Полиспорангиофит
| Полиспорангиофит Временной диапазон:
| |
|---|---|
| |
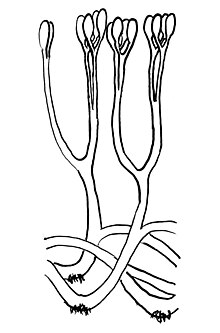
| Реконструкция аглаофитона с изображением раздвоенных осей с терминальными спорангиями и ризоидами. | |

| |
| Современный полиспорангиофит, папоротник монарх — сосудистое растение . | |
| Научная классификация | |
| Королевство: | Растения |
| Клэйд : | Эмбриофиты |
| Клэйд : | Полиспорангиофиты Кенрик и Крейн (1997) |
| Подгруппы | |
| |
Полиспорангиофиты , также называемые полиспорангиатами или формально Polysporangiophyta , представляют собой растения, у которых спороносное поколение ( спорофит ) имеет ветвящиеся стебли (оси), несущие спорангии . Название буквально означает «растение с множеством спорангиев». Клада (печеночников, мхов и роголистников), спорофиты которых обычно включает все наземные растения ( эмбриофиты ), за исключением мохообразных неразветвлены, даже если встречаются несколько исключительных случаев. [ 1 ] Хотя это определение не зависит от наличия сосудистой ткани , все живые полиспорангиофиты также имеют сосудистую ткань, т. е. являются сосудистыми растениями или трахеофитами. Известны вымершие полиспорангиофиты, не имеющие сосудистой ткани и поэтому не являющиеся трахеофитами.
Ранние полиспорангиофиты
[ редактировать ]История открытия
[ редактировать ]Палеоботаники различают микро- и мегаокаменелости. Микрофоссилии представляют собой в основном споры , одиночные или в группах. Мегаокаменелости — это сохранившиеся части растений, достаточно большие, чтобы можно было увидеть их структуру, например поперечное сечение стебля или структуру ветвления. [ 2 ]
Доусон , канадский геолог и палеоботаник, первым обнаружил и описал мегаокаменелость полиспорангиофита. В 1859 году он опубликовал реконструкцию девонского растения, собранного в качестве окаменелости в регионе Гаспе в Канаде, которое он назвал Psilophyton Princeps . Реконструкция показывает горизонтальные и вертикальные стеблеобразные конструкции; ни листьев, ни корней нет. пары спорообразующих органов ( спорангиев Прямостоячие стебли или оси ветвятся дихотомически и к ним прикреплены ). Поперечные сечения вертикальных осей показали сосудистой ткани наличие . Позже он описал другие экземпляры. Открытия Доусона изначально не имели большого научного значения; Тейлор и др. предполагают, что это произошло потому, что его реконструкция выглядела очень необычно, а окаменелость оказалась старше, чем ожидалось. [ 3 ]
Начиная с 1917 года, Роберт Кидстон и Уильям Х. Лэнг опубликовали серию статей, описывающих ископаемые растения из кремня Райни — мелкозернистой осадочной породы, найденной недалеко от деревни Райни в Абердиншире , которая теперь датируется пражским периодом нижнего девона (ок. 411–408 миллионов лет назад ). Окаменелости сохранились лучше, чем окаменелости Доусона, и ясно показали, что эти ранние наземные растения действительно состояли из обычно голых вертикальных стеблей, возникающих из аналогичных горизонтальных структур. Вертикальные стебли дихотомически разветвлены, некоторые ветви заканчиваются спорангиями. [ 3 ]
С момента этих открытий подобные мегаокаменелости были обнаружены в породах силурийского и среднего девона по всему миру, включая Арктическую Канаду, восточную часть США, Уэльс, Рейнскую область Германии, Казахстан, Синьцзян и Юньнань в Китае, а также Австралию. [ 4 ]
По состоянию на 2019 год [update], Eohostimella , датируемая эпохой Лландовери ( 444–433 миллиона лет назад ), является одной из самых ранних окаменелостей, идентифицированных как полиспорангиофит. [ 5 ] [ 6 ] Окаменелости, отнесенные к роду Cooksonia , который, скорее, является полиспорангиофитом, были датированы последующей эпохой Венлока ( от 433 до 427 миллионов лет назад ). [ 7 ] [ 8 ]
Таксономия
[ редактировать ]Концепция полиспорангиофитов, более формально называемых Polysporangiophyta, была впервые опубликована в 1997 году Кенриком и Крейном. [ 9 ] (Таксобокс справа представляет их взгляд на классификацию полиспорангиофитов.) Определяющей особенностью клады является то, что спорофит разветвляется и несет множественные спорангии. Это отличает полиспорангиофиты от печеночников , мхов и роголистников , у которых есть неразветвленные спорофиты, каждый из которых имеет один спорангий. Полиспорангиофиты могут иметь сосудистую ткань , а могут и не иметь ее ; те, у кого она есть, являются сосудистыми растениями или трахеофитами. [ нужна ссылка ]
До этого большинство ранних полиспорангиофитов были отнесены к одному отряду Psilophytales, классу Psilophyta , установленному в 1917 году Кидстоном и Лангом. [ 10 ] К классу, который тогда обычно назывался Psilopsida, иногда добавлялись современные Psilotaceae , папоротники-венчики. [ 11 ]
Когда были обнаружены и описаны дополнительные окаменелости, стало очевидно, что Psilophyta не являются однородной группой растений. В 1975 году Бэнкс расширил свое более раннее предложение 1968 года, разделив его на три группы по рангу подразделений. [ 12 ] [ 13 ] С тех пор к этим группам относились как к дивизиям, [ 14 ] сорт [ 15 ] и заказать. [ 16 ] Были использованы различные имена, которые суммированы в таблице ниже.
| Разделение | Подразделение | Сорт | Заказ | Неофициальный |
|---|---|---|---|---|
| Риниофиты | Риниофитина | Риниопсида (Rhyniophytopsida) [ 17 ] | Риниалес | риниофит |
| Зостерофиллофита | Зостерофиллофитина | Зостерофиллопсида | Зостерофиллалес | зостерофилл (зостерофиллофит) |
| Тримерофита (Trimerophytophyta) [ 18 ] | Тримерофитина (Trimerophytophytina) | Тримеропсида (Trimerophytopsida) | Тримерофиталес | тримерофит |
По Бэнксу, риниофиты включали простые безлистные растения с терминальными спорангиями (например, Cooksonia , Rhynia ) с центрархической ксилемой ; зостерофиллы включали растения с боковыми спорангиями, которые расщеплялись дистально (вдали от места прикрепления) для высвобождения спор, и имели экзарховые нити ксилемы (например, госслингии ). Тримерофиты включали растения с большими скоплениями изогнутых вниз концевых спорангиев, которые разделялись по длине для высвобождения спор и имели центральные тяжи ксилемы (например, Psilophyton ). [ 19 ]
Исследования Кенрика и Крейна, установившие полиспорангиофиты, пришли к выводу, что ни одна из трех групп Бэнкса не была монофилетической . Риниофиты включали «протрахеофиты», которые были предшественниками сосудистых растений (например, Horneophyton , Aglaophyton ); базальные трахеофиты (например, Stockmansella , Rhynia gwynne-vaughanii ); и растения, родственные линиям, которые привели к живым плаунам и их родственникам, а также папоротникам и семенным растениям (например, Cooksonia виды ). Зостерофиллы содержали монофилетическую кладу, но некоторые роды, ранее входившие в группу, выпали за пределы этой клады (например, Hicklingia , Nothia ). Тримерофиты были парафилетическими стеблевыми группами как для кроновой группы, папоротников кроновой группы так и для семенных растений . [ 20 ] [ 21 ]
Многие исследователи призывают к осторожности при классификации ранних полиспорангиофитов. Тейлор и др. Обратите внимание, что базальные группы ранних наземных растений по своей сути трудно охарактеризовать, поскольку они имеют много общих признаков со всеми более поздними группами (т. е. имеют множественные плезиоморфии ). [ 14 ] Обсуждая классификацию тримерофитов, Берри и Фейрон-Демаре говорят, что для достижения значимой классификации требуется «прорыв в знаниях и понимании, а не просто переосмысление существующих данных и окружающей мифологии». [ 22 ] Кладограммы Кенрика и Крейна были подвергнуты сомнению - см. Раздел «Эволюция» ниже.
По состоянию на февраль 2011 г. [update], по-видимому, не существует полной Линнеевской (т. е. ранговой) классификации ранних полиспорангиофитов, которая согласовывалась бы с кладистическим анализом Кенрика и Крейна и последующими исследованиями, хотя Cantino et al. опубликовали классификацию Филокодов . [ 23 ] Для удобства по-прежнему используются три группы банков. [ 14 ]
Филогения
[ редактировать ]Крупное кладистическое исследование наземных растений было опубликовано в 1997 году Кенриком и Крейном; это одновременно установило концепцию полиспорангиофитов и представило взгляд на их филогению . [ 9 ] С 1997 года наблюдается постоянный прогресс в понимании эволюции растений с использованием последовательностей генома РНК и ДНК и химического анализа окаменелостей (например, Taylor et al. 2006). [ 24 ] ), что привело к пересмотру этой филогении.
В 2004 году Крейн и др. опубликовали упрощенную кладограмму полиспорангиофитов (которые они называют полиспорангиатами), основанную на ряде цифр Кенрика и Крейна (1997). [ 10 ] Их кладограмма воспроизведена ниже (некоторые ветви свернуты в «базальные группы», чтобы уменьшить размер диаграммы). Их анализ не принимается другими исследователями; например, Ротвелл и Никсон говорят, что широко определенная группа папоротников (монилиформные или монилофиты) не является монофилетической. [ 25 ]
| полиспорангиофиты |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Совсем недавно Жерриенн и Гонез предложили несколько иную характеристику ранних дивергентных полиспорангиофитов: [ 26 ]
| Полиспорангиофиты |
| ||||||||||||
Парафилетические протрахеофиты, например Aglaophyton , имеют водопроводящие сосуды, как у мхов, т. е. без клеток с утолщенными клеточными стенками. Паратрахеофиты (название призвано заменить Rhyniaceae или Rhyniopsida) имеют водопроводящие клетки S-типа, т. е. клетки, стенки которых утолщены, но гораздо проще, чем у настоящих сосудистых растений, эвтрахеофитов. [ 26 ]
Эволюция
[ редактировать ]
Если приведенная выше кладограмма верна, это имеет значение для эволюции наземных растений. Самыми ранними дивергентными полиспорангиофитами в кладограмме являются Horneophytopsida , клада на уровне «протрахеофитов», которая является сестрой всех других полиспорангиофитов. По сути, у них наблюдалось изоморфное чередование поколений (это означает, что спорофиты и гаметофиты были одинаково свободноживущими), что может свидетельствовать о том, что как образ жизни мохообразных с доминированием гаметофитов, так и образ жизни сосудистых растений с доминированием спорофитов развились из этого изоморфного состояния. Они были безлистными и не имели настоящих сосудистых тканей. В частности, у них не было трахеид : удлиненных клеток, которые помогают транспортировать воду и минеральные соли и которые в зрелом возрасте образуют толстую одревесневшую стенку, обеспечивающую механическую прочность. В отличие от растений класса мохообразных , их спорофиты были разветвленными. [ 27 ]
Согласно кладограмме, род Rhynia иллюстрирует два этапа эволюции современных сосудистых растений. Растения имеют сосудистую ткань, хотя и значительно более простую, чем у современных сосудистых растений. Их гаметофиты заметно меньше спорофитов (но имеют сосудистую ткань, в отличие почти от всех современных сосудистых растений). [ 28 ]
Остальная часть полиспорангиофитов разделяется на две линии, глубокий филогенетический раскол, который произошел в раннем и среднем девоне, около 400 миллионов лет назад. Обе линии имеют листья, но разного типа. Ликофиты, составляющие менее 1% видов живых сосудистых растений, имеют небольшие листья ( микрофиллы или, точнее, ликофиллы), которые развиваются из интеркалярной меристемы (т. е. листья эффективно растут от основания). Эуфиллофиты - безусловно, самая многочисленная группа сосудистых растений как по численности, так и по видам. Эуфиллофиты имеют большие «настоящие» листья (мегафиллы), которые развиваются за счет краевых или апикальных меристем (т. е. листья эффективно растут по бокам или на верхушке). ( У хвоща вторично редуцированы мегафиллы, напоминающие микрофиллы.) [ 29 ]
И кладограмма, полученная на основе исследований Кенрика и Крейна, и ее значение для эволюции наземных растений были подвергнуты сомнению другими. В обзоре Генселя, проведенном в 2008 году, отмечается, что недавно обнаруженные ископаемые споры позволяют предположить, что трахеофиты присутствовали раньше, чем считалось ранее; возможно, раньше, чем предполагали члены стволовой группы . Разнообразие спор позволяет предположить, что существовало множество групп растений, другие остатки которых неизвестны. У некоторых ранних растений могла наблюдаться гетероморфная смена поколений с последующим приобретением изоморфных гаметофитов в определенных линиях. [ 30 ]
На кладограмме выше видно, что «протрахеофиты» расходятся раньше, чем ликофиты; однако ликофиты присутствовали на лудфордском этапе силура около 430–420 миллионов лет назад , задолго до того, как «протрахеофиты», обнаруженные в райнийских кремнях , датировались пражским этапом девона около 410 миллионов лет назад . [ 31 ] Однако было высказано предположение, что плохо сохранившаяся Eohostimella , обнаруженная в отложениях раннесилурийского возраста (Лландовери, около 440–430 миллионов лет назад ), может быть риниофитом. [ 6 ]
Бойс показал, что спорофиты некоторых видов Cooksonia и их родственников («куксониоиды») имели стебли, которые были слишком узкими, чтобы поддерживать достаточную фотосинтетическую активность и быть независимыми от своих гаметофитов, что несовместимо с их положением в кладограмме. [ 32 ]
Поскольку устьица мхов называемой , роголистников и полиспорангиофитов считаются гомологичными, было высказано предположение, что они принадлежат к естественной группе, стоматофитами . [ 33 ]
Эволюционная история растений далека от завершения. [ нужна ссылка ]
Примечания и ссылки
[ редактировать ]- ^ Харрисон, К. Джилл; Моррис, Дженнифер Л. (2017). «Происхождение и ранняя эволюция побегов и листьев сосудистых растений» . Философские труды Королевского общества B: Биологические науки . 373 (1739): 20160496. doi : 10.1098/rstb.2016.0496 . ПМЦ 5745332 . ПМИД 29254961 .
- ^ См., например, Эдвардс, Д. и Веллман, К. (2001), «Эмбриофиты на суше: запись от ордовика до лохкова (нижнего девона)» в Gensel & Edwards 2001 , стр. 3–28.
- ^ Перейти обратно: а б Тейлор, Теннесси; Тейлор, Э.Л. и Крингс, М. (2009), Палеоботаника, Биология и эволюция ископаемых растений (2-е изд.), Амстердам; Бостон: Академическая пресса, ISBN 978-0-12-373972-8 , с. 225 и далее
- ^ Гензель П.Г. и Эдвардс Д., ред. (2001), Растения вторгаются на землю: эволюционные и экологические перспективы , Нью-Йорк: издательство Колумбийского университета, ISBN. 978-0-231-11161-4 , главы 2, 6, 7
- ^ Эдвардс, Д. и Веллман, К. (2001), «Эмбриофиты на суше: от ордовика до лочковского (нижнего девона) периода», в Гензеле, П. и Эдвардсе, Д. (ред.), Растения вторгаются в землю: эволюционные и экологические перспективы , Нью-Йорк: издательство Колумбийского университета, стр. 3–28, ISBN. 978-0-231-11161-4 , с. 4
- ^ Перейти обратно: а б Никлас, Карл Дж. (1979), «Оценка химических свойств для классификации ископаемых растений», Taxon , 28 (5/6): 505–516, doi : 10.2307/1219787 , JSTOR 1219787
- ^ Эдвардс Д. и Фихан Дж. (1980), «Записи о спорангиях типа Cooksonia из слоев позднего Венлока в Ирландии», Nature , 287 (5777): 41–42, Бибкод : 1980Natur.287...41E , doi : 10.1038/287041a0 , S2CID 7958927
- ^ Либертин, Милан; Квачек, Иржи; Бек, Иржи; Жарский, Виктор и Шторх, Петр (2018), «Спорофиты полиспорангиальных наземных растений раннего силура, возможно, были фотосинтетически автономными» , Nature Plants , 4 (5): 269–271, doi : 10.1038/s41477-018-0140 -y , PMID 29725100 , S2CID 19151297
- ^ Перейти обратно: а б Кенрик и Крейн 1997a , стр. 139–140, 249.
- ^ Перейти обратно: а б Крейн, PR; Херендин, П. и Фриис, Э.М. (2004), «Ископаемые и филогения растений», Американский журнал ботаники , 91 (10): 1683–99, doi : 10.3732/ajb.91.10.1683 , PMID 21652317
- ^ Тейлор, Тейлор и Крингс 2009 , с. 226.
- ^ Бэнкс, HP (1968), «Ранняя история наземных растений», в книге Дрейка, ET (редактор), « Эволюция и окружающая среда: симпозиум, представленный по случаю 100-летия основания Музея естественной истории Пибоди в Йельском университете». Университет , Нью-Хейвен, Коннектикут: Издательство Йельского университета, стр. 73–107 , цитируется в Banks 1980.
- ^ Бэнкс, HP (1975), «Реклассификация Psilophyta», Taxon , 24 (4): 401–413, doi : 10.2307/1219491 , JSTOR 1219491.
- ^ Перейти обратно: а б с Тейлор, Тейлор и Крингс 2009 , с. 227
- ^ См., например, Берри, К.М. и Фэйрон-Демаре, М. (2001), «Возвращение к флоре среднего девона», в Gensel & Edwards 2001 , стр. 120–139.
- ^ Бэнкс, HP (1970), Эволюция и растения прошлого , Лондон: Macmillan Press, ISBN 978-0-333-14634-7 , с. 57
- ^ Хотя это имя встречается в некоторых источниках, например, Нолл, Эндрю Х. (1998-01-01), «Обзор происхождения и ранней диверсификации наземных растений: кладистическое исследование Пола Кенрика; Питера Крейна», Международный журнал наук о растениях , 159 (1): 172–174 , doi : 10.1086/297535 , JSTOR 2474949 , похоже, это ошибка, так как это не так. в соответствии со статьей 16 Международного кодекса ботанической номенклатуры .
- ^ Название основано на роде Trimerophyton ; Статья 16.4 Международного кодекса ботанической номенклатуры позволяет фитона опускать часть перед -ophyta , -ophytina и -opsida .
- ^ Бэнкс, HP (1980), «Роль Psilophyton в эволюции сосудистых растений», Обзор палеоботаники и палинологии , 29 : 165–176, doi : 10.1016/0034-6667(80)90056-1
- ^ Кенрик, Пол и Крейн, Питер Р. (1997a), Происхождение и ранняя диверсификация наземных растений: кладистическое исследование , Вашингтон, округ Колумбия: Издательство Смитсоновского института, ISBN 978-1-56098-730-7
- ^ Кенрик, П. и Крейн, PR (1997b), «Происхождение и ранняя эволюция наземных растений», Nature , 389 (6646): 33–39, Бибкод : 1997Natur.389...33K , doi : 10.1038/37918 , S2CID 3866183
- ^ Берри, К.М. и Фэйрон-Демаре, М. (2001), «Возвращение к флоре среднего девона», в Gensel & Edwards 2001 , стр. 127
- ^ Кантино, Филип Д.; Джеймс А. Дойл; Шон В. Грэм; Уолтер С. Джадд; Ричард Г. Олмстед; Дуглас Э. Солтис; Памела С. Солтис; Майкл Дж. Донохью (2007), «К филогенетической номенклатуре Tracheophyta», Taxon , 56 (3): 822–846, doi : 10.2307/25065865 , JSTOR 25065865
- ^ Тейлор, Д.В.; Ли, Хунци; Даль, Джереми; Фаго, Ф.Дж.; Зиннекер, Д.; Молдован, Дж. М. (2006), «Биогеохимические доказательства присутствия молекулярного ископаемого олеанана покрытосеменных растений в палеозойских и мезозойских не покрытосеменных окаменелостях» , Paleobiology , 32 (2): 179–90, doi : 10.1666/0094-8373(2006)32[179:BEFTPO]2.0.CO;2 , ISSN 0094-8373 , S2CID 83801635
- ^ Ротвелл, Г.В. и Никсон, К.К. (2006), «Как включение ископаемых данных меняет наши выводы о филогенетической истории эвфиллофитов?», International Journal of Plant Sciences , 167 (3): 737–749, doi : 10.1086/ 503298 , S2CID 86172890
- ^ Перейти обратно: а б Герриенн, П. и Гонез, П. (2011), «Ранняя эволюция жизненных циклов эмбриофитов: акцент на ископаемых доказательствах размера гаметофита/спорофита и морфологической сложности», Journal of Systematics and Evolution , 49 : 1–16, дои : 10.1111/j.1759-6831.2010.00096.x , S2CID 29795245
- ^ Бейтман, РМ; Крейн, PR; Димишель, Вашингтон; Кенрик, PR; Роу, Северная Каролина; Спек, Т.; Штейн, WE (1998), «Ранняя эволюция наземных растений: филогения, физиология и экология первичной земной радиации», Annual Review of Ecology and Systematics , 29 (1): 263–92, doi : 10.1146/annurev.ecolsys .29.1.263 , S2CID 44508826 , с. 270
- ^ Керп, Х.; Тревин, Нью-Хэмпшир; Хасс, Х. (2004), «Новые гаметофиты из раннедевонских рини-чертов», Труды Королевского общества Эдинбурга: Науки о Земле , 94 (4): 411–28, doi : 10.1017/s026359330000078x , S2CID 128629425
- ^ Прайер, К.М.; Шуэттпельц, Э.; Вольф, П.Г.; Шнайдер, Х.; Смит, Арканзас; Крэнфилл, Р. (2004), «Филогения и эволюция папоротников (монилофитов) с акцентом на ранние расхождения лептоспорангиатов», American Journal of Botany , 91 (10): 1582–98, doi : 10.3732/ajb.91.10.1582 , ПМИД 21652310 , стр. 1582–3
- ^ Гензель, Патрисия Г. (2008), «Самые ранние наземные растения», Annu. Преподобный Экол. Эвол. Сист. , 39 : 459–77, doi : 10.1146/annurev.ecolsys.39.110707.173526 , стр. 470–2
- ^ Котык, МЭ; Бейсингер, Дж. Ф.; Гензель, П.Г.; де Фрейтас, Т.А. (2002), «Морфологически сложные макрофоссилии растений из позднего силура Арктической Канады», Am. Дж. Бот. , 89 (6): 1004–1013, doi : 10.3732/ajb.89.6.1004 , PMID 21665700
- ^ Бойс, К.К. (2008), «Насколько зеленой была Куксония ? Важность размера для понимания ранней эволюции физиологии линии сосудистых растений», Paleobiology , 34 (2): 179–194, doi : 10.1666/0094-8373( 2008)034[0179:HGWCTI]2.0.CO;2 , ISSN 0094-8373 , S2CID 36688488
- ^ Лигроне, Р.; Дакетт, Дж. Г.; Ренцалья, Канзас (2012). «Основные переходы в эволюции ранних наземных растений: бриологический взгляд» . Анналы ботаники . 109 (5): 851–71. дои : 10.1093/aob/mcs017 . ПМК 3310499 . ПМИД 22356739 .
Внешние ссылки
[ редактировать ]- Кладограмма. Архивировано 4 декабря 2012 г. в archive.today от Crane, Herendeen & Friis 2004.
Классификация Archaeplastida или Plantae sl |
|---|