гемозоин


Гемозоин — продукт утилизации, образующийся в результате переваривания крови некоторыми кровососущими паразитами . Эти гематофаги , такие как малярийные паразиты ( Plasmodium spp. ), Rhodnius и Schistosoma, переваривают гемоглобин и выделяют большое количество свободного гема , который является небелковым компонентом гемоглобина . Гем представляет собой простетическую группу, состоящую из атома железа, содержащегося в центре гетероциклического порфиринового кольца . Свободный гем токсичен для клеток, поэтому паразиты переводят его в нерастворимую кристаллическую форму, называемую гемозоином. У малярийных паразитов гемозоин часто называют малярийным пигментом .
Поскольку образование гемозоина необходимо для выживания этих паразитов, оно является привлекательной мишенью для разработки лекарств и широко изучается в Plasmodium как способ поиска лекарств для лечения малярии малярии ( «ахиллесова пята» ). Считается, что некоторые используемые в настоящее время противомалярийные препараты , такие как хлорохин и мефлохин , убивают малярийных паразитов путем ингибирования биокристаллизации гемозоина .
Открытие
[ редактировать ]Черно-коричневый пигмент наблюдал Иоганн Генрих Меккель. [ 1 ] в 1847 году в крови и селезенке человека, страдающего безумием. [ 2 ] Однако только в 1849 году наличие этого пигмента было связано с заражением малярией. [ 3 ] Первоначально считалось, что этот пигмент вырабатывается организмом в ответ на инфекцию, но в 1880 году Шарль Луи Альфонс Лаверан понял, что «пигмент малярии» вместо этого вырабатывается паразитами, когда они размножаются внутри эритроцитов . [ 4 ] Связь между пигментом и малярийными паразитами была использована Рональдом Россом для определения стадий жизненного цикла плазмодия , которые происходят внутри комара, поскольку, хотя эти формы паразита внешне отличаются от стадий крови, они все же содержат следы пигмента. . [ нужна ссылка ]
Позже, в 1891 г., Т. Карбон и У. Браун (1911) опубликовали работы, связывающие деградацию гемоглобина с образованием пигмента, описывая малярийный пигмент как форму гематина и опровергая широко распространенное представление о его связи с меланином . Браун заметил, что все меланины быстро отбеливаются перманганатом калия, в то время как с этим реагентом малярийный пигмент не проявляет ни малейшего признака истинной реакции отбеливания. [ 5 ] [ 6 ] Название «гемозоин» было предложено Луи Вестенра Самбоном . [ 7 ] В 1930-х годах несколько авторов идентифицировали гемозоин как чистую кристаллическую форму α-гематина и показали, что это вещество не содержит белков внутри кристаллов. [ 4 ] но не было дано никакого объяснения различий в растворимости малярийного пигмента и кристаллов α-гематина. [ нужна ссылка ]
Формирование
[ редактировать ]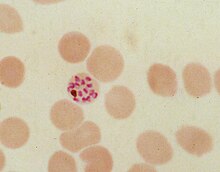
В ходе внутриэритроцитарного бесполого цикла размножения Plasmodium falciparum потребляет до 80% гемоглобина клетки-хозяина . [ 8 ] [ 9 ] При расщеплении гемоглобина высвобождается мономерный α-гематин ( феррипротопорфирин IX). Это соединение токсично, так как является прооксидантом и катализирует выработку активных форм кислорода . Считается, что окислительный стресс возникает во время превращения гема (ферропротопорфирина) в гематин (феррипротопорфирин). Свободный гематин также может связываться с клеточными мембранами и разрушать их , повреждая клеточные структуры и вызывая лизис эритроцитов хозяина. [ 10 ] Уникальная реакционная способность этой молекулы была продемонстрирована в нескольких in vitro и in vivo . экспериментальных условиях [ 11 ]

Таким образом, малярийный паразит обезвреживает гематин путем биокристаллизации β-гематина , превращая его в нерастворимые и химически инертные кристаллы (называемые гемозоином). [ 13 ] [ 14 ] [ 15 ] У Plasmodium пищевая вакуоль заполняется кристаллами гемозоина длиной около 100–200 нанометров , каждый из которых содержит около 80 000 молекул гема. [ 4 ] Детоксикация посредством биокристаллизации отличается от процесса детоксикации у млекопитающих, где фермент, называемый гемоксигеназой, вместо этого расщепляет избыток гема на биливердин , железо и окись углерода . [ 16 ]
Было предложено несколько механизмов производства гемозоина у плазмодий , и эта область весьма спорна: мембранные липиды , [ 17 ] [ 18 ] белки, богатые гистидином, [ 19 ] или даже комбинацию того и другого, [ 20 ] предлагается катализировать образование гемозоина. Другие авторы описали белок детоксикации гема, который, как утверждается, более эффективен, чем липиды или белки, богатые гистидином. [ 12 ] Возможно, что образованию гемозоина способствуют многие процессы. [ 21 ] Образование гемозоина у других кровососущих организмов изучено не так хорошо, как у Plasmodium . [ 22 ] Однако исследования Schistosoma mansoni показали, что этот паразитический червь производит большое количество гемозоина во время своего роста в кровотоке человека. Хотя форма кристаллов отличается от формы, вырабатываемой малярийными паразитами, [ 23 ] химический анализ пигмента показал, что он состоит из гемозоина. [ 24 ] [ 25 ] Аналогичным образом кристаллы, образующиеся в кишечнике целующегося клопа Rhodnius prolixus при переваривании кровяной муки, также имеют уникальную форму, но состоят из гемозоина. [ 26 ] Образование Hz в средней кишке R. prolixus происходит при физиологически значимых физико-химических условиях, а липиды играют важную роль в биокристаллизации гема. Выявлено, что автокаталитическая кристаллизация гема до Гц является неэффективным процессом, и эта конверсия еще больше снижается по мере увеличения концентрации Гц. [ 27 ]
Было разработано несколько других механизмов для защиты большого числа гематофагов от токсического воздействия свободного гема. Комары переваривают пищу, полученную из крови внеклеточно и не продуцируют гемозоин. Гем сохраняется в перитрофическом матриксе — слое белка и полисахаридов, который покрывает среднюю кишку и отделяет клетки кишечника от комка крови. [ 28 ]
Хотя β-гематин может спонтанно вырабатываться в анализах при низком pH , разработка простого и надежного метода измерения выработки гемозоина оказалась затруднительной. Частично это связано с продолжающейся неопределенностью относительно того, какие молекулы участвуют в производстве гемозоина, а частично из-за трудности измерения разницы между агрегированным или осажденным гемом и настоящим гемозоином. [ 29 ] Современные анализы чувствительны и точны, но требуют нескольких этапов промывки, поэтому они медленны и не идеальны для высокопроизводительного скрининга . [ 29 ] Тем не менее, с помощью этих анализов было проведено несколько скринингов. [ 30 ]
Структура
[ редактировать ]

Кристаллы β-гематина состоят из димеров молекул гематина, которые, в свою очередь, соединены водородными связями с образованием более крупных структур. В этих димерах железо - кислород координационная связь связывает центральное железо одного гематина с кислородом карбоксилатной боковой цепи соседнего гематина. Эти взаимные связи железо-кислород весьма необычны и не наблюдались ни в одном другом димере порфирина. β-гематин может представлять собой либо циклический димер , либо линейный полимер . [ 31 ] полимерная форма никогда не была обнаружена в гемозоине, что опровергает широко распространенное представление о том, что гемозоин вырабатывается ферментом гем -полимеразой. [ 32 ]
Кристаллы гемозоина имеют отчетливое триклинное строение и слабомагнитны . Разницу между диамагнитным низкоспиновым оксигемоглобином и парамагнитным гемозоином можно использовать для выделения. [ 33 ] [ 34 ] Они также демонстрируют оптический дихроизм , то есть сильнее поглощают свет по длине, чем по ширине, что позволяет автоматически обнаруживать малярию. [ 35 ] Гемозоин образуется в форме, которая под действием приложенного магнитного поля вызывает индуцированный оптический дихроизм , характерный для концентрации гемозоина; и точное измерение этого индуцированного дихроизма ( магнитного кругового дихроизма ) можно использовать для определения уровня малярийной инфекции. [ 36 ]
Ингибиторы
[ редактировать ]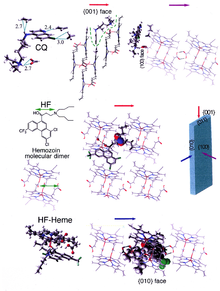
Образование гемозоина является отличной мишенью для лекарств, поскольку он необходим для выживания малярийных паразитов и отсутствует в организме человека. Гематин-мишень для лекарственного средства возникает у хозяина и в значительной степени находится вне генетического контроля паразита, что затрудняет развитие лекарственной устойчивости. Считается, что многие клинически используемые препараты действуют путем ингибирования образования гемозоина в пищевой вакуоли. [ 37 ] Это предотвращает детоксикацию гема, выделяющегося в этом отсеке, и убивает паразита. [ 38 ]
Наиболее изученными примерами таких ингибиторов биокристаллизации гематина являются хинолиновые препараты, такие как хлорохин и мефлохин . Эти препараты связываются как со свободным гемом, так и с кристаллами гемозоина. [ 39 ] и, следовательно, блокируют добавление новых единиц гема к растущим кристаллам. Считается, что маленькая, наиболее быстро растущая грань – это та грань, с которой связываются ингибиторы. [ 40 ] [ 41 ]
Роль в патофизиологии
[ редактировать ]Гемозоин высвобождается в кровоток во время реинфекции и фагоцитируется in vivo и in vitro фагоцитами хозяина и изменяет важные функции этих клеток. Большинство функциональных изменений были долгосрочными постфагоцитарными эффектами. [ 42 ] [ 43 ] включая ингибирование эритропоэза, показанное in vitro. [ 44 ] [ 45 ] [ 46 ] Напротив, было показано, что во время фагоцитоза nHZ происходит мощная кратковременная стимуляция окислительного взрыва моноцитами человека. [ 47 ] В иммунных клетках описано перекисное окисление липидов, неферментативно катализируемое гемозоиновым железом. [ 48 ] [ 49 ] Продукты липопероксидации, такие как гидроксиэйкозатетраеновые кислоты (HETE) и 4-гидроксиноненаль (4-HNE), функционально участвуют в иммуномодуляции. [ 43 ] [ 46 ] [ 49 ] [ 50 ] [ 51 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Янхуа Р.М., Шультка Р., Геббель Л., Пайт Т.Г., Шилдс CB (2010). «Наследие Иоганна Фридриха Меккеля Старшего (1724-1774): династия анатомов в четырех поколениях». Нейрохирургия . 66 (4): 758–770. дои : 10.1227/01.NEU.0000367997.45720.A6 . ПМИД 20305497 . S2CID 207143167 .
- ^ Меккель Х (1847 г.). «О черном пигменте в селезенке и крови психически больного человека». Журнал психиатрии . IV : 198–226.
- ^ Вирхов Р. (1849). «К патологической физиологии крови». Арх Патол Анатомия Физиол Клин Мед . 2 (3): 587–598. дои : 10.1007/BF02114475 . S2CID 40172296 .
- ^ Jump up to: а б с Салливан DJ (декабрь 2002 г.). «Теории образования малярийного пигмента и действия хинолина». Инт Дж Паразитол . 32 (13): 1645–53. дои : 10.1016/S0020-7519(02)00193-5 . ПМИД 12435449 .
- ^ Карбоне Т (1891). «О химической природе малярийного пигмента». GR Accad Med Турин . 39 : 901–906.
- ^ Браун WH (1911). «Малярийный пигмент (так называемый меланин): его природа и способ производства» . Джей Эксп Мед . 13 (2): 290–299. дои : 10.1084/jem.13.2.290 . ПМК 2124860 . ПМИД 19867409 .
- ^ Синтон Дж. А., Гош Б. Н. (1934). «Исследование малярийного пигмента (гемозоина). Часть I. Исследование действия растворителей на гемозоин и спектроскопических явлений, наблюдаемых в растворах». Отчеты обследования малярии в Индии . 4 : 15–42.
- ^ Розенталь П.Дж., Мешник С.Р. (декабрь 1996 г.). «Катаболизм гемоглобина и утилизация железа малярийными паразитами». Мол Биохим Паразитол . 83 (2): 131–139. дои : 10.1016/S0166-6851(96)02763-6 . ПМИД 9027746 .
- ^ Эспозито А., Тифферт Т., Мауриц Дж.М., Шлахтер С., Баннистер Л.Х., Камински К.Ф., Лью В.Л. (2008). Шнур Дж. М. (ред.). «FRET-визуализация концентрации гемоглобина в эритроцитах, инфицированных Plasmodium falciparum» . ПЛОС ОДИН . 3 (11): е3780. Бибкод : 2008PLoSO...3.3780E . дои : 10.1371/journal.pone.0003780 . ПМК 2582953 . ПМИД 19023444 .
- ^ Fitch CD, Chevli R, Kanjananggulpan P, Dutta P, Chevli K, Chou AC (1983). «Внутриклеточный феррипротопорфирин IX является литическим агентом» . Кровь . 62 (6): 1165–1168. дои : 10.1182/blood.V62.6.1165.1165 . ПМИД 6640106 .
- ^ Хеббель Р.П., Итон Дж.В. (апрель 1989 г.). «Патобиология взаимодействия гема с мембраной эритроцитов». Семин Гематол . 26 (2): 136–149. ПМИД 2658089 .
- ^ Jump up to: а б Яни Д., Нагаркатти Р., Битти В., Энджел Р., Слебодник С., Андерсен Дж., Кумар С., Ратор Д. (апрель 2008 г.). Ким К. (ред.). «HDP — новый белок для детоксикации гема от малярийного паразита» . ПЛОС Патог . 4 (4): e1000053. дои : 10.1371/journal.ppat.1000053 . ПМК 2291572 . ПМИД 18437218 .
- ^ Fitch CD, Kanjananggulpan P (ноябрь 1987 г.). «Состояние феррипротопорфирина IX в малярийном пигменте» . J Биол Хим . 262 (32): 15552–1555. дои : 10.1016/S0021-9258(18)47761-7 . ПМИД 3119578 .
- ^ Пагола С., Стивенс П.В., Боле Д.С., Косар А.Д., Мэдсен С.К. (март 2000 г.). «Структура малярийного пигмента бета-гематина». Природа . 404 (6775): 307–310. Бибкод : 2000Natur.404..307P . дои : 10.1038/35005132 . ПМИД 10749217 . S2CID 4420567 .
- ^ Хемпельманн Э (2007). «Биокристаллизация гемозоина у Plasmodium falciparum и противомалярийная активность ингибиторов кристаллизации» . Исследования паразитов . 100 (4): 671–676. дои : 10.1007/s00436-006-0313-x . ПМИД 17111179 . S2CID 30446678 . Архивировано из оригинала 10 июня 2011 г.
- ^ Кикучи Г., Ёсида Т., Ногучи М. (декабрь 2005 г.). «Гемоксигеназа и деградация гема». Биохимия Биофиз Рес Коммьюнити . 338 (1): 558–567. дои : 10.1016/j.bbrc.2005.08.020 . ПМИД 16115609 .
- ^ Пишотта Дж. М., Салливан Д. (июнь 2008 г.). «Гемозоин: масло против воды» . Паразитол Инт . 57 (2): 89–96. дои : 10.1016/j.parint.2007.09.009 . ПМК 2442017 . ПМИД 18373972 .
- ^ Хай НТ, Сима Й, Маэда А, Мен ТТ, Хираяма К, Хирасе А, Миядзава А, Камей А (2013). «Формирование гемозоина, опосредованное фосфолипидной мембраной: влияние физических свойств и свидетельства наличия мембраны, окружающей гемозоин» . ПЛОС ОДИН . 8 (7): e70025. Бибкод : 2013PLoSO...870025H . дои : 10.1371/journal.pone.0070025 . ПМЦ 3720957 . ПМИД 23894579 .
- ^ Салливан DJ, Глузман И.Ю., Голдберг Д.Е. (январь 1996 г.). «Образование гемозоина плазмодия, опосредованное белками, богатыми гистидином». Наука . 271 (5246): 219–222. Бибкод : 1996Sci...271..219S . дои : 10.1126/science.271.5246.219 . ПМИД 8539625 . S2CID 3489880 .
- ^ Пандей А.В., Баббарвал В.К., Окойе Дж.Н., Джоши Р.М., Пури С.К., Сингх Р.Л., Чаухан В.С. (сентябрь 2003 г.). «Образование гемозоина при малярии: двухэтапный процесс с участием богатых гистидином белков и липидов». Биохимия Биофиз Рес Коммьюнити . 308 (4): 736–743. дои : 10.1016/S0006-291X(03)01465-7 . ПМИД 12927780 .
- ^ Чуг М., Сундарараман В., Кумар С., Редди В.С., Сиддики В.А., Стюарт К.Д., Малхотра П. (апрель 2013 г.). «Белковый комплекс направляет образование гемоглобина в гемозоин у Plasmodium falciparum» . Proc Natl Acad Sci США . 110 (14): 5392–7. Бибкод : 2013PNAS..110.5392C . дои : 10.1073/pnas.1218412110 . ПМЦ 3619337 . ПМИД 23471987 .
- ^ Иган Т.Дж. (февраль 2008 г.). «Образование гемозоина». Мол Биохим Паразитол . 157 (2): 127–136. дои : 10.1016/j.molbiopara.2007.11.005 . ПМИД 18083247 .
- ^ Мур Г.А., Хомвуд Калифорния, Жиль Х.М. (сентябрь 1975 г.). «Сравнение пигмента Schistosoma mansoni и Plasmodium berghei». Энн Троп Мед Паразитол . 69 (3): 373–374. дои : 10.1080/00034983.1975.11687021 . ПМИД 1098591 .
- ^ Оливейра М.Ф., д'Авила Х.К., Торрес Ч.Р., Оливейра П.Л., Темпоне А.Дж., Румянек Ф.Д., Брага К.М., Силва Дж.Р., Данса-Петрецкий М., Оливейра М.А., де Соуза В., Феррейра С.Т. (ноябрь 2000 г.). «Гемозоин у Schistosoma mansoni». Мол Биохим Паразитол . 111 (1): 217–221. дои : 10.1016/S0166-6851(00)00299-1 . ПМИД 11087932 .
- ^ Корреа Соареш Ж.Б., Менезеш Д., Ваннье-Сантос М.А., Феррейра-Перейра А., Алмейда Г.Т., Венансио Т.М., Верёвски-Алмейда С., Зишири В.К., Кутер Д., Хантер Р., Иган Т.Дж., Оливейра М.Ф. (2009). Джонс МК (ред.). «Вмешательство в образование гемозоина представляет собой важный механизм шистосомицидного действия противомалярийных метаноидов» . PLOS Негль Троп Дис . 3 (7): е477. дои : 10.1371/journal.pntd.0000477 . ПМК 2703804 . ПМИД 19597543 .
- ^ Оливейра М.Ф., Кисия С.В., Гомес А., Косар А.Дж., Боле Д.С., Хемпельманн Е., Менезеш Д., Ваннье-Сантос М.А., Оливейра П.Л., Феррейра С.Т. (2005). «Структурная и морфологическая характеристика гемозоина, продуцируемого Schistosoma mansoni и Rhodnius prolixus» . ФЭБС Летт . 579 (27): 6010–6016. дои : 10.1016/j.febslet.2005.09.035 . ПМИД 16229843 .
- ^ Стиблер Р., Тимм Б.Л., Оливейра П.Л., Хирн Г.Р., Иган Т.Дж., Оливейра М.Ф. (2010). «О физико-химических и физиологических требованиях образования гемозоина, стимулируемого перимикровилларными мембранами в средней кишке Rhodnius prolixus». Насекомое Биохимия Мол Биол . 40 (3): 284–292. дои : 10.1016/j.ibmb.2009.12.013 . ПМИД 20060043 .
- ^ Паскоа В., Оливейра П.Л., Данса-Петрецкий М., Сильва Дж.Р., Альваренга П.Х., Джейкобс-Лорена М., Лемос Ф.Дж. (май 2002 г.). «Перитрофический матрикс Aedes aegypti и его взаимодействие с гемом во время пищеварения крови». Насекомое Биохимия Мол Биол . 32 (5): 517–523. дои : 10.1016/S0965-1748(01)00130-8 . ПМИД 11891128 .
- ^ Jump up to: а б Дорн А., Виппагунта С.Р., Матиле Х., Бубендорф А., Веннерстрем Дж.Л., Ридли Р.Г. (март 1998 г.). «Сравнение и анализ нескольких способов стимулирования полимеризации гематина (гема) и оценка ее инициирования in vitro». Биохим Фармакол . 55 (6): 737–747. дои : 10.1016/S0006-2952(97)00509-1 . ПМИД 9586945 .
- ^ Теквани Б.Л., Уокер Л.А. (февраль 2005 г.). «Нацеливание на путь синтеза гемозоина для открытия новых противомалярийных препаратов: технологии анализа образования бета-гематина in vitro». Экран Comb Chem с высокой пропускной способностью . 8 (1): 63–79. дои : 10.2174/1386207053328101 . ПМИД 15720198 .
- ^ Лемберг Р., Легге Дж.В. (1949). «Соединения гематина и желчные пигменты» . Интерсайенс, Нью-Йорк .
- ^ Хемпельманн Э., Маркес Х.М. (сентябрь 1994 г.). «Анализ малярийного пигмента Plasmodium falciparum». J Фармаколические токсиколовые методы . 32 (1): 25–30. дои : 10.1016/1056-8719(94)90013-2 . ПМИД 7833503 .
- ^ Пол Ф., Роат С., Мелвилл Д., Уорхерст, округ Колумбия, Осисанья Д.О. (1981). «Выделение инфицированных малярией эритроцитов из цельной крови: использование метода селективной высокоградиентной магнитной сепарации». Ланцет . 2 (8237): 70–71. дои : 10.1016/S0140-6736(81)90414-1 . ПМИД 6113443 . S2CID 42076 .
- ^ Ким CC, Уилсон Э.Б., Дериси Дж.Л. (2010). «Улучшенные методы магнитной очистки малярийных паразитов и гемозоина» (PDF) . Малар Дж . 9 (1): 17. дои : 10.1186/1475-2875-9-17 . ПМЦ 2817699 . ПМИД 20074366 .
- ^ Менделоу Б.В., Лайонс С., Нланготи П., Тана М., Мюнстер М., Випкема Э., Либовиц Л., Маршалл Л., Скотт С., Кутцер Т.Л. (1999). «Автоматическое обнаружение малярии путем деполяризации лазерного света» . Br J Гематол . 104 (3): 499–503. дои : 10.1046/j.1365-2141.1999.01199.x . ПМИД 10086786 . S2CID 26054638 . [ мертвая ссылка ]
- ^ Ньюман Д.М., Хептинстал Дж., Мателон Р.Дж., Сэвидж Л., Уэрс М.Л., Беддоу Дж., Кокс М., Шаллиг HD, Менс П (2008). «Магнитооптический путь к диагностике малярии in vivo: предварительные результаты и данные доклинических испытаний» (PDF) . Биофиз Дж . 95 (2): 994–1000. Бибкод : 2008BpJ....95..994N . дои : 10.1529/biophysj.107.128140 . ПМК 2440472 . ПМИД 18390603 . Архивировано из оригинала (PDF) 27 июля 2011 г. Проверено 19 декабря 2009 г.
- ^ Зиглер Дж., Линк Р., Райт Д.В. (февраль 2001 г.). «Ингибиторы агрегации гема: противомалярийные препараты, воздействующие на важный процесс биоминерализации». Curr Med Chem . 8 (2): 171–89. дои : 10.2174/0929867013373840 . ПМИД 11172673 .
- ^ Коронадо Л.М., Надович К.Т., Спадафора К. (2014). «Малярийный гемозоин: от мишени к инструменту» . Биохим Биофиз Акта . 1840 (6): 2032–2041. дои : 10.1016/j.bbagen.2014.02.009 . ПМК 4049529 . ПМИД 24556123 .
- ^ Салливан DJ, Глузман И.Ю., Рассел Д.Г., Голдберг Д.Е. (октябрь 1996 г.). «О молекулярном механизме противомалярийного действия хлорохина» . Proc Natl Acad Sci США . 93 (21): 11865–70. Бибкод : 1996PNAS...9311865S . дои : 10.1073/pnas.93.21.11865 . ПМК 38150 . ПМИД 8876229 .
- ^ де Вильерс К.А., Маркес Х.М., Иган Т.Дж. (август 2008 г.). «Кристаллическая структура галофантрин-феррипротопорфирина IX и механизм действия арилметаноловых противомалярийных препаратов». J Inorg Biochem . 102 (8): 1660–1667. дои : 10.1016/j.jinorgbio.2008.04.001 . ПМИД 18508124 .
- ^ Вайсбух I, Лейзеровиц Л (2008). «Взаимодействие между малярией, образованием кристаллического гемозоина, действием и дизайном противомалярийных препаратов». Хим. преп . 108 (11): 4899–4914. дои : 10.1021/cr078274t . ПМИД 19006402 .
- ^ Арез П., Шварцер Э (1997). «Малярийный пигмент (гемозоин): очень активное «инертное» вещество». Энн Троп Мед Паразитол . 91 (5): 501–516. дои : 10.1080/00034989760879 . ПМИД 9329987 .
- ^ Jump up to: а б Скороход О.А., Алессио М., Мордмюллер Б., Арезе П., Шварцер Э. (2004). «Гемозоин (малярийный пигмент) ингибирует дифференцировку и созревание дендритных клеток, происходящих из моноцитов человека: гамма-опосредованный эффект, активируемый пролифератором пероксисомы» . Дж Иммунол . 173 (6): 4066–74. дои : 10.4049/jimmunol.173.6.4066 . ПМИД 15356156 .
- ^ Гирибальди Г., Уллиерс Д., Шварцер Э., Робертс И., Пьячибелло В., Арезе П. (2004). «Гемозоин- и 4-гидроксиноненаль-опосредованное ингибирование эритропоэза. Возможная роль в малярийном дизэритропоэзе и анемии». Гематологическая . 89 (4): 492–493. ПМИД 15075084 .
- ^ Казальс-Паскуаль С, Кай О, Чунг Дж.О., Уильямс С., Лоу Б., Ньяноти М., Уильямс Т.Н., Мейтленд К., Молинье М., Ньютон С.Р., Пешу Н., Ватт С.М., Робертс DJ (2006). «Подавление эритропоэза при малярийной анемии связано с гемозоином in vitro и in vivo» . Кровь . 108 (8): 2569–77. дои : 10.1182/blood-2006-05-018697 . ПМИД 16804108 .
- ^ Jump up to: а б Скороход О.А., Кайоне Л., Маррокко Т., Мильярди Г., Баррера В., Арезе П., Пьячибелло В., Шварцер Э. (2010). «Ингибирование эритропоэза при малярийной анемии: роль гемозоина и генерируемого гемозоином 4-гидроксиноненаля» . Кровь . 116 (20): 4328–37. doi : 10.1182/blood-2010-03-272781 . ПМИД 20686121 .
- ^ Баррера В., Скороход О.А., Бачи Д., Гремо Г., Арезе П., Шварцер Э. (2011). «Фибриноген хозяина, стабильно связанный с гемозоином, быстро активирует моноциты посредством TLR-4 и CD11b/CD18-интегрина: новая парадигма действия гемозоина» . Кровь . 117 (21): 5674–82. дои : 10.1182/кровь-2010-10-312413 . ПМИД 21460246 .
- ^ Шварцер Э., Мюллер О., Арезе П., Симс В.Г., Грюн Т. (1996). «Повышенные уровни 4-гидроксиноненаля в моноцитах человека, питающихся малярийным пигментом гемозоином. Возможный ключ к разгадке токсичности гемозоина» . ФЭБС Летт . 388 (2–3): 119–22. дои : 10.1016/0014-5793(96)00523-6 . ПМИД 8690068 . S2CID 23594376 .
- ^ Jump up to: а б Шварцер Э., Арезе П., Скороход О.А. (2015). «Роль продукта липопероксидации 4-гидроксиноненала в патогенезе тяжелой малярийной анемии и иммунодепрессии против малярии» . Оксид Мед Селл Лонгев . 2015 : 638416. doi : 10.1155/2015/638416 . ПМК 4417603 . ПМИД 25969702 .
- ^ Скороход О, Баррера В, Мандили Г, Костанца Ф, Валенте Э, Уллиерс Д, Шварцер Э (2021). «Малярийный пигмент гемозоин ухудшает экспрессию и функцию рецептора GM-CSF посредством 4-гидроксиноненала» . Антиоксиданты . 10 (8): 1259. doi : 10.3390/antiox10081259 . ПМЦ 8389202 . ПМИД 34439507 .
- ^ Скороход О.А., Баррера В., Хеллер Р., Карта Ф., Туррини Ф., Арез П., Шварцер Э. (2014). «Малярийный пигмент гемозоин нарушает хемотаксическую подвижность и трансэндотелиальную миграцию моноцитов посредством 4-гидроксиноненаля» . Свободный Радик Биол Мед . 75 : 210–221. doi : 10.1016/j.freeradbiomed.2014.07.004 . hdl : 2318/148326 . ПМИД 25017964 .