Частый гребень
Зубной гребень (также называемый зубным гребнем или зубным гребнем ) — это зубная структура, встречающаяся у некоторых млекопитающих, содержащая группу передних зубов, расположенных таким образом, чтобы облегчить уход, подобно расческе для волос . Зубной гребень встречается у лемуриформных приматов (к которым относятся лемуры и лорисоиды ), землероек , колуго , даманов и некоторых африканских антилоп . Структуры развивались независимо у разных типов млекопитающих в результате конвергентной эволюции и различаются как по составу, так и по строению зубов. У большинства млекопитающих гребень образован группой зубов с небольшими промежутками между ними. Зубные гребни у большинства млекопитающих включают только резцы, тогда как у лемуриформных приматов они включают резцы и клыки , которые наклонены вперед в передней части нижней челюсти, за которыми следует клыкообразный первый премоляр . Зубные гребни колуго и даманов принимают другую форму: отдельные резцы имеют зазубрины, образуя несколько зубцов на зуб.
Зубная расческа обычно используется для ухода за шерстью . Вылизывая шерсть, животное проводит по ней зубной расческой, чтобы расчесать ее. Тонкие бороздки или полосы обычно прорезаются на зубах во время ухода за волосами и их можно увидеть по бокам зубов, если смотреть через сканирующий электронный микроскоп . Зубной гребень поддерживается в чистоте либо языком, либо, в случае лемуриформ, подъязычной частью , специальным «под языком». Зубной гребень может выполнять и другие функции, например, заготовку еды и выдавливание коры. Среди лемуриформ с раздвоенными метками лемуры и индрииды имеют более прочные зубные гребни для поддержки этих вторичных функций. У некоторых лемуров, таких как ай-ай , зубной гребень полностью утерян и заменен другим специализированным зубным рядом.
У лемуриформных приматов ученые использовали зубной гребень для интерпретации эволюции лемуров и их родственников. Считается, что они произошли от ранних адаптивных приматов примерно в эоцене или раньше. Одна из популярных гипотез заключается в том, что они произошли от европейских адапидов , но летопись окаменелостей предполагает, что они произошли от более древней линии, которая мигрировала в Африку во время палеоцена (66–55 млн лет назад ) и, возможно, произошли от ранних церкамониинов из Азии. Ископаемые приматы, такие как Джебелемур , Anchomomys milleri и Плезиопитек , возможно, были их ближайшими родственниками. Отсутствие отчетливого зубного гребня в летописи окаменелостей до 40 млн лет назад создало конфликт с исследованиями молекулярных часов , которые предполагают более древнее расхождение между лемурами и лорисоидами, а также существование призрачной линии лемуриформных приматов в Африке.
Гомологичные и аналогичные структуры
[ редактировать ]
Зубной гребень, особое морфологическое расположение зубов в передней части нижней челюсти , наиболее известен у современных приматов-стрепсирринов , к которым относятся лемуры и лорисоидные приматы (известные под общим названием лемуриформные приматы). [ а ] ). [ 3 ] Эта гомологичная структура является диагностическим признаком, который помогает определить эту кладу (родственную группу) приматов . [ 4 ] [ 5 ] Аналогичная черта обнаружена у лысого уакари ( Cacajao Calvus ), разновидности обезьян Нового Света . [ 6 ]
Зубные гребни также можно найти у колуго и землероек, близких родственников приматов; однако структуры разные, и они считаются примерами конвергентной эволюции . [ 7 ] [ 8 ] малого или среднего размера Точно так же африканские антилопы , такие как импала ( Aepyceros melampus ), [ 9 ] имеют аналогичную структуру, которую иногда называют «аппаратом для бокового ухода за зубами». [ 10 ] [ 11 ] У живых и вымерших даманов (гиракоидов) также имеется зубной гребень, хотя количество зубцов в гребне варьируется в зависимости от летописи окаменелостей. [ 12 ]
Начиная с эпохи эоцена (более 50 млн лет назад) , Chriacus и Thryptacodon — два типа арктоционид (примитивные плацентарные млекопитающие ) — также обладали независимо развившимся зубным гребнем. [ 13 ] [ 14 ]
Анатомическое строение
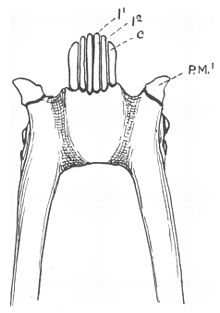
[ редактировать ]Зубной гребень большинства лемуриформ включает шесть мелко расположенных зубов, четыре резца и два клыка , лежащих (наклоненных вперед) в передней части рта. [ 4 ] [ 15 ] Стоящие нижние клыки имеют такую же форму, как и расположенные между ними резцы. [ 15 ] но они более крепкие и изогнуты вверх и внутрь сильнее, чем резцы. [ 13 ] В постоянном прикусе клыки прорезываются после резцов. [ 16 ] Коронки резцов также наклонены в сторону наклона вперед. [ 15 ] а коронки резцов и клыков удлинены и сжаты из стороны в сторону. [ 17 ] Верхушечный гребень, следующий по передним краям зубцов зубного гребня, у большинства лемуриформ имеет V-образную форму, сужается к средней линии. [ 18 ] В результате такой реконфигурации зубов верхние и нижние резцы не соприкасаются друг с другом, а часто верхние резцы редуцируются или полностью теряются. [ 15 ]

Французский анатом Анри Мари Дюкроте де Бленвиль впервые идентифицировал два боковых зуба лемуриформного зубного гребня как клыки в 1840 году. [ 19 ] Клыки обычно используются для прокалывания или захвата предметов. [ 20 ] При модифицированных нижних клыках первые нижние премоляры , следующие за зубным гребнем, обычно имеют форму типичных клыков (собачьи). [ 4 ] [ 21 ] и взять на себя их функцию. [ 15 ] Эти премоляры обычно путают с клыками. [ 22 ] В норме истинные клыки нижней челюсти располагаются впереди верхних, а у приматов с гребневидными зубами клыковидные премоляры лежат позади них. [ 15 ]
Лемуриформный зубной гребень поддерживается в чистоте подъязычной частью языка , или «под языком», специальной мышечной структурой, которая действует как зубная щетка, удаляя волосы и другой мусор. Подъязычная часть языка может простираться ниже конца языка и иметь ороговевшие зазубренные кончики, входящие между передними зубами. [ 23 ] [ 24 ] [ 25 ]
У лемуров зубной гребешок имеет изменчивое строение. [ 3 ] У индриид (Indriidae) зубной гребень менее выступающий. [ 26 ] и состоит из четырех зубов вместо шести. Зубной гребень индриидов более крепкий и широкий, с более короткими резцами, более широкими промежутками между зубами (межзубными промежутками) и более широким апикальным гребнем. [ 18 ] Неясно, состоит ли этот четырехзубый гребешок из двух пар резцов или из одной пары резцов и одной пары клыков. [ 4 ] [ 27 ] У лемуров с раздвоенными метками ( Phaner ) зубной гребень более сжат, межзубные промежутки значительно уменьшены. Все шесть зубов длиннее, прямее и образуют более непрерывный апикальный гребень. У недавно вымерших лемуров-обезьян (Archaeolemuridae) и лемуров-ленивцев (Palaeopropithecidae) зубной гребень был утрачен, а резцы и клыки вновь приобрели типичную конфигурацию в передней части рта. [ 28 ] Ай-ай также лишился зубного гребня, заменив его постоянно растущими (гипселодонтными) передними зубами, похожими на резцы грызунов . [ 4 ] [ 29 ]
У колуго зубной гребень имеет совершенно иное строение. Вместо того, чтобы отдельные резцы и клыки были расположены на мелком расстоянии друг от друга, чтобы действовать как зубья расчески, прикусная кромка четырех резцов стала зазубренной, с 15 зубцами каждый. [ 30 ] в то время как клык больше похож на коренной зуб. [ 31 ] Эти зазубренные резцы поддерживаются в чистоте с помощью передней части языка, которая имеет зазубрины, соответствующие зубцам резцов. [ 23 ] Точно так же зубной гребень гиракоида состоит из резцов с множеством зубцов, называемых « гребенками ». В отличие от колуго размер и форма зубцов более однородны. [ 12 ]
Зубной гребень землеройки похож на лемуриформный зубной гребень тем, что для образования зубцов гребня он использует межзубные промежутки, но в состав зубного гребня входят только две из трех пар нижних резцов. [ 8 ] [ 30 ] [ 32 ] и клыки также исключены. [ 30 ] Два боковых резца зубного гребня обычно крупнее. [ 8 ] У вымерших арктоционид все шесть нижних резцов входили в состав зубного гребня. [ 30 ] Зубной гребень африканских антилоп поразительно похож на гребень лемуриформ, поскольку состоит из двух пар резцов и пары клыков. [ 10 ]
Функции
[ редактировать ]Зубной гребень, являющийся гомологичной структурой у лемуриформ, выполняет различные биологические роли, несмотря на свою внешне стереотипную форму и внешний вид. [ 5 ] В основном он используется в качестве туалетного приспособления или расчески для ухода. [ 33 ] [ 34 ] Кроме того, некоторые виды используют зубные гребни для добычи пищи. [ 33 ] или выдолбить кору дерева. [ 34 ]
Уход
[ редактировать ]
Основная функция зубного гребня — уход за собой — впервые была отмечена французским натуралистом Жоржем Кювье в 1829 году, который указал, что у кошачьего лемура ( Lemur catta ) нижние резцы « sont de véritables peignes » («являются настоящими гребешками»). ). [ 31 ] Более 100 лет спустя функция ухода за шерстью была поставлена под сомнение, поскольку ее было трудно наблюдать, а межзубные промежутки считались слишком маленькими для шерсти. [ 35 ] Позже наблюдения показали, что зубы использовались для этой цели и что сразу после ухода волосы могут оказаться застрявшими в зубах, но позже они удаляются подъязычной частью. [ 36 ]
В 1981 году сканирующая электронная микроскопия выявила на зубах лемуриформных зубных гребней мелкие бороздки или полосы. Эти бороздки были обнаружены только по бокам зубов, на вогнутых поверхностях между сторонами, а также на заднем гребне зубов. Эти бороздки шириной от 10 до 20 мкм указывают на то, что волосы неоднократно перемещались по зубам. [ б ] Внутри этих борозд находились еще более тонкие бороздки, менее 1 мкм, образовавшиеся в результате истирания кутикулярного слоя волоса. [ 14 ]
Среди неприматов у вымершего Chriacus имеются микроскопические бороздки на зубном гребне. [ 14 ] а вот филиппинский колуго ( Cynocephalus volans ) этого не делает. Обычно считается, что зубной гребень колуго функционирует как зубной гребень, но из-за отсутствия бороздок на зубах и отсутствия документированных наблюдений за использованием зубного гребня во время ухода за полостью рта его использование, по-видимому, ограничивается добычей пищи. [ 30 ]
У африканских антилоп боковой аппарат для ухода за зубами, по-видимому, не используется во время выпаса или просмотра . Вместо этого он используется во время ухода, когда голова совершает характерное движение вверх. Считается, что шерсть расчесывают и удаляют эктопаразитов . [ 10 ] [ 11 ]
Обоняние у лемуриформ
[ редактировать ]У лемуриформных приматов зубной гребень может также играть второстепенную роль в обонянии , что может быть причиной уменьшения размеров малоизученных верхних резцов. [ 40 ] [ 41 ] Зубная расческа может оказывать давление, стимулируя выделения желез, которые затем распространяются по шерсти. [ 42 ] Кроме того, уменьшение размера верхних резцов может привести к образованию щели между зубами (межрезцовая диастема), которая соединяет желобок ( расщелину в середине влажного носа или ринарий ) с сошниково-носовым органом в нёбе. Это позволит феромонам легче переноситься в сошниково-носовой орган. [ 41 ] [ 43 ]
Заготовка продуктов питания и другие виды использования
[ редактировать ]Мышиные лемуры ( Microcebus ), сифаки ( Propithecus ) и индри ( Indri ) используют свои зубные гребни, чтобы зачерпывать мякоть фруктов. [ 14 ] Другие мелкие лемуриформы, такие как лемуры с развилкой ( Phaner ), карликовый лемур с волосатыми ушами ( Allocebus ) и галаго (особенно роды Galago и Euoticus ), используют свои зубные гребни для соскабливания с зубов экссудатов растений, таких как камедь и сок . [ 14 ] [ 44 ] У лемуров с метками вилки зубной гребень специально приспособлен для минимизации застревания пищи, поскольку межзубные промежутки значительно уменьшены. [ 45 ] Травоядные также могут использовать свои зубные гребни для добычи колуго рода Cynocephalus пищи. [ 30 ]
Индрииды, такие как сифаки, используют свои зубные гребни, чтобы выдолбить кору или сухостой (кора-призинг). [ 14 ] [ 34 ] [ 44 ] это делается перед тем, как нанести ароматическую метку железой на груди. [ 46 ] Считается, что более прочная структура их зубного гребня помогает им противостоять сжимающим силам, испытываемым во время регулярного поднятия коры. [ 18 ]
Эволюция лемуриформ
[ редактировать ]Происхождение лемуриформного зубного гребня и характерной для него клады уже более столетия является центром серьезных дискуссий. В 1920 году британский палеоантрополог Уилфрид Ле Гро Кларк предположил, что зубной гребень, обнаруженный у землероек (которые, как он считал, были приматами), был ранней версией структуры зубов, обнаруженной у лемуриформ. Поскольку он считал, что ископаемые лорисоиды миоцена не полностью развили современный лемуриформный зубной гребешок, он предположил, что лемуры и лорисоиды развили этот признак независимо. Позже эта точка зрения была опровергнута, и монофилетическая связь между лемурами и лорисоидами. теперь признана [ 47 ]
Наследственное состояние переднего зубного ряда нижней челюсти, основанное на окаменелостях приматов эоцена, позволяет предположить, что у самых ранних приматов не было дифференцированного зубного гребня. [ 48 ] У большинства ископаемых стрепсирринов отсутствовал стереотипный лемуриформный зубной гребень. В совокупности ранние стреппсириновые приматы известны как адаптиформы . [ 49 ] Адапиформы считаются парафилетической группой (содержащей многих, но не всех потомков последнего общего предка членов группы), поскольку предполагается, что лемуриформы произошли от одной из нескольких групп адапиформ. [ 50 ] С точки зрения экологии, предполагается, что эволюция зубного гребня потребовала фоливорной (листоядной) диеты среди предковой популяции адаптивных форм, поскольку это привело бы к отбору редуцированных резцов, что послужило бы экзаптацией ( признак, имеющий адаптивное значение для что-то иное, чем то, для чего оно было изначально выбрано), которое затем можно было использовать для личного или социального ухода . [ 34 ] Однако включение клыков в зубной гребень, должно быть, потребовало исключительных условий, поскольку у крупных лемуриформ клыкообразные премоляры вторично видоизменились, чтобы заменить утраченные. [ 51 ]
Популярная гипотеза о происхождении клады лемуриформ состоит в том, что они произошли от европейских адапидов, известных как адапиды . [ 52 ] У некоторых адапидов гребни нижних резцов и клыков совпадают, образуя функциональную единицу роста, и американский палеонтолог Филип Д. Джинджерич предположил, что это предвещало развитие лемуриформного зубного гребня. [ 53 ] Однако в летописи окаменелостей эоцена не обнаружено ни одного лемуриформного зубного гребня. [ 54 ] а европейские адаптированные нижние челюсти того времени не напоминали производное состояние, наблюдаемое у лемуриформ. [ 55 ]
В настоящее время считается, что лемуриформы развились в Африке, а самые ранние известные приматы-стрепсиррины из Африки - это азибииды из раннего эоцена. [ 49 ] которые, вероятно, произошли в результате очень ранней колонизации афро-арабской суши в палеоцене (66–55 млн лет назад). [ 56 ] Стеблевые лемуриформы, в том числе Djebelemur и Anchomomys milleri , были обнаружены в Африке, датируются периодом от 50 до 48 млн лет назад и сильно отличаются от европейских адаптиформ. [ 49 ] Однако у них отсутствует зубной гребень. [ 56 ] [ 57 ] Эти стеблевые лемуриформы предполагают раннее общее происхождение с церкамониинами из-за пределов Европы. [ с ] [ 60 ] Судя по большим выступающим нижним зубам, плезиопитек , ископаемый примат, обнаруженный в отложениях позднего эоцена в Фаюмской впадине в Египте , считается наиболее близким родственником лемуриформ. [ 61 ] [ 62 ] Вместе Djebelemur , Anchomomys milleri и Plesiopithecus считаются сестринскими таксонами (ближайшими родственниками) лемуриформных приматов. [ 56 ]
Несоответствия в свиданиях
[ редактировать ]Хотя стеблевые лемуриформы, такие как Джебелемур , возможно, были современниками родственных приматов с зубными гребнями примерно от 50 до 48 млн лет назад, скудные африканские окаменелости предполагают, что дифференциация зубных гребней произошла примерно от 52 до 40 млн лет назад, по мнению французского палеоантрополога Марка Годино . [ 49 ] Это противоречит оценкам молекулярных часов , сделанным эволюционным антропологом Анной Йодер и другими, которые предсказывают, что расхождение лемуров и лорисоидов датируется периодом от 61 до 90,8 млн лет назад. [ 63 ]
В 2001 году открытие Bugtilemur , ископаемого примата из Пакистана, датируемого олигоценом и первоначально считавшегося хейрогалеидным лемуром, еще больше поставило под сомнение теорию происхождения лемуров ; [ 64 ] однако позже было показано, что это тип адаптивного примата, а не лемура. [ 65 ]
Минимальная палеонтологическая оценка дивергенции лемуров и лорисоидов почти удвоилась, когда в 2000-х годах в северном Египте были сделаны дополнительные открытия стволового галагида ( Сахарагалаго ) и стеблевого или корончатого лорисоида ( Каранисия ), датируемых 37 и 40 млн лет назад соответственно. [ 66 ] [ 67 ] Каранисия — старейший ископаемый примат, обладающий отчетливым лемуриформным зубным гребнем. [ 68 ] Это, а также исследования других африканских адаптиформ, таких как Anchomomys milleri , позволяют предположить более древнюю призрачную линию лемуриформ в Африке. [ 56 ] [ 69 ]
Оригинальная функция лемуровидного зубного гребня
[ редактировать ]Избирательное давление , которое сформировало оригинальный лемуриформный зубной гребень, было предметом серьезных дискуссий с 1970-х годов. Доказательства можно рассматривать как подтверждающие функцию ухода, функцию закупки продуктов питания или и то, и другое. [ 70 ] В начале 1900-х годов дискуссий было меньше. Уход считался основной функцией, поскольку у приматов нет когтей, необходимых для адекватного расчесывания шерсти, хотя полуобезьяньи приматы (стрепсиррины и долгопяты) в качестве компенсации обладают по крайней мере одним когтем для ухода на каждой ноге. [ 31 ] Уход за шерстью — в форме расчесывания меха — обычно считается основной функцией и первоначальной ролью лемуриформного зубного гребня, а последующие изменения в морфологии в различных линиях изменили его функцию и затмили его первоначальную функцию. [ 71 ]
Гипотеза о том, что зубной гребень эволюционировал для добывания пищи, была основана на наблюдениях за недавними таксонами лемуриформ , такими как хейрогалеидные лемуры (особенно лемуры с вилкообразными метками и волосатый ушастый карликовый лемур) и галаго, которые демонстрируют соскребание зубами экссудатов растений, а также как сифаки, практикующие кору. Каждый из них считался «примитивной» формой среди ныне живущих стрепсирин, что позволяет предположить, что первые лемуриформы демонстрировали похожее поведение. [ 44 ] Кроме того, было подчеркнуто сильное селективное давление со стороны экологии питания, оказываемое на передний зубной ряд, на основе специализированного верхнего переднего зубного ряда, наблюдаемого у недавно вымерших лемуров-коал ( Megaladapis ). Если экология питания могла иметь такое глубокое влияние на форму переднего зубного ряда, то конвергентная эволюция могла бы объяснить сходство, наблюдаемое между сжатыми нижними резцами лемуриформного зубного гребня и адаптацией к питанию экссудатом у рода Callithrix (тип игрунки ). [ 72 ]
Напротив, гипотеза ухода подчеркивала, что все лемуриформы используют свои зубные гребни для ухода за собой, а длинные тонкие зубы плохо приспособлены к механическому стрессу, связанному с выдавливанием и питанием экссудатом. [ 72 ] Кроме того, межзубные промежутки, наблюдаемые у большинства лемуриформ, способствуют расчесыванию шерсти, а также способствуют росту бактерий и разрушению зубов, если их использовать для питания экссудатом. В подтверждение этого у лемуриформ, питающихся экссудатом, обнаружено уменьшение межзубного расстояния. [ 73 ] Кроме того, клыки, входящие в состав зубного гребня, обеспечивают дополнительное межзубное пространство для расчесывания шерсти. [ 74 ] Даже поведение молодых лемуриформ позволяет предположить, что уход за ними играет более важную роль в использовании зубного гребня, чем добывание пищи. [ 75 ]
Примечания
[ редактировать ]- ^ Хотя монофилетические отношения между лемурами и лорисоидами широко признаны, название их клады - нет. Термин «стрепсиррин» использовался в качестве таксона мусорной корзины для приматов, не относящихся к галлоринам , большинство из которых не имеют характерного зубного гребня, а термин «лемуриформ» был проблематичным из-за его исторической принадлежности к живым и ископаемым приматам, включая малагасийских приматов. лемуры и европейские адапиды , но исключили лорисоидов. [ 1 ] Термин «лемуриформные» используется здесь, потому что он происходит от одной популярной таксономии, которая объединяет приматов с зубными гребнями в один инфраотряд , а вымерших адапиформ без зубных гребней - в другой, оба относятся к подотряду Strepsirrini. [ 2 ]
- ^ Основываясь на образцах зубного микроизноса, характерных для расчесывания зубов, а также на сходстве между расширенными в боковом направлении передними резцами и внешним видом латерального клыка в лемуровидном зубном гребне, американский физический антрополог Джеффри Х. Шварц предположил, что долгопяты обладают очень высокой производный зубной гребень, гомологичный таковому у лемуриформ. [ 37 ] Тем не менее, исследования зубных рядов долгопятов показали, что только некоторые виды долгопятов имеют характерные полоски на резцах, однако все исследованные виды, как сообщается, использовали передние зубы при уходе за шерстью, что позволяет предположить, что на наличие или отсутствие полосатости может влиять частота ухода за шерстью, шерсть. Толщина или межзубное расстояние. [ 38 ] Расчесывание зубов (как функция зубов с гребешком или без него) также было зарегистрировано у ранних приматов позднего эоцена. Necrolemur , тип европейской микрохериды (близкие родственники долгопятовидных омомид ), не имел зубного гребня, но имел признаки микроизноса зубов, приписываемые расчесыванию зубов. [ 14 ] [ 39 ]
- ^ Cercamoniines были ранней ветвью приматов-стрепсирринов, населявших северное полушарие в раннем палеогене . [ 58 ] Они возникли в Азии и считаются подсемейством нотарктид , вымерших североамериканских приматов, напоминавших современных лемуров. [ 59 ]
Ссылки
[ редактировать ]Цитаты
[ редактировать ]- ^ Расмуссен и Некарис 1998 , стр. 252.
- ^ Картмилл 2010 , с. 15.
- ^ Перейти обратно: а б Куоццо и Ямасита 2006 , с. 68.
- ^ Перейти обратно: а б с д и Куоццо и Ямасита 2006 , с. 73.
- ^ Перейти обратно: а б Салай и Селигсон 1977 , с. 76.
- ^ Иглен 1980 , стр. 278 и 280.
- ^ Вуд Джонс 1918 , стр. 345–346.
- ^ Перейти обратно: а б с Иглен 1980 , стр. 279–280.
- ^ Муринг, Маккензи и Харт 1996 , стр. 970.
- ^ Перейти обратно: а б с Маккензи 1990 , с. 117.
- ^ Перейти обратно: а б Муринг, Маккензи и Харт 1996 , с. 966.
- ^ Перейти обратно: а б Стивенс и др. 2009 , с. 974
- ^ Перейти обратно: а б Иглен 1980 , с. 277.
- ^ Перейти обратно: а б с д и ж г Роуз, Уокер и Джейкобс 1981 , с. 583.
- ^ Перейти обратно: а б с д и ж Анкель-Саймонс 2007 , стр. 235.
- ^ Иглен 1980 , с. 281.
- ^ Осман Хилл 1953 , стр. 52–53.
- ^ Перейти обратно: а б с Салай и Селигсон 1977 , с. 78.
- ^ Иглен 1980 , с. 279.
- ^ Анкель-Саймонс 2007 , стр. 234.
- ^ Осман Хилл 1953 , с. 53.
- ^ Вуд Джонс 1918 , с. 345.
- ^ Перейти обратно: а б Вуд Джонс 1918 , с. 347.
- ^ Осман Хилл 1953 , с. 73.
- ^ Анкель-Саймонс 2007 , стр. 421–423.
- ^ Анкель-Саймонс 2007 , стр. 254.
- ^ Анкель-Саймонс 2007 , стр. 233.
- ^ Салай и Селигсон 1977 , стр. 78–79.
- ^ Анкель-Саймонс 2007 , стр. 257.
- ^ Перейти обратно: а б с д и ж Роуз, Уокер и Джейкобс 1981 , с. 585.
- ^ Перейти обратно: а б с Вуд Джонс 1918 , с. 346.
- ^ Саргис 2004 , с. 58.
- ^ Перейти обратно: а б Анкель-Саймонс 2007 , стр. 238.
- ^ Перейти обратно: а б с д Розенбергер 2010 , с. 291.
- ^ Робертс 1941 , с. 236.
- ^ Робертс 1941 , стр. 236–237.
- ^ Шварц 2003 , стр. 66–67.
- ^ Массер и Дагосто 1987 , стр. 33–34.
- ^ Шмид 1983 , стр. 6–8.
- ^ Куоццо и Ямасита 2006 , с. 72.
- ^ Перейти обратно: а б Куоццо и Ямасита 2006 , стр. 74–76.
- ^ Розенбергер и Штрассер 1985 , с. 73.
- ^ Ашер 1998 , с. 355.
- ^ Перейти обратно: а б с Салай и Селигсон 1977 , стр. 76–77.
- ^ Салай и Селигсон 1977 , с. 80.
- ^ Миттермайер и др. 2010 , с. 572.
- ^ Розенбергер и Штрассер 1985 , с. 74.
- ^ Годино 2006 , с. 455 .
- ^ Перейти обратно: а б с д Годино 2006 , с. 446.
- ^ Годино 2006 , стр. 446–447.
- ^ Салай и Селигсон 1977 , с. 77.
- ^ Таттерсолл 2006 , с. 9.
- ^ Годино 2006 , с. 447.
- ^ Расмуссен и Некарис 1998 , стр. 256–258.
- ^ Годино 2006 , стр. 455–456.
- ^ Перейти обратно: а б с д Зейферт и др. 2005 , с. 11400.
- ^ Годино 2006 , с. 452.
- ^ Борода 2004 , стр. 55–56.
- ^ Борода 2004 , стр. 51–52.
- ^ Годино 2006 , стр. 453–455.
- ^ Саймонс и Расмуссен 1994 , с. 9946.
- ^ Расмуссен и Некарис 1998 , стр. 261.
- ^ Годино 2006 , с. 458 .
- ^ Таттерсолл 2006 , стр. 8–9.
- ^ Годино 2006 , стр. 447–449.
- ^ Джагер и др. 2010 , с. 1095.
- ^ Зейферт и др. 2005 , с. 11396.
- ^ Таттерсолл 2006 , с. 7.
- ^ Seiffert, Simons & Attia 2003 , стр. 423–424.
- ^ Куоццо и Ямасита 2006 , стр. 73–74.
- ^ Салай и Селигсон 1977 , с. 75.
- ^ Перейти обратно: а б Розенбергер и Штрассер 1985 , с. 75.
- ^ Салай и Селигсон 1977 , стр. 80–81.
- ^ Куоццо и Ямасита 2006 , с. 74.
- ^ Иглен 1980 , с. 276.
Цитируемая литература
[ редактировать ]- Анкель-Саймонс, Ф. (2007). Анатомия приматов (3-е изд.). Академическая пресса. ISBN 978-0-12-372576-9 .
- Ашер, Р.Дж. (1998). «Морфологическое разнообразие анатомического стреппсирринизма и эволюция лемуриформного зубного гребня». Американский журнал физической антропологии . 105 (3): 355–367. doi : 10.1002/(SICI)1096-8644(199803)105:3<355::AID-AJPA5>3.0.CO;2-Q . ПМИД 9545077 .
- Борода, К. (2004). Охота на рассветную обезьяну: раскрытие происхождения обезьян, человекообразных обезьян и людей . Издательство Калифорнийского университета. ISBN 978-0-520-23369-0 .
- Картмилл, М. (2010). «Глава 2: Классификация и разнообразие приматов» . В Платте, М.; Газанфар, А. (ред.). Нейроэтология приматов . Издательство Оксфордского университета. стр. 10–30. ISBN 978-0-19-532659-8 .
- Куоццо, ФП; Ямасита, Н. (2006). «Глава 4: Влияние экологии на зубы современных лемуров: обзор зубной адаптации, функций и истории жизни». Ин Гулд, Л.; Заутер, М.Л. (ред.). Лемуры: экология и адаптация . Спрингер. стр. 67–96. ISBN 978-0-387-34585-7 .
- Иглен, Р.Х. (1980). «Гомология зубных гребней и функция зубных гребней у существующих стрепсиринов». Международный журнал приматологии . 1 (3): 275–286. дои : 10.1007/BF02692275 . S2CID 34472983 .
- Годино, М. (2006). «Происхождение лемуриформ, как видно из летописи окаменелостей». Фолиа Приматологическая . 77 (6): 446–464. дои : 10.1159/000095391 . ПМИД 17053330 . S2CID 24163044 .
- Джагер, Джей-Джей; Борода, КЦ; Чаймани, Ю.; Салем, М.; Бенамми, М.; Хлал, О.; Костер, П.; Билал, А.А.; Дюринер, П.; Шустер, М.; Валентин, X.; Марандат, Б.; Мариво, Л.; Метаис, Э.; Хаммуда, О.; Брюне, М. (2010). «Эпоха позднего среднего эоцена в Ливии дает начало самой ранней известной радиации африканских антропоидов» (PDF) . Природа . 467 (7319): 1095–1098. Бибкод : 2010Natur.467.1095J . дои : 10.1038/nature09425 . ПМИД 20981098 . S2CID 4431606 . Архивировано из оригинала (PDF) 8 марта 2013 г.
- Маккензи, А.А. (1990). «Аппарат для ухода за зубами жвачных животных». Зоологический журнал Линнеевского общества . 99 (2): 117–128. дои : 10.1111/j.1096-3642.1990.tb00564.x .
- Миттермайер, РА ; Луи, EE ; Ричардсон, М.; Швитцер, Дж.; и др. (2010). Лемуры Мадагаскара . Иллюстрировано С.Д. Нэшем (3-е изд.). Интернационал Консервации . ISBN 978-1-934151-23-5 . OCLC 670545286 .
- Муринг, М.; Маккензи, А.А.; Харт, Б.Л. (1996). «Уход за импалой: роль ухода за полостью рта в удалении клещей и влияние клещей на увеличение скорости ухода» (PDF) . Физиология и поведение . 59 (4–5): 965–971. дои : 10.1016/0031-9384(95)02186-8 . PMID 8778894 . S2CID 15051712 . [ постоянная мертвая ссылка ]
- Мюссер, Г.Г.; Дагосто, М. (1987). «Особенность Tarsius pumilus , карликового вида, эндемичного для горных мшистых лесов центрального Сулавеси» (PDF) . Американский музей Novitates (2867): 1–53. ISSN 0003-0082 .
- Осман Хилл, WC (1953). Сравнительная анатомия и систематика приматов I — Strepsirhini . Пабы Эдинбургского университета «Наука и математика», № 3. Издательство Эдинбургского университета. OCLC 500576914 .
- Расмуссен, Д.Т.; Некарис, КАИ (1998). «Эволюционная история лориобразных приматов» (PDF) . Фолиа Приматологическая . 69 (Приложение 1): 250–285. дои : 10.1159/000052716 . ПМИД 9595692 . S2CID 14113728 .
- Робертс, Д. (1941). «Зубной гребень лемуров» . Журнал анатомии . 75 (Часть 2): 236–238. ПМЦ 1252661 . ПМИД 17104854 .
- Роуз, К.Д.; Уокер, А.; Джейкобс, LL (1981). «Функция зубного гребня нижней челюсти у современных и вымерших млекопитающих». Природа . 289 (5798): 583–585. Бибкод : 1981Natur.289..583R . дои : 10.1038/289583a0 . ПМИД 7007889 . S2CID 4326558 .
- Розенбергер, Ал. (2010). «Глава 14: Адаптивный профиль против адаптивной специализации: окаменелости и мармелад в ранней эволюции приматов». В Берроузе, AM; Нэш, LT (ред.). Эволюция экссудативности у приматов . Спрингер. стр. 273–295. ISBN 978-1-4419-6660-5 .
- Розенбергер, Алабама; Штрассер, Э. (1985). «Происхождение зубных гребней: поддержка гипотезы ухода». Приматы . 26 : 73–84. дои : 10.1007/BF02389048 . S2CID 11191729 .
- Саргис, Э.Дж. (2004). «Новые взгляды на землероек: роль тупаид в надпорядковых отношениях приматов» (PDF) . Эволюционная антропология: проблемы, новости и обзоры . 13 (2): 56–66. дои : 10.1002/evan.10131 . S2CID 84815380 . Архивировано из оригинала (PDF) 11 декабря 2015 г.
- Шмид, П. (1983). «Передний зубной ряд Omomyiformes (приматов)». Фолиа Приматологическая . 40 (1–2): 1–10. дои : 10.1159/000156088 . ПМИД 6407925 .
- Шварц, Дж. Х. (2003). «Глава 3: Насколько близко сходство Тарсиуса с другими приматами?» . В Райте, ПК; Саймонс, Эл.; Гурски, С. (ред.). Долгопяты: прошлое, настоящее и будущее . Издательство Университета Рутгерса. стр. 50–96. ISBN 978-0-8135-3236-3 .
- Зейферт, ER; Саймонс, Эл.; Аттиа, Ю. (2003). «Ископаемые свидетельства древнего расхождения лори и галаго» . Природа . 422 (6930): 421–424. Бибкод : 2003Natur.422..421S . дои : 10.1038/nature01489 . ПМИД 12660781 . S2CID 4408626 .
- Зейферт, ER; Саймонс, Эл.; Райан, ТМ; Аттиа, Ю. (2005). «Дополнительные останки Wadilemur elegans , примитивного стволового галагида из позднего эоцена Египта» . Труды Национальной академии наук . 102 (32): 11396–11401. Бибкод : 2005PNAS..10211396S . дои : 10.1073/pnas.0505310102 . ПМЦ 1183603 . ПМИД 16087891 .
- Саймонс, Эл.; Расмуссен, Д.Т. (1994). «Замечательный череп Plesiopithecus teras (Primates, Prosimii) из эоцена Египта» . Труды Национальной академии наук . 91 (21): 9946–9950. Бибкод : 1994PNAS...91.9946S . дои : 10.1073/pnas.91.21.9946 . ПМК 44934 . ПМИД 7937923 .
- Стивенс, Нью-Джерси; О'Коннор, премьер-министр; Робертс, EM; Готфрид, доктор медицины (2009). «Гиракоид из позднеолигоценовой группы красных песчаников Танзании, Rukwalorax jinokitana (род. и зр. ноябрь)» (PDF) . Журнал палеонтологии позвоночных . 29 (3): 972–975. дои : 10.1671/039.029.0302 . S2CID 14894853 . Архивировано из оригинала (PDF) 10 декабря 2015 г.
- Салай, Ф.С.; Селигсон, Д. (1977). «Почему возникла зубная расческа из стрепспирина?». Фолиа Приматологическая . 27 (1): 75–82. дои : 10.1159/000155778 . ПМИД 401757 .
- Таттерсолл, И. (2006). «Глава 1: Происхождение малагасийских стрепспириновых приматов». Ин Гулд, Л.; Заутер, М.Л. (ред.). Лемуры: экология и адаптация . Спрингер. стр. 3–20. ISBN 978-0-387-34585-7 .
- Вуд Джонс, Ф. (1918). «Подъязычная и бахромчатая складка» . Журнал анатомии . 52 (4): 345–353. ПМЦ 1262842 . ПМИД 17103846 .