ЭРАП1
| ЭРАП1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | ERAP1 , A-LAP, ALAP, APPILS, ARTS-1, ARTS1, ERAAP, ERAAP1, PILS-AP, PILSAP, аминопептидаза 1 эндоплазматического ретикулума | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 606832 ; МГИ : 1933403 ; Гомологен : 140581 ; GeneCards : ERAP1 ; ОМА : ERAP1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||






Аминопептидаза 1 эндоплазматического ретикулума ( ERAP1 ) представляет собой фермент , который у человека кодируется ERAP1 геном . M1 Эта цинк- аминопептидаза участвует в пути процессинга и презентации антигена. ERAP1 в основном расположен в эндоплазматическом ретикулуме (ER), где он обрезает пептиды на их N-конце, адаптируя их для презентации молекулами MHC класса I (MHC-I). [ 5 ] .
Номенклатура
[ редактировать ]Исторические названия ERAP1, [ Комитет по номенклатуре генов HUGO ]:
- Аминопептидазный регулятор выделения рецептора фактора некроза опухоли 1 (TNFR1) (ARTS-1)
- Лейцинаминопептидаза, полученная из адипоцитов (A-LAP)
- Пуромицин-нечувствительная лейцилспецифическая аминопептидаза (PILS-AP)
- КИАА0525
- У мышей ER-аминопептидаза, связанная с процессингом антигена (ERAAP)
Биология / Функции
[ редактировать ]Эффективная презентация антигенных пептидов молекулами MHC класса I обеспечивает ключевой сигнал для адаптивных иммунных ответов цитотоксическими (CD8 + ) Т-лимфоциты . В «эндогенном» пути презентации антигена белки, синтезируемые клетками, подвергаются цитозольной деградации, и некоторые из их пептидных фрагментов транспортируются в ЭР, где пептиды подходящей длины загружаются на молекулы MHC класса I. В ER ERAP1 укорачивает более длинные пептиды до оптимальной длины для стабильного связывания с молекулами MHC класса I (рис. 1). ERAP1, как и другие компоненты APP, индуцируется гамма-интерфероном , цитокином, индуцирующим презентацию антигена. [ 6 ] ERAP1 предпочтительно обрезает N-удлиненные субстраты из 9–16 остатков до пептидов оптимального размера из 8–10 остатков. [ 7 ] Этот механизм «молекулярной линейки» уникален для ERAP1. ERAP1 также участвует в презентации внеклеточных антигенов по пути перекрестной презентации.

«Обрезка» ERAP1 модулирует репертуар пептидов, представленный молекулами MHC класса I, и тем самым формирует адаптивный иммунный ответ. [ 6 ] [ 8 ] [ 9 ] В мышиных моделях дефицит ERAAP (мышиного гомолога человеческого ERAP1) приводит к сильному изменению и повышению иммуногенности пептидного репертуара, представленного MHC-I. [ 10 ] Мышиные модели с генетическим дефицитом ERAAP сыграли важную роль в понимании роли обрезки пептидов в ER. [ 8 ] [ 11 ] [ 12 ]
Было предложено, что помимо обрезки пептидов в ЭР, ERAP1 выполняет дополнительные функции в зависимости от своего местоположения. ERAP1 может секретироваться во внеклеточное пространство в ответ на воспалительные стимулы, что может привести к активации иммунных клеток, таких как макрофаги или естественные клетки-киллеры , и усилению экспрессии провоспалительных цитокинов . [ 13 ]
Генетика/Клиническое значение
[ редактировать ]Джин / локация
[ редактировать ]Ген ERAP1 ( HGNC: 18173 ) расположен на длинном плече хромосомы 5 ( 5q15 ). Длина гена составляет ~47 КБ. [ 14 ] и содержит 20 экзонов и 19 интронов, [ 15 ] которые кодируют 9 различных вариантов сплайсинга. Кодирующая последовательность демонстрирует высокую степень консервативности среди плацентарных млекопитающих (идентичность >80%). последовательности 227 ортологов ERAP1, идентифицированных примерно у 200 видов. Доступны [ 16 ]
SNP
[ редактировать ]ERAP1 представляет собой полиморфный ген , который имеет множество однонуклеотидных вариантов (SNV), включая несколько распространенных миссенс-вариантов , которые изменяют аминокислотную последовательность ERAP1. Различные комбинации общих SNV в ERAP1 организуются в отдельные гаплотипы , которые кодируют разные изоформы белка, часто называемые « аллотипами ». [ 17 ] Аллотипы ERAP1 можно в общих чертах разделить на категории на основе их ферментативной активности в диапазоне от «высокой» до «низкой». [ 18 ] [ 19 ] Ферментативная активность ERAP1 зависит от распознавания пептида субстратом, о чем свидетельствует эффективность обрезки конкретных пептидных субстратов, значительно различающаяся для данного аллотипа. Как и в случае с аллотипом 10, аллотипом с плохой обрезкой, который, как было показано, в десять раз менее активен по сравнению с предковым аллотипом в гидролизе субстрата l-лейцин-7-амидо-4-метилкумарина (Leu-AMC). [ 19 ]
Ассоциация заболеваний
[ редактировать ]Генетические варианты и гаплотипы (т.е. аллотипы) ERAP1 связаны с широким спектром воспалительных состояний, инфекционных заболеваний и рака. В частности, ERAP1 является основным геном риска, выявленным в полногеномных исследованиях ассоциации воспалительных состояний, связанных с MHC-I (или «MHC-I-опатий»), включая анкилозирующий спондилит. [ 20 ] , болезнь Беше [ 21 ] , Бёрдшот Увеит [ 22 ] и псориаз . [ 23 ] В этих условиях ERAP1 часто находится в эпистазе с аллелем MHC-I первичного риска. [ 24 ] . Другие ассоциации заболеваний включают инсулинозависимый сахарный диабет и рассеянный склероз. [ 25 ] . Исторически ассоциации генов ERAP1 впервые были зарегистрированы при гипертонии. [ 26 ] . Новые данные связывают SNV ERAP1 с развитием рака и восприимчивостью к инфекционным заболеваниям [ 27 ] , такие как SNV ERAP1, которые изменяют устойчивость к инфекции вируса гриппа [ 28 ] .
Структура/Механизм
[ редактировать ]Структура
[ редактировать ]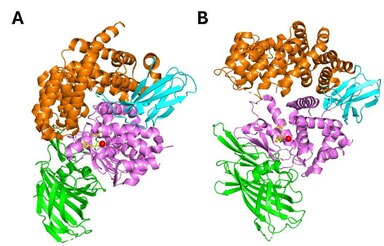
ERAP1 принадлежит к подсемейству окситоциназ семейства M1 металлопротеаз цинка . Он состоит из четырех структурных доменов. Домен I (остатки 1–254) состоит из восьминитевого β-листа и обеспечивает сайты связывания для N-конца субстратов. Он прилегает к каталитическому домену II и взаимодействует с доменом IV через удлиненную петлю. Домен II (остатки 255-529) представляет собой термолизин-подобный каталитический домен, состоящий из альфа-спирали и пятинитевого бета-листа. Этот лист содержит специфический для экзопептидаз мотив GAMEN, который создает одну часть щели для связывания субстрата. Каталитический атом Zn координируется остатками His353, His357 и Glu386, обнаруженными в цинксвязывающем мотиве (HEXXH-X18-E) на спирали 6a. Домен III (остатки 530–614) состоит из двух бета-листов, образующих бета-сэндвич, и действует как линкер между доменами II и IV. Наконец, домен IV (615–941) состоит преимущественно из а-спиралей и имеет чашеобразную форму. В закрытом (активном) состоянии он соприкасается с доменом II, образуя большую внутреннюю полость, в которой находится С-концевой сайт связывания субстрата (рис. 2А). Это наиболее вариабельный домен среди этого семейства аминопептидаз. [ 29 ] [ 30 ]
Механизм
[ редактировать ]ERAP1 использует каталитический механизм, аналогичный предложенному для гидролазы LTA4. [ 29 ] ERAP1 имеет термолизинподобную складку и кристаллизуется в двух различных конформациях: а. открытый и б. закрытый (рис. 2). В открытой конформации домен IV расположен вдали от активного центра, что делает внутреннюю полость более доступной для субстратов. В закрытой конформации внутренняя полость закрыта от внешнего растворителя и имеет достаточный размер для размещения пептида из 16 остатков. Каталитические остатки и, в частности, Tyr438 оптимально расположены для катализа в закрытой конформации. Следовательно, предполагается, что связывание субстрата происходит в открытой конформации, а разрыв N-концевой связи — в закрытой. [ 29 ] [ 30 ] Было высказано предположение, что связывание субстрата или небольших ингибиторов индуцирует конформационное закрытие ERAP1 в растворе. [ 31 ]
ERAP1 предпочитает пептидные субстраты длиной 9–16 аминокислот и гораздо менее активен для пептидов длиной 8–9 аминокислот. Считается, что ERAP1 использует механизм «молекулярной линейки», согласно которому субстрат связывается через свой гидрофобный С-конец в гидрофобном кармане на стыке домена III и домена IV, а N-конец связывается с активным центром. Когда длина пептида короче 8 или 9 аминокислот, пептид слишком короток, чтобы достичь активного центра, что ограничивает скорость расщепления. [ 7 ]
ERAP1 обладает широкой субстратной специфичностью с предпочтением гидрофобных остатков (например, лейцина и метионина) на N-конце пептидного субстрата. [ 32 ] [ 33 ] Триптофан, аргинин, цистеин и заряженные аминокислоты, такие как аспарагиновая и глутаминовая кислоты, удаляются плохо. [ 30 ] [ 32 ] На эффективность обрезки ERAP1 также может влиять внутренняя последовательность пептида с предпочтением гидрофобных и положительно заряженных остатков. [ 32 ]
Взаимодействия
[ редактировать ]Хисацунэ и др. продемонстрировали с помощью совместной иммунопреципитации взаимодействие между ERAP1 и дисульфид-перетасовывающим шапероном ERp44 , чему способствуют дисульфидные связи, образованные остатками цистеина в петле экзона 10 ERAP1. [ 34 ] Было предложено, чтобы это взаимодействие было основным механизмом удержания ER.
- Гетеродимеризация с ERAP2
Некоторые экспериментальные данные указывают на возможность образования гетеродимеров между ERAP1 и ERAP2, другим членом подсемейства окситоциназ аминопептидаз M1, который имеет структурное и функциональное сходство. Совместное элюирование ERAP1 и ERAP2 было обнаружено посредством фракционирования микросом во фракции 230 кДа, что указывает на образование гетеродимеров. [ 35 ] Анализ методом бесконтактного лигирования позволил предположить прямое физическое взаимодействие между двумя ферментами. [ 35 ] Комплекс ERAP1/ERAP2, опосредованный лейциновой застежкой, продемонстрировал повышенную эффективность обрезки по сравнению со смесью двух ферментов. [ 36 ] Вычислительная динамика показала, что гетеродимеризация ERAP1/ERAP2 может быть опосредована петлей экзона 10, [ 37 ] известно, что он участвует во взаимодействиях ERAP1-ERp44.
- МХК I предложил взаимодействие
Чен и др. предположили, что ERAP1 может обрезать N-концевые удлиненные антигенные пептиды-предшественники при связывании с MHC I. [ 38 ] Однако повторная оценка этой модели обрезки с помощью кинетического и биохимического анализа показала, что большинство связанных с MHC-I пептидов имели ограниченный доступ к активному сайту ERAP. [ 39 ]
Терапевтические подходы и фармакология
[ редактировать ]Терапевтические подходы к регуляции ERAP1 в основном основаны на разработке низкомолекулярных ингибиторов. Наиболее изученными классами ингибиторов ERAP1 являются ингибиторы каталитического или аллостерического сайта.
Ингибиторы каталитического сайта ERAP1
[ редактировать ]- Фосфиновые производные
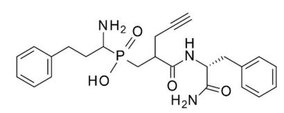
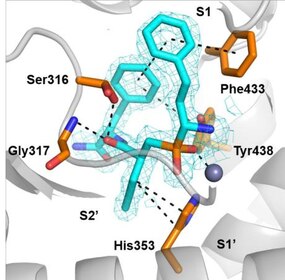
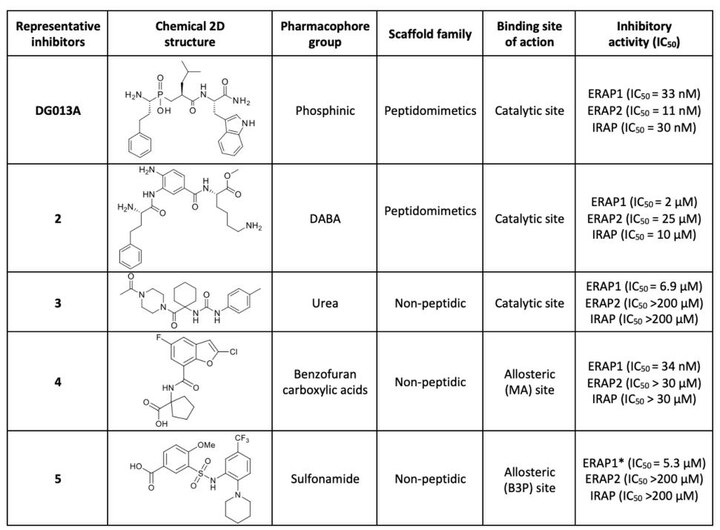
Первое поколение ингибиторов ERAP1 представляет собой серию фосфиновых псевдопептидов, полученных в результате рационального подхода к разработке, нацеленного на каталитический сайт связывания цинка, в 2013 году. 1 . Примечательно, что DG013A (таблица 1, рисунок 3) продемонстрировал высокую эффективность (IC 50 ERAP1 = 33 нМ), но плохую селективность в отношении ERAP2 и IRAP, при этом сообщалось об исследованиях по оптимизации SAR, демонстрирующих важность боковых цепей в положениях P 1 ' и P 2 '. [ 40 ] [ 41 ] Была получена кристаллическая структура высокого разрешения фосфинового аналога DG046, связанного с активным центром ERAP1 (рис. 4). [ 42 ]
- Аналоги ДАБА
Новое семейство соединений диаминобензойной кислоты (DABA), нацеленных на цинк, было рационально спроектировано и разработано в 2013 году, демонстрируя микромолярную эффективность ингибирования ERAP1 (соединение 2, IC 50 = 2 мкМ, таблица 1). Была достигнута умеренная селективность по сравнению с ERAP2 и IRAP с дополнительными усилиями по оптимизации, основанными на обширном исследовании SAR. [ 43 ] [ 44 ]
- Производные мочевины
(ВТС) было идентифицировано производное мочевины 3 (табл. 1) В 2020 г. с помощью высокопроизводительного скрининга как конкурентный ингибитор аминопептидазной активности ERAP1 (IC 50 = 6,9 мкМ) с повышенной селективностью по сравнению с ERAP2 и IRAP. [ 45 ] Исследования SAR и стыковки показали, что карбонильная группа N -ацетилпиперазина имеет решающее значение для активности благодаря своим свойствам цинксвязывающей группы. [ 45 ]
Ингибиторы аллостерического сайта ERAP1
[ редактировать ]- Бензофураны
Это семейство соединений было идентифицировано как потенциальный аллостерический (сайт распознавания С-концевого пептида) ингибитор посредством высокопроизводительного скрининга на основе флуоресценции в 2021 году. [ 46 ] Соединение 4 (таблица 1) проявляло высокую активность (IC 50 ERAP1 = 34 нМ) и в то же время селективность в отношении ERAP2 и IRAP. [ 46 ]
- Сульфонамиды
Сульфонамидное соединение 5 (таблица 1) было идентифицировано в ходе высокопроизводительных скрининговых исследований как потенциальный аллостерический селективный ингибитор, связывающийся на границе раздела между доменами II и IV ERAP1. Он активирует гидролиз небольших пептидов, но эффективно ингибирует процессинг длинных пептидов с 8–13 остатками (IC 50 = 5,3 мкМ) и проявляет селективность по сравнению с ERAP2 и IRAP. [ 45 ]
Клинические испытания
[ редактировать ]По состоянию на 2023 год ингибитор ERAP1 (GRWD5769), разработанный Gray Wolf Therapeutics, вступил в фазу I/II. Его безопасность, переносимость, эффективность и фармакокинетика оцениваются у пациентов с вирус-ассоциированными солидными опухолями (плоскоклеточный рак головы и шеи, рак шейки матки и гепатоцеллюлярная карцинома), которые особенно чувствительны к ингибированию ERAP1, в качестве монотерапии или в сочетании с ПД. -1 ингибитор иммунного контрольного пункта Либтайо ® (цемиплимаб). [ 47 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000164307 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000021583 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Шастри Н., Нагараджан Н., Линд К.К., Канасеки Т. (февраль 2014 г.). «Мониторинг процессинга пептидов молекул MHC класса I в эндоплазматическом ретикулуме» . Современное мнение в иммунологии . 26 : 123–127. дои : 10.1016/j.coi.2013.11.006 . ПМК 3932006 . ПМИД 24556408 .
- ^ Jump up to: а б Сарик Т., Чанг С.К., Хаттори А., Йорк И.А., Маркант С., Рок К.Л. и др. (декабрь 2002 г.). «Аминопептидаза, индуцированная ИФН-гамма в ЭР, ERAP1, удаляет предшественники пептидов, представленных MHC класса I». Природная иммунология . 3 (12): 1169–1176. дои : 10.1038/ni859 . ПМИД 12436109 .
- ^ Jump up to: а б Чанг С.К., Момбург Ф., Бутани Н., Голдберг А.Л. (ноябрь 2005 г.). «ER-аминопептидаза, ERAP1, урезает предшественники до длин пептидов MHC класса I с помощью механизма «молекулярной линейки» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (47): 17107–17112. Бибкод : 2005PNAS..10217107C . дои : 10.1073/pnas.0500721102 . ПМК 1287962 . ПМИД 16286653 .
- ^ Jump up to: а б Хаммер Дж.Э., Гонсалес Ф., Чампсаур М., Кадо Д., Шастри Н. (январь 2006 г.). «Аминопептидаза ERAAP формирует пептидный репертуар, отображаемый молекулами главного комплекса гистосовместимости класса I». Природная иммунология . 7 (1): 103–112. дои : 10.1038/ni1286 . ПМИД 16299505 .
- ^ Серволд Т., Гонсалес Ф., Ким Дж., Джейкоб Р., Шастри Н. (октябрь 2002 г.). «ERAAP настраивает пептиды для молекул MHC класса I в эндоплазматическом ретикулуме». Природа . 419 (6906): 480–483. Бибкод : 2002Natur.419..480S . дои : 10.1038/nature01074 . ПМИД 12368856 .
- ^ Хаммер Дж.Э., Гонсалес Ф., Джеймс Э., Нолла Х., Шастри Н. (январь 2007 г.). «В отсутствие аминопептидазы ERAAP молекулы MHC класса I представляют множество нестабильных и высокоиммуногенных пептидов». Природная иммунология . 8 (1): 101–108. дои : 10.1038/ni1409 . ПМИД 17128277 .
- ^ Фират Э., Савану Л., Айхеле П., Стаэхели П., Хуай Дж., Гаедике С. и др. (февраль 2007 г.). «Роль аминопептидазы 1, связанной с эндоплазматическим ретикулумом, в иммунитете к инфекции и в перекрестной презентации». Журнал иммунологии . 178 (4): 2241–2248. дои : 10.4049/jimmunol.178.4.2241 . ПМИД 17277129 .
- ^ Йорк И.А., Брем М.А., Зендзиан С., Таун К.Ф., Рок К.Л. (июнь 2006 г.). «Аминопептидаза 1 эндоплазматического ретикулума (ERAP1) урезает пептиды, представленные MHC класса I, in vivo и играет важную роль в иммунодоминировании» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (24): 9202–9207. Бибкод : 2006PNAS..103.9202Y . дои : 10.1073/pnas.0603095103 . ПМЦ 1482590 . ПМИД 16754858 .
- ^ Цудзимото М, Аоки К, Ониси А, Гото Ю (01 февраля 2020 г.). «Аминопептидаза 1 эндоплазматического ретикулума, помимо фермента, обрабатывающего антигенные пептиды, в эндоплазматическом ретикулуме» . Биологический и фармацевтический вестник . 43 (2): 207–214. дои : 10.1248/bpb.b19-00857 . ПМИД 32009107 .
- ^ «Аминопептидаза 1 эндоплазматического ретикулума ERAP1 [Homo sapiens (человек)]» .
- ^ Хаттори А., Мацумото К., Мизутани С., Цудзимото М. (август 2001 г.). «Геномная организация гена лейцинаминопептидазы, полученного из адипоцитов человека, и его связь с плацентарным геном лейцинаминопептидазы/окситоциназы». Журнал биохимии . 130 (2): 235–241. doi : 10.1093/oxfordjournals.jbchem.a002977 . ПМИД 11481040 .
- ^ «ERAP1 ENSG00000164307» .
- ^ Тран Т.М., Кольбер Р.А. (июль 2015 г.). «Аминопептидаза 1 эндоплазматического ретикулума и ревматические заболевания: функциональные вариации» . Современное мнение в ревматологии . 27 (4): 357–363. doi : 10.1097/BOR.0000000000000188 . ПМЦ 4495904 . ПМИД 26002027 .
- ^ Ривз Э., Джеймс Э. (сентябрь 2018 г.). «Роль полиморфного ERAP1 при аутовоспалительных заболеваниях» . Отчеты по биологическим наукам . 38 (4). дои : 10.1042/BSR20171503 . ПМК 6131210 . ПМИД 30054427 .
- ^ Jump up to: а б Хатчинсон Дж. П., Темпонерас И., Койпер Дж., Кортес А., Корчинска Дж., Китчен С. и др. (январь 2021 г.). «Общие аллотипы ER-аминопептидазы 1 обладают субстрат-зависимыми и сильно варьирующими ферментативными свойствами» . Журнал биологической химии . 296 : 100443. doi : 10.1016/j.jbc.2021.100443 . ПМК 8024916 . ПМИД 33617882 .
- ^ Эванс Д.М., Спенсер К.С., Пойнтон Дж.Дж., Су З., Харви Д., Кочан Г. и др. (июль 2011 г.). «Взаимодействие между ERAP1 и HLA-B27 при анкилозирующем спондилите предполагает использование пептидов в механизме HLA-B27 в предрасположенности к заболеванию» . Природная генетика . 43 (8): 761–767. дои : 10.1038/ng.873 . ПМК 3640413 . ПМИД 21743469 .
- ^ Кирино Ю., Берциас Г., Исигацубо Ю., Мизуки Н., Тугал-Туткун И., Сеяхи Э. и др. (февраль 2013 г.). «Общегеномный анализ ассоциаций идентифицирует новые локусы восприимчивости к болезни Бехчета и эпистазу между HLA-B*51 и ERAP1» . Природная генетика . 45 (2): 202–207. дои : 10.1038/ng.2520 . ПМЦ 3810947 . ПМИД 23291587 .
- ^ [Бесплатная статья PMC] [PubMed] Kuiper JJ, Setten JV, Devall M, Cretu-Stancu M, Hiddingh S, Ophoff RA и др. (декабрь 2018 г.). «Функционально разные ERAP1 и ERAP2 являются отличительной чертой увеита HLA-A29-(Birdshot)» . Молекулярная генетика человека . 27 (24): 4333–4343. дои : 10.1093/hmg/ddy319 . ПМК 6276832 . ПМИД 30215709 .
- ^ Стрэндж А., Кейпон Ф., Спенсер CC, Найт Дж., Уил М.Э., Аллен М.Х. и др. (ноябрь 2010 г.). «Полногеномное исследование ассоциации идентифицирует новые локусы предрасположенности к псориазу и взаимодействие между HLA-C и ERAP1» . Природная генетика . 42 (11): 985–990. дои : 10.1038/ng.694 . ПМЦ 3749730 . ПМИД 20953190 .
- ^ Лопес де Кастро Х.А., Альварес-Наварро К., Брито А., Гуасп П., Мартин-Эстебан А., Санс-Браво А. (сентябрь 2016 г.). «Молекулярные и патогенетические эффекты аминопептидаз эндоплазматического ретикулума ERAP1 и ERAP2 при воспалительных заболеваниях, связанных с MHC-I: на пути к единому взгляду». Молекулярная иммунология . 77 : 193–204. дои : 10.1016/j.molimm.2016.08.005 . ПМИД 27522479 .
- ^ Пепеляева Ю., Амальфитано А (май 2019 г.). «Роль ERAP1 в аутовоспалении и аутоиммунитете». Иммунология человека . 80 (5): 302–309. дои : 10.1016/j.humimm.2019.02.013 . ПМИД 30817945 .
- ^ Ямамото Н., Накаяма Дж., Ямакава-Кобаяши К., Хамагути Х., Миядзаки Р., Аринами Т. (март 2002 г.). «Идентификация 33 полиморфизмов в гене лейцинаминопептидазы (ALAP), полученного из адипоцитов, и возможная связь с гипертонией». Человеческая мутация . 19 (3): 251–257. дои : 10.1002/humu.10047 . ПМИД 11857741 .
- ^ Чифальди Л., Румыния П., Лоренци С., Локателли Ф., Фручи Д. (4 июля 2012 г.). «Роль аминопептидаз эндоплазматического ретикулума в здоровье и болезнях: от инфекции к раку» . Международный журнал молекулярных наук . 13 (7): 8338–8352. дои : 10.3390/ijms13078338 . ПМЦ 3430237 . ПМИД 22942706 .
- ^ Шотт Б.Х., Ван Л., Чжу X, Хардинг А.Т., Ко Э.Р., Буржуа Дж.С. и др. (ноябрь 2022 г.). «Объединение всего генома одной клетки показывает, что несинонимичный вариант ERAP1 обеспечивает повышенную восприимчивость к вирусу гриппа» . Клеточная геномика . 2 (11): 100207. doi : 10.1016/j.xgen.2022.100207 . ПМЦ 9718543 . PMID 36465279 .
- ^ Jump up to: а б с Кочан Г., Кройер Т., Харви Д., Фишер Р., Чен Л., Фоллмар М. и др. (май 2011 г.). «Кристаллические структуры аминопептидазы-1 эндоплазматического ретикулума (ERAP1) раскрывают молекулярную основу для обрезки N-концевого пептида» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (19): 7745–7750. Бибкод : 2011PNAS..108.7745K . дои : 10.1073/pnas.1101262108 . ПМЦ 3093473 . ПМИД 21508329 .
- ^ Jump up to: а б с Нгуен Т.Т., Чанг С.К., Евнучиду И., Йорк И.А., Зикос С., Рок К.Л. и др. (май 2011 г.). «Структурная основа процессинга предшественников антигенных пептидов аминопептидазой эндоплазматического ретикулума ERAP1» . Структурная и молекулярная биология природы . 18 (5): 604–613. дои : 10.1038/nsmb.2021 . ПМК 3087843 . ПМИД 21478864 .
- ^ Мабен З., Арья Р., Георгиадис Д., Стратикос Э., Стерн Л.Дж. (сентябрь 2021 г.). «Конформационная динамика, связанная с закрытием домена и связыванием субстрата, объясняет механизм аллостерической регуляции ERAP1» . Природные коммуникации . 12 (1): 5302. Бибкод : 2021NatCo..12.5302M . doi : 10.1038/s41467-021-25564-w . ПМЦ 8421391 . ПМИД 34489420 .
- ^ Jump up to: а б с Хирн А., Йорк, Айова, Рок КЛ (ноябрь 2009 г.). «Специфика обрезки пептидов, представленных MHC класса I, в эндоплазматическом ретикулуме» . Журнал иммунологии . 183 (9): 5526–5536. doi : 10.4049/jimmunol.0803663 . ПМЦ 2855122 . ПМИД 19828632 .
- ^ Евнушиду И., Момбург Ф., Папакириаку А., Хрони А., Леондиадис Л., Чанг С.К. и др. (06.11.2008). Эль-Шеми ХА (ред.). «Внутренняя последовательность пептида-субстрата определяет его обрезку N-конца с помощью ERAP1» . ПЛОС ОДИН . 3 (11): е3658. Бибкод : 2008PLoSO...3.3658E . дои : 10.1371/journal.pone.0003658 . ПМЦ 2573961 . ПМИД 18987748 .
- ^ Хисацуне С., Эбисуи Е., Усуи М., Огава Н., Сузуки А., Матага Н. и др. (июнь 2015 г.). «ERp44 осуществляет окислительно-восстановительный контроль артериального давления в отделении скорой помощи». Молекулярная клетка . 58 (6): 1015–1027. doi : 10.1016/j.molcel.2015.04.008 . ПМИД 25959394 .
- ^ Jump up to: а б Савану Л., Кэрролл О., Линдо В., Дель Валь М., Лопес Д., Лепеллетье Ю. и др. (июль 2005 г.). «Согласованная обрезка пептидов с помощью аминопептидазных комплексов ERAP1 и ERAP2 человека в эндоплазматическом ретикулуме». Природная иммунология . 6 (7): 689–697. дои : 10.1038/ni1208 . ПМИД 15908954 .
- ^ Евнушиду И., Ваймерсхаус М., Савану Л., ван Эндерт П. (июль 2014 г.). «Димеризация ERAP1-ERAP2 повышает эффективность обрезки пептидов». Журнал иммунологии . 193 (2): 901–908. doi : 10.4049/jimmunol.1302855 . ПМИД 24928998 .
- ^ Папакириаку А, Мпакали А, Стратикос Э (20 апреля 2022 г.). «Могут ли ERAP1 и ERAP2 образовывать функциональные гетеродимеры? Исследование структурной динамики» . Границы в иммунологии . 13 : 863529. дои : 10.3389/fimmu.2022.863529 . ПМЦ 9065437 . ПМИД 35514997 .
- ^ Чен Х., Ли Л., Ваймерсхаус М., Евнушиду И., ван Эндерт П., Бувье М. (август 2016 г.). «Димеры ERAP1-ERAP2 обрезают пептиды-предшественники, связанные с MHC I; значение для понимания редактирования пептидов» . Научные отчеты . 6 (1): 28902. Бибкод : 2016NatSR...628902C . дои : 10.1038/srep28902 . ПМЦ 4981824 . ПМИД 27514473 .
- ^ Мавридис Г., Арья Р., Домник А., Зойдакис Дж., Макридакис М., Влаху А. и др. (май 2020 г.). «Систематическое повторное исследование обработки предшественников антигенных пептидов, связанных с MHCI, аминопептидазой 1 эндоплазматического ретикулума» . Журнал биологической химии . 295 (21): 7193–7210. дои : 10.1074/jbc.RA120.012976 . ПМЦ 7247305 . ПМИД 32184355 .
- ^ Зервоуди Э., Саридакис Э., Бертли Дж.Р., Серегин С.С., Ривз Э., Коккала П. и др. (декабрь 2013 г.). «Рационально разработанный ингибитор, нацеленный на аминопептидазы, обрезающие антиген, усиливает презентацию антигена и цитотоксические реакции Т-клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (49): 19890–19895. Бибкод : 2013PNAS..11019890Z . дои : 10.1073/pnas.1309781110 . ПМЦ 3856834 . ПМИД 24248368 .
- ^ Коккала П., Мпакали А., Мове Ф.К., Папакириаку А., Даскалаки I, Петропулу I и др. (октябрь 2016 г.). «Оптимизация и взаимосвязь структура-активность фосфиновых псевдотрипептидных ингибиторов аминопептидаз, генерирующих антигенные пептиды». Журнал медицинской химии . 59 (19): 9107–9123. doi : 10.1021/acs.jmedchem.6b01031 . ПМИД 27606717 .
- ^ Гиастас П., Ной М., Роуленд П., Стратикос Э. (май 2019 г.). «Кристаллическая структура высокого разрешения аминопептидазы 1 эндоплазматического ретикулума со связанным фосфиновым аналоговым ингибитором переходного состояния» . Письма ACS по медицинской химии . 10 (5): 708–713. doi : 10.1021/acsmedchemlett.9b00002 . ПМК 6511960 . ПМИД 31097987 .
- ^ Папакириаку А., Зервуди Э., Теодоракис Э.А., Савану Л., Стратикос Э., Вурлумис Д. (сентябрь 2013 г.). «Новые селективные ингибиторы аминопептидаз, генерирующих антигенные пептиды» . Письма по биоорганической и медицинской химии . 23 (17): 4832–4836. дои : 10.1016/j.bmcl.2013.07.024 . ПМИД 23916253 .
- ^ Папакириаку А., Зервуди Е., Цукалиду С., Мове Ф.К., Сфироера Г., Мастеллос Д.С. и др. (февраль 2015 г.). «Производные 3,4-диаминобензойной кислоты как ингибиторы подсемейства окситоциназ аминопептидаз М1 с иммунорегулирующими свойствами» . Журнал медицинской химии . 58 (3): 1524–1543. дои : 10.1021/jm501867s . ПМИД 25635706 .
- ^ Jump up to: а б с Мабен З., Арья Р., Рэйн Д., Ан В.Ф., Меткар С., Хики М. и др. (январь 2020 г.). «Открытие селективных ингибиторов аминопептидазы 1 эндоплазматического ретикулума» . Журнал медицинской химии . 63 (1): 103–121. doi : 10.1021/acs.jmedchem.9b00293 . ПМЦ 8218592 . ПМИД 31841350 .
- ^ Jump up to: а б Деддуш-Грасс С., Андуш С., Беренц Ф., Холтер С., Хохвальд А., Лебрен Л. и др. (июль 2021 г.). «Открытие и оптимизация серии бензофурановых селективных ингибиторов ERAP1: биохимические исследования и исследования in silico » . Письма ACS по медицинской химии . 12 (7): 1137–1142. doi : 10.1021/acsmedchemlett.1c00235 . ПМК 8274102 . PMID 34267884 .
- ^ «Grey Wolf Therapeutics объявляет о дозировании первого пациента в фазе 1/2 клинического исследования GRWD5769 у пациентов с распространенными солидными опухолями» . 28 марта 2023 г.