 18315792 . S2CID 15679825 .
- ^ Кардозо-Лейте, Педро; Горя, Андрей (2010). «О перцептивной/моторной диссоциации: обзор концепций, теории, экспериментальных парадигм и интерпретации данных». Видение и восприятие . 23 (2): 89–151. дои : 10.1163/187847510X503588 . ПМИД 20550823 .
- ^ Гудейл М.А. (2011). «Превращение видения в действие» . Видение Рез . 51 (14): 1567–87. дои : 10.1016/j.visres.2010.07.027 . ПМИД 20691202 .
- ^ Гессен, К.; Болл, К.; Шенк, Т. (январь 2012 г.). «Визуомоторные функции, основанные на периферийном зрении, нарушены у пациента DF без зрительной формы». Нейропсихология . 50 (1): 90–7. doi : 10.1016/j.neuropsychologia.2011.11.002 . ПМИД 22085864 . S2CID 3092864 .
- ^ Jump up to: а б Макинтош, доктор медицинских наук; Шенк, Т. (май 2009 г.). «Два визуальных потока для восприятия и действия: современные тенденции». Нейропсихология . 47 (6): 1391–6. doi : 10.1016/j.neuropsychologia.2009.02.009 . ПМИД 19428404 . S2CID 32937236 .
- ^ Милнер, AD; Гудейл, Массачусетс (2006), Визуальный мозг в действии , ISBN 978-0-19-852472-4 , получено 6 декабря 2012 г.