Ортакантус
| Ортакантус | |
|---|---|
| |
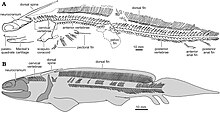
| Ископаемая интерпретация молодого экземпляра O. platypternus (вверху) и реконструкция скелета молодого экземпляра O. bohemicus (внизу) | |

| |
| Череп спереди, Американский музей естественной истории. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Хондриктиес |
| Подкласс: | Elasmobranchii |
| Заказ: | † Ксенакантида |
| Семья: | † Ортакантиды Хейлер и Поплин, 1989 г. |
| Род: | † Ортакантус Агассис , 1843 г. |
| Разновидность | |
| |
Orthacanthus — вымерший род пресноводных ксенакантиформных хрящевых рыб , названный Луисом Агассисом в 1843 году. [ 3 ] [ 4 ] начиная с верхнего карбона [ 3 ] в нижнюю пермь . [ 1 ] Ортакантус имел нектобентосную среду обитания с плотоядной диетой. [ 5 ] Многие авторы также обнаружили доказательства каннибализма в рационе ортакантуса и «сыновнего каннибализма», когда взрослые ортакантусы охотились на молодых ортакантусов. [ 6 ] Синонимы рода Orthacanthus : Dittodus Owen, 1867, Didymodus Cope, 1883, Diplodus Agassiz, 1843, [ 5 ] Хилодус Гибель, 1848 г. (занят Хилодусом Мюллером и Трошелем, 1844 г.). [ 3 ]
В позднем карбоне — начале перми ортакантус был главным хищником пресноводных болот и заливов Европы и Северной Америки. [ 6 ] Взрослый ортакантус достигал почти 3 метров (10 футов) в длину. [ 6 ] Зубы Orthacanthus имеют как минимум три бугра, два главных бугра и промежуточный бугор, где основные бугорки имеют различную зазубренность, со сложной морфологией основания. [ 7 ] [ 4 ] Кроме того, ортакантус можно диагностировать по большим поперечным осям проксимальных концов, расположенным под углом 45 градусов и часто почти параллельно лабиальному краю основания между бугорками. [ 4 ] Деформированные зубы характерны для ксенакантовых акул и ортакантусов. [ 8 ]
Открытие и история
[ редактировать ]
Два рода Orthacanthus и Pleuracanthus были выделены Луи Агассисом на основе изолированных «ихтиодорулитов» Британской каменноугольной системы и в то время ошибочно считались первыми индикаторами скатов. [ 9 ] Первоначально они были найдены в Соединенном Королевстве в Дадли, Лидсе, Северном Уэльсе, Карлуке и Эдинбурге. [ 9 ] Еще три вида из каменноугольной формации Огайо были описаны Джоном Стронгом Ньюберри, но два из них были младшими омонимами другого вида, Orthacanthus gracilis (Giebel, 1848). Соответственно, эти два вида получили замещающие названия: O. adamas Babcock 2024 и O. lintonensis Babcock, 2024. [ 3 ] Зубы, связанные с Diplodus , были найдены в каменноугольных сланцах Англии в Стаффорде, Карлуке и Бердихаусе, а также в Новой Шотландии. акулами [ 9 ] Хорошо сохранившийся отпечаток из Руппельсдорфа, Богемия, был описан Гольдфусом, а в отдельной статье тот же экземпляр был описан под названием Xenacanthus dechenii . Год спустя, в 1849 году, доктор Джордан ошибочно идентифицировал этот экземпляр как останки ископаемой акулы Triodus sessilis. идентифицировал этот экземпляр как Ксенакант . Эта ошибка была исправлена, и г-н Шнур [ 9 ]
Описание
[ редактировать ]Зубы
[ редактировать ]
Более крупные зубы Orthacanthus compressus и Orthacanthus texensis отличаются более выраженным базальным бугорком у O. compressus. [ 7 ] Базальный бугорок типичного зубного файла находится на апикальной бугорке нижележащего зуба. [ 8 ] Более крупные взрослые зубы O. compressus имеют более широкое, а не длинное основание, как и у O. texensis , и имеют тенденцию иметь зазубрины на обоих килях каждого бугорка, в то время как медиальные кили более мелких взрослых зубов не имеют зазубрин. [ 7 ] Ювенильные зубы O. compressus длиннее, чем ширина, имеют более тонкое основание и лишены зазубрин, как у зубов O. platypternus. [ 7 ]
Orthacanthus platypternus из слоя акул в Крэддоке в Техасе, США, демонстрирует признаки резорбции и является эквивалентом « эмалевой жемчужины ». [ 8 ] Некоторые образцы зубов, найденные в этом месте, демонстрируют признаки резорбции, которая ранее не наблюдалась у других представителей фауны в том же месте. [ 8 ] Если ожидается, что расположенный выше базальный бугорок будет резорбирован, если зубы подвергнутся резорбции, вместо этого резорбируется апикальная бугорок. [ 8 ]
Половой диморфизм
[ редактировать ]Разница в характеристиках больших и малых зубов взрослых особей O. compressus может указывать на половой диморфизм . [ 7 ]
Шипы O. platypternus, имеющие от 3 до 4 слоев дентина, интерпретируются как полувзрослые или молодые взрослые особи и делятся на два класса размеров, где у самок самые большие шипы по сравнению с самцами, что указывает на половой диморфизм. [ 10 ]
Дорсальный шип, дентин и зубцы
[ редактировать ]Дорсальные шипы Orthacanthus platypternus из костного слоя Крэддок в Техасе, США, сохраняют сильно васкуляризированную стенку, состоящую в основном из центробежно растущего дентина (наружного слоя стенки шипа) в последовательности растущих внутрь слоев дентина, выстилающих пульпу. полость. [ 10 ] Эти слои дентина, вероятно, откладываются периодически в соответствии с сезонными колебаниями температуры воды и наличия пищи. [ 10 ] Более конкретно, периодический характер отложений слоя дентина может быть обусловлен изменением отложения фосфата кальция в результате изменений температуры воды. [ 11 ] Шипы особей с 1-2 слоями дентина, вероятно, являются ювенильными и имеют наименьшие размеры, тогда как особи, имеющие как минимум 3-4 слоя дентина, относятся к двум отдельным размерным классам. [ 10 ] Поперечное сечение овальное вблизи отверстия полости пульпы, круглое/субтреугольное в дистальной части беззубчатой области и круглое в зубчатой области. [ 10 ] Пульпарная полость позвоночника заполнена кальцитом, кварцем и непрозрачными минералами. [ 10 ]
Затылочный отдел позвоночника и зубцы
[ редактировать ]
Шип вставляется в кожу поверхностно, где он растет и перемещается из глубокого положения в дерме, где формируется трабекулярный дентин , в поверхностное место, где формируется центробежно растущий пластинчатый дентин. [ 11 ] Количество зубчиков за годовой цикл варьируется в зависимости от скорости роста и представляет собой независимые элементы дермы, образованные дермальным сосочком и вторично прикрепленные дентином к собственно шипу. [ 11 ] Плотность зубцов также варьируется в зависимости от скорости роста затылочного отдела позвоночника. [ 11 ] Отношение длины зубчатого отдела к общей длине позвоночника меняется на протяжении онтогенеза . [ 11 ]
Классификация
[ редактировать ]
Зубы Orthacanthus texensis и Orthacanthus platypternus из костных пластов нижней перми Техаса и зубы Orthacanthus compressus из верхнего пенсильванского периода Небраски и бассейна Данкард центральных Аппалачей были использованы для определения происхождения O. texensis и O. platypternus. . [ 7 ] Было высказано предположение, что и O. texensis , и O. platypternus могут происходить от O. compressus , при этом ювенильные особенности O. compressus сохраняются во взрослых зубах O. platypternus посредством педоморфоза, а ювенильные особенности O. compressus зубов наблюдаются в зубах взрослых особей O. texensis. [ 7 ]
Таксон Mimia находится за пределами клады, содержащей две монофилетические сестринские группы. Первая монофилетическая стволовой группы сестринская группа определяет кладу, включающую хондрихтианы , которая визуализируется как сестринская группа Doliodus , и большую кладу, включающую кладодонтовых акул и Orthacanthus . «Кладодонтовые акулы» плюс Orthacanthus составляют две монофилетические сестринские группы: Orthacanthus и Cladodoides и Tamiobatis с одной стороны и Symmoriiformes с другой. Вторая монофилетическая сестринская группа характеризует корончатые хондрихтианы, которые содержат две монофилетические сестринские группы ( Euchondrocephali с одной стороны и Euselachii с другой). [ 12 ]
Следующая кладограмма основана на анализе Алана Праделя и его коллег, проведенном в 2011 году. [ 12 ]
Палеобиология
[ редактировать ]
В 2013 году был проведен анализ изотопного состава кислорода и стронция в зубах и шипах таксонов акул позднего карбона и ранней перми, чтобы сделать вывод о гидрохимии окружающей их воды, что способствовало разногласиям между облигатным пресноводным или эвригалинным диадромным образом жизни. [ 13 ] Фациальные интерпретации перми Северной Америки показали, что толерантность к солености ксенакантиформ была ограничена прибрежной морской средой, тогда как только Orthacanthus мог переносить солоноватую водную среду. [ 13 ] В исследовании, посвященном морфологии и гистологии спинных шипов Orthacanthus platypternus, также сообщается, что «Сравнительный анализ онтогенетических стадий зарегистрированных экземпляров O. platypternus и их распределения по разным фациям и локалитетам показывает, что этот вид был эвригалинным , диадромным с катадромный жизненный цикл, который строго регулировался полузасушливым и сезонно сухим тропическим климатом, затрагивавшим западную Пангею в начале Пермь. [ 10 ] «Анализ 2013 года предоставил доказательства, склоняющиеся к облигатному пресноводному образу жизни акул из варисканских европейских бассейнов, а неморские соотношения показали, что на формирование зубов повлияли метеоритные волны, обогащенные испарением. [ 13 ] Эвригалинная адаптация не была подтверждена в анализе 2013 года. [ 13 ]
Отношения Хищник-Жертва
[ редактировать ]Ортакантус и Триодус имеют отношения хищник-жертва, в которых Ортакантус охотился на Триодуса . [ 14 ] Черепные останки экземпляров Orthacanthus и Triodus из верхнего карбона в бассейне Пуэртольяно, Испания, свидетельствуют об этих отношениях хищник-жертва. [ 14 ] Многочисленные и хорошо сохранившиеся головные элементы Triodus были связаны с черепными остатками Orthacanthus , что объясняется включением затылочных шипов Triodus в ротовую полость Orthacanthus. [ 14 ] Дополнительным доказательством является сочетание одного шипа Orthacanthus со многими шипами Triodus , которые, вероятно, проникли в мягкие ткани и хрящи рта Orthacanthus, подобно современным акулам, которые питаются скатами, у которых шипы скатов были обнаружены внутри и вокруг щечные полости Carcharhinus, Galeocerdo, Negaprion и Sphyrna.
Исследование Orthacanthus копролитов из Канады, проведенное Аодханом О'Гогейном и др. выявили, что в трудные времена ортакантус, скорее всего, был каннибалом , поскольку зубы молодых ортакантусов были обнаружены в копролитах взрослых особей. [ 15 ] [ 16 ] На основании анализа копролитов и содержимого кишечника ортакантус имел диету, состоящую из актиноптеригиев, акантодий, двукрылых, ксенакантидов и четвероногих. [ 6 ] Также были подозрения в сыновнем каннибализме из-за присутствия молодых зубов Orthacanthus внутри копролита Orthacanthus . Кал Orthacanthus имеет спиралевидную форму из-за штопорообразной прямой кишки. [ 6 ]
Палеоэкология
[ редактировать ]Палеобиогеографическое распространение O. platypternus позволяет предположить онтогенетическое разделение местообитаний. [ 10 ] Теория онтогенетической ниши предсказывает, что люди могут менять свою среду обитания или рацион, чтобы поддерживать оптимальные темпы роста или улучшать компромисс между риском смертности и ростом. [ 17 ] В то время как более мелкие особи, вероятно, жили на мелководье, например, в небольших прудах и ручьях прибрежной равнины, более крупные особи, вероятно, жили в более глубоких водах, таких как речные и озерные (реки и озера) и окраинные морские районы. [ 10 ]
Самый старый известный экземпляр Orthacanthus , Diploduspromaticus, был найден в Нью-Брансуике, Канада, в нижнем девоне (эмсский период, ок. 407–393 миллиона лет назад). [ 5 ] Другие экземпляры были найдены в таких местах, как США, Великобритания, Польша и Франция. [ 5 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Хампе, О. «О зубном ряде Orthacanthus (Chondrichthyes, Xencanthida) верхнего карбона-нижней перми». Палаонт З 62.3-4 (1988).
- ^ Бэрд, Дональд; Крюк, Роберт В. (1986). «Алмазная угольная шахта в Линтоне, штат Огайо, и ее позвоночные пенсильванского возраста» . Журнал палеонтологии позвоночных . 6 (2): 174–190. Бибкод : 1986JVPal...6..174H . дои : 10.1080/02724634.1986.10011609 . JSTOR 4523086 .
- ^ Перейти обратно: а б с д Бэбкок, Луизиана (2024). « Замена названий для двух видов Orthacanthus Agassiz, 1843 г. (Chondrichthyes, Xenacanthiformes) и обсуждение Giebelodus Whitley, 1940 г., заменяющее название Chilodus Giebel, 1848 г. (Chondrichthyes, Xenacanthiformes), озабоченное Хилодусом Мюллером и Трошелем, 1844 г. (Actinopterygii, Characiformes)." " . ZooKeys (1188): 219–226. дои : 10.3897/zookeys.1188.108571 . ПМЦ 10790574 . ПМИД 38230382 .
- ^ Перейти обратно: а б с Джонсон, Гэри Д.; Тайер, Дэвид В. (2009). «Ранние пенсильванские Xenacanth Chondrichthyans из гор Свиссхельм, Аризона, США» . Acta Palaeontologica Polonica . 54 (4): 649–668. дои : 10.4202/app.2008.0051 . ISSN 0567-7920 . S2CID 54093505 .
- ^ Перейти обратно: а б с д «ПБДБ» . paleobiodb.org . Проверено 04 марта 2020 г.
- ^ Перейти обратно: а б с д и Гогайн, Аодхан О; Фалькон-Лэнг, Ховард Дж.; Карпентер, Дэвид К.; Миллер, Рэндалл Ф.; Бентон, Майкл Дж.; Пуфаль, Пейр К.; Рута, Марчелло; Дэвис, Томас Г.; Хиндс, Стивен Дж.; Стимсон, Мэтью Р. (11 августа 2016 г.). «Данные из: Сообщества рыб и четвероногих в градиенте солености от морской до солоноватой в пенсильванской (раннемосковской) формации Минто в Нью-Брансуике, Канада, и их палеоэкологические и палеогеографические последствия» (PDF) . Палеонтология . дои : 10.1111/пала.12249 . ISSN 1475-4983 . S2CID 132845922 .
- ^ Перейти обратно: а б с д и ж г Джонсон, Гэри Д. (2012). «Возможное происхождение ксенакантовых акул Orthacanthus texensis и Orthacanthus platypternus в нижней перми Техаса, США». Историческая биология . 24 (4): 369–379. Бибкод : 2012HBio...24..369J . дои : 10.1080/08912963.2012.669128 . ISSN 0891-2963 . S2CID 128880622 .
- ^ Перейти обратно: а б с д и Джонсон, Джорджия (2018). «Зубы Orthacanthus platypternus (Cope, 1883) (Chondrichthyes: Xenacanthiformes) и другие изолированные останки позвоночных из одного горизонта в ранней перми (артинском) костного пласта Крэддок, нижняя группа Клир-Форк, округ Бэйлор, Техас, США» . Акта Геологика Полоника . 68 (3). doi : 10.1515/agp-2018-0025 (неактивен 31 января 2024 г.). ISSN 0001-5709 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Перейти обратно: а б с д Люткен, Кристиан (1868). «II. — О Ксенаканте (Ортаканте) Дечении, Гольдфусе. Профессор Кнер из Вены» . Геологический журнал . 5 (50): 376–380. Бибкод : 1868GeoM....5..376L . дои : 10.1017/S0016756800205025 . ISSN 0016-7568 . S2CID 129946774 .
- ^ Перейти обратно: а б с д и ж г час я Бек, Кимберли; Солер-Хихон, Родриго; Карлуччи, Джесси; Уиллис, Раймонд (2014). «Морфология и гистология спинных шипов ксенакантидной акулы Orthacanthus platypternus из нижней перми Техаса, США: палеобиологические и палеоэкологические последствия» . Acta Palaeontologica Polonica . дои : 10.4202/app.00126.2014 . ISSN 0567-7920 . S2CID 53979047 .
- ^ Перейти обратно: а б с д и Солер-Хихон, Родриго (1999). «Затылочный шип Orthacanthus (Xenacanthidae, Elasmobranchii): строение и рост». Журнал морфологии . 242 (1): 1–45. doi : 10.1002/(sici)1097-4687(199910)242:1<1::aid-jmor2>3.0.co;2-9 . ISSN 0362-2525 . ПМИД 10493780 . S2CID 6208698 .
- ^ Перейти обратно: а б Прадел, Алан; Таффоро, Пол; Мэйси, Джон Г.; Жанвье, Филипп (27 сентября 2011 г.). «Новые палеозойские Symmoriiformes (Chondrichthyes) из позднего карбона Канзаса (США) и кладистический анализ ранних хондрихтиев» . ПЛОС ОДИН . 6 (9): e24938. Бибкод : 2011PLoSO...624938P . дои : 10.1371/journal.pone.0024938 . ISSN 1932-6203 . ПМК 3181253 . ПМИД 21980367 .
- ^ Перейти обратно: а б с д Фишер, Ян; Шнайдер, Йорг В.; Фойгт, Силке; Иоахимски, Майкл М.; Тихомирова, Марион; Тюткен, Томас; Гетце, Йенс; Бернер, Ульрих (2013). «Изотопы кислорода и стронция из зубов ископаемых акул: экологические и экологические последствия для позднепалеозойских европейских бассейнов». Химическая геология . 342 : 44–62. Бибкод : 2013ЧГео.342...44Ф . doi : 10.1016/j.chemgeo.2013.01.022 . ISSN 0009-2541 .
- ^ Перейти обратно: а б с Солер-Хихон, Родриго (1995). «Свидетельства взаимоотношений хищник-жертва у ксенакантовых акул верхнего карбона (Стефаниан C) из Пуэртольяно; Испания». Геобиос . 28 : 151–156. Бибкод : 1995Geobi..28..151S . дои : 10.1016/s0016-6995(95)80104-9 . ISSN 0016-6995 .
- ^ «Ученые только что узнали нечто тревожное о огромном древнем виде акул» . Бизнес-инсайдер .
- ^ «Раскопанные: акулы-людоеды забытой эпохи» .
- ^ Рамирес, Мэтью Д.; Авенс, Лариса; Семинофф, Джеффри А.; Гоше, Лиза Р.; Хеппелл, Селина С. (16 февраля 2017 г.). «Динамика роста молодых морских черепах, претерпевающих онтогенетический сдвиг среды обитания». Экология . 183 (4): 1087–1099. Бибкод : 2017Oecol.183.1087R . дои : 10.1007/s00442-017-3832-5 . ISSN 0029-8549 . ПМИД 28210809 . S2CID 225674 .





