Метастабильность в мозге
В области вычислительной нейробиологии теория метастабильности относится к способности человеческого мозга интегрировать несколько функциональных частей и производить нейронные колебания совместным и скоординированным образом, обеспечивая основу для сознательной деятельности.
Метастабильность, состояние, в котором сигналы (например, колебательные волны) выходят за пределы своего естественного состояния равновесия, но сохраняются в течение длительного периода времени, — это принцип, описывающий способность мозга находить смысл в кажущихся случайными сигналах окружающей среды. За последние 25 лет интерес к метастабильности и основам нелинейной динамики подогревался достижениями в методах, с помощью которых компьютеры моделируют активность мозга.
Обзор
[ редактировать ]ЭЭГ измеряет общую электрическую активность мозга, которую можно наблюдать на поверхности черепа. В теории метастабильности выходные данные ЭЭГ производят колебания, которые можно описать как имеющие идентифицируемые паттерны, коррелирующие друг с другом на определенных частотах . Каждый нейрон в нейронной сети обычно выдает динамический колебательный сигнал, но также имеет возможность выдавать хаотичный сигнал. [1] Когда нейроны интегрируются в нейронную сеть путем взаимодействия нейронов друг с другом, динамические колебания, создаваемые каждым нейроном, могут быть объединены для формирования высоко предсказуемых колебаний ЭЭГ.
Выявляя эти корреляции и отдельные нейроны, которые способствуют предсказуемым колебаниям ЭЭГ, ученые могут определить, какие корковые домены обрабатывают данные параллельно и какие нейронные сети переплетаются. Во многих случаях метастабильность описывает случаи, когда дистальные части мозга взаимодействуют друг с другом, реагируя на раздражители окружающей среды.
Частотные области метастабильности
[ редактировать ]Было высказано предположение, что одним из неотъемлемых аспектов динамики мозга, лежащих в основе сознательного мышления, является способность мозга преобразовывать кажущиеся шумными или хаотичные сигналы в предсказуемые колебательные паттерны. [2]
В ЭЭГ-колебаниях нейронных сетей частоты соседних сигналов коррелируют в логарифмическом, а не в линейном масштабе. В результате средние частоты в колебательных полосах не могут связываться друг с другом согласно линейности их средних частот. Вместо этого фазовые переходы связаны в соответствии с их способностью сочетаться с соседними фазовыми сдвигами в постоянном состоянии перехода между нестабильной и стабильной фазовой синхронизацией . [2] Эта фазовая синхронизация лежит в основе метастабильного поведения нейронных сетей.
Метастабильное поведение происходит в высокочастотной области, известной как режим 1/f . Этот режим описывает среду, в которой был индуцирован шумный сигнал (также известный как розовый шум ), где количество мощности , выдаваемой сигналом в определенной полосе пропускания (его спектральная плотность мощности ), обратно пропорционально его частоте.
Шум в режиме 1/f можно обнаружить во многих биологических системах – например, в выходных сигналах сердцебиения в форме волны ЭКГ – но он служит уникальной цели для фазовой синхронности в нейронных сетях. В режиме 1/f мозг находится в критическом состоянии, необходимом для сознательной реакции на слабые или хаотичные сигналы окружающей среды, поскольку он может превращать случайные сигналы в идентифицируемые и предсказуемые колебательные формы. [2] Хотя эти сигналы часто преходящи, они существуют в стабильной форме достаточно долго, чтобы способствовать тому, что можно рассматривать как сознательную реакцию на стимулы окружающей среды.
Теории метастабильности
[ редактировать ]Колебательная активность и координационная динамика
[ редактировать ]Модель динамической системы , которая представляет собой сети, состоящие из интегрированных нейронных систем, взаимодействующих друг с другом между нестабильными и стабильными фазами, становится все более популярной теорией, лежащей в основе понимания метастабильности. [3] Координационная динамика [4] формирует основу для этой модели динамической системы, описывая математические формулы и парадигмы, управляющие связью стимулов окружающей среды с их эффекторами.
История динамики координации и модель Хакена-Келсо-Бунца (HKB)
[ редактировать ]Так называемая модель HKB — одна из самых ранних и уважаемых теорий, описывающих динамику координации в мозге. В этой модели формирование нейронных сетей можно частично описать как самоорганизацию , при которой отдельные нейроны и небольшие нейрональные системы объединяются и координируются, чтобы либо адаптироваться или реагировать на локальные стимулы, либо разделить труд и специализироваться на функциях. [5]
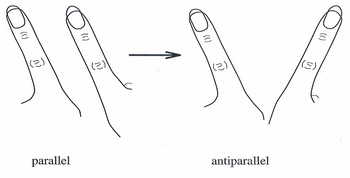
За последние 20 лет, [ когда? ] Модель HKB стала широко распространенной теорией, объясняющей скоординированные движения и поведение отдельных нейронов в больших сквозных нейронных сетях. Первоначально модель описывала систему, в которой спонтанные переходы, наблюдаемые в движениях пальцев, можно было описать как серию синфазных и противофазных движений. [6]
В экспериментах на модели HKB в середине 1980-х годов испытуемых просили помахать одним пальцем на каждой руке в двух направлениях: первый, известный как противофазный, когда оба пальца двигались в одном и том же направлении вперед и назад (как могут двигаться дворники). ; и второй, известный как синфазный, когда оба пальца соединяются и движутся к средней линии тела и от нее. Чтобы проиллюстрировать динамику координации, испытуемых просили двигать пальцами в противофазе с возрастающей скоростью до тех пор, пока их пальцы не стали двигаться как можно быстрее. Когда движение приближалось к критической скорости, пальцы испытуемых переходили от противофазного движения (как у дворников) к синфазному (движению по средней линии).
Модель HKB, которая также была объяснена несколькими сложными математическими дескрипторами, по-прежнему остается относительно простым, но мощным способом описания, казалось бы, независимых систем, которые достигают синхронности непосредственно перед состоянием самоорганизованной критичности . [6] [7]
Эволюция динамики когнитивной координации
[ редактировать ]За последние 10 лет, [ когда? ] Модель HKB была согласована с передовыми математическими моделями и вычислениями на базе суперкомпьютеров, чтобы связать элементарную динамику координации с процессами более высокого порядка, такими как обучение и память.
Традиционная ЭЭГ по-прежнему полезна для изучения координации между различными частями мозга. частотой 40 Гц Активность гамма-волн является ярким примером способности мозга к динамическому моделированию и распространенным примером динамики координации. Постоянное изучение этих и других колебаний привело к важному выводу: анализ волн как имеющих общую фазу сигнала, но разную амплитуду, приводит к возможности того, что эти разные сигналы выполняют синергетическую функцию. [8]
Некоторые необычные характеристики этих волн: они практически одновременны и имеют очень короткую задержку начала, что означает, что они действуют быстрее, чем синаптическая проводимость позволяет ; и что их узнаваемые закономерности иногда прерываются периодами случайности . Последняя особенность послужила основанием для предположения о взаимодействии и переходе между нервными подсистемами. Анализ активации и деактивации областей коры показал динамический сдвиг между зависимостью и взаимозависимостью природу мозга , отражающий метастабильную как функции скоординированной динамической системы.
фМРТ , крупномасштабные электродные матрицы и МЭГ расширяют закономерности, наблюдаемые на ЭЭГ, обеспечивая визуальное подтверждение скоординированной динамики. МЭГ, которая обеспечивает улучшение пространственно-временных характеристик по сравнению с ЭЭГ, позволяет исследователям стимулировать определенные части мозга сигналами окружающей среды и наблюдать реакцию в целостной модели мозга . Кроме того, время отклика MEG составляет около одной миллисекунды , что позволяет практически в реальном времени исследовать активное включение и выключение выбранных частей мозга в ответ на сигналы окружающей среды и сознательные задачи. [9]
Динамика социальной координации и фи-комплекс
[ редактировать ]Развивающаяся область динамики координации включает в себя теорию социальной координации, которая пытается связать DC с нормальным человеческим развитием сложных социальных сигналов, следующих определенным моделям взаимодействия. Целью данной работы является понимание того, как социальное взаимодействие человека опосредовано метастабильностью нейронных сетей. фМРТ и ЭЭГ особенно полезны для картирования таламокортикальной реакции на социальные сигналы в экспериментальных исследованиях.
Новая теория под названием «фи-комплекс» была разработана Дж. А. Скоттом Келсо и его коллегами из Атлантического университета Флориды с целью предоставить экспериментальные результаты для теории динамики социальной координации. [10] В экспериментах Келсо двух испытуемых разделяли непрозрачным барьером и просили помахивать пальцами; затем барьер убрали, и испытуемых попросили продолжать вилять пальцами, как будто никаких изменений не произошло. Через непродолжительный период движения двух испытуемых иногда становились скоординированными и синхронизированными (но в других случаях продолжали быть асинхронными). Связь между ЭЭГ и сознательным социальным взаимодействием описывается как Фи — один из нескольких ритмов мозга, работающих в диапазоне 10 Гц. Фи состоит из двух компонентов: один способствует одиночному поведению, а другой — интерактивному ( межличностному ) поведению. Дальнейший анализ Фи может выявить социальные и межличностные последствия дегенеративных заболеваний , таких как шизофрения , или может дать представление об общих социальных отношениях, таких как динамика альфа- и омега-самцов или популярный эффект свидетеля, описывающий, как люди распределяют личную ответственность в чрезвычайных ситуациях. в зависимости от количества присутствующих других людей.
Динамическое ядро
[ редактировать ]Вторая теория метастабильности включает в себя так называемое динамическое ядро — термин, обозначающий таламокортикальную область, которая считается интеграционным центром сознания . Гипотеза динамического ядра (DCH) отражает использование и неиспользование взаимосвязанных нейронных сетей во время стимуляции этой области. Компьютерная модель 65 000 импульсных нейронов. [8] показывает, что группы нейронов, существующие в коре и таламусе, взаимодействуют в форме синхронных колебаний. Взаимодействие между отдельными группами нейронов формирует динамическое ядро и может помочь объяснить природу сознательного опыта. Важнейшей особенностью DCH является то, что вместо бинарного подхода к переходам между нейронной интеграцией и неинтеграцией (т. е. что эти два явления являются либо одним, либо другим, без промежуточного состояния), метастабильная природа динамического ядра может допускать континуум интеграции. [8]
Нейронный дарвинизм
[ редактировать ]Одна теория, используемая для интеграции динамического ядра с сознательным мышлением, включает в себя развивающуюся концепцию, известную как нейронный дарвинизм . [11] В этой модели метастабильные взаимодействия в таламокортикальной области вызывают процесс селекции посредством повторного входа (феномен, описывающий общую взаимность и интерактивность между сигналами в удаленных частях мозга посредством связанной задержки сигнала). Нейрональная селективность включает в себя механохимические события, которые происходят пре- и постнатально, в результате чего на нейрональные связи влияют условия окружающей среды. [12] Модификация синаптических сигналов в отношении динамического ядра дает дальнейшее объяснение DCH.
Несмотря на растущее количество доказательств существования DCH, способность генерировать математические конструкции для моделирования и прогнозирования динамического поведения ядра развивается медленно. [13] Продолжающееся развитие стохастических процессов, предназначенных для графического отображения нейронных сигналов как хаотических и нелинейных, обеспечило некоторую алгоритмическую основу для анализа того, как хаотические сигналы окружающей среды соединяются для повышения избирательности нейронного роста или координации в динамическом ядре.
Гипотеза глобального рабочего пространства
[ редактировать ]Гипотеза глобального рабочего пространства — это еще одна теория, объясняющая метастабильность, которая в той или иной форме существует с 1983 года. [14] Эта гипотеза также фокусируется на феномене повторного входа, способности рутины или процесса одновременно использоваться несколькими частями мозга. [8] И модель DCH, и модель глобального нейронного рабочего пространства (GNW) предполагают повторный вход, но модель GNW развивает возвратную связь между удаленными частями мозга и потоком сигналов на большие расстояния. Нейроны рабочей области схожи анатомически, но пространственно отделены друг от друга.
Одним из интересных аспектов GNW является то, что при достаточной интенсивности и длине сигнала небольшой инициирующий сигнал может быть составлен для активации «зажигания» критического состояния, вызывающего всплеск. Эта идея аналогична лыжнику на склоне горы, который, разбивая лыжами несколько глыб льда, вызывает гигантскую лавину за собой . Чтобы доказать теорию схемоподобного усиления, исследования показали, что повреждение соединений на больших расстояниях снижает производительность интегративных моделей. [8]
Популярный эксперимент, демонстрирующий гипотезу глобального рабочего пространства, включает в себя показ испытуемому серии визуальных слов, замаскированных назад (например, «собака тихо спит» отображается как «илтейук произносит бог эт» ), а затем просят испытуемого определить, что написано вперед. перевод» этих слов. ФМРТ не только обнаружила активность в части коры головного мозга, отвечающей за распознавание слов, но, кроме того, активность часто обнаруживается в теменной и префронтальной коре . [15] Почти в каждом эксперименте сознательный ввод слов и задач на аудирование показывает гораздо более широкое использование интегрированных частей мозга, чем при идентичном бессознательном вводе. Широкое распространение и постоянная передача сигналов между различными областями мозга в экспериментальных результатах является распространенным методом попытки доказать гипотезу нейронного рабочего пространства. Проводятся дополнительные исследования, чтобы точно определить корреляцию между сознательным и бессознательным обдумыванием задач в сфере глобального рабочего пространства.
Теория оперативной архитектоники мозга и разума
[ редактировать ]Хотя концепция метастабильности уже некоторое время существует в нейробиологии, [16] Конкретную интерпретацию метастабильности в контексте мозговых операций различной сложности разработали Эндрю и Александр Фингелькурты в рамках своей модели Операционной архитектоники функционирования мозга и разума . Метастабильность — это, по сути, теория о том, как глобальные интегративные и локальные сегрегационные тенденции сосуществуют в мозге. [17] [18] В центре внимания Операционной архитектоники лежит тот факт, что в метастабильном режиме функционирования мозга отдельные части мозга проявляют тенденции к функционированию автономно и в то же время проявляют тенденции к скоординированной деятельности. [19] [20] В соответствии с Операционной Архитектоникой, [21] синхронизированные операции, производимые распределенными нейронными группами, составляют метастабильные пространственно-временные паттерны. Они метастабильны, поскольку внутренние различия в активности между нейронными группами достаточно велики, чтобы каждый из них выполнял свою собственную работу (операцию), сохраняя при этом тенденцию координироваться вместе для реализации сложной операции мозга. [22] [23]
Будущее метастабильности
[ редактировать ]В дополнение к изучению влияния метастабильных взаимодействий на традиционные социальные функции, многие исследования, вероятно, будут сосредоточены на определении роли скоординированной динамической системы и глобального рабочего пространства в прогрессировании изнурительных заболеваний, таких как болезнь Альцгеймера , болезнь Паркинсона , инсульт и шизофрения. [24]
Интерес к влиянию черепно-мозговой травмы ( ЧМТ ) на координированную динамическую систему возрос в последние пять лет, поскольку число случаев ЧМТ возросло из-за травм, связанных с войной.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Тиран, П; М. Хаслер (18 декабря 1994 г.). «Обработка информации с использованием устойчивых и неустойчивых колебаний: Учебное пособие». Труды третьего международного семинара IEEE по сотовым нейронным сетям и их приложениям (CNNA-94) . стр. 127–136. дои : 10.1109/cnna.1994.381695 . ISBN 978-0-7803-2070-3 . S2CID 61939952 .
- ^ Перейти обратно: а б с Бужаки, Дьёрдь (2006). Ритмы мозга . США: Издательство Оксфордского университета. стр. 128–31 . ISBN 978-0-19-530106-9 .
- ^ Фингелькуртс, А.; А. Фингелькурц (2004). «Упрощая сложность: многовариантность и метастабильность в мозге». Международный журнал неврологии . 114 (7): 843–862. дои : 10.1080/00207450490450046 . ПМИД 15204050 . S2CID 11376883 .
- ^ А.С. Келсо, Джеймс (июль 2014 г.). Координационная динамика. В Энциклопедии сложности и системных наук . Спрингер. стр. 1–17. ISBN 978-3-642-27737-5 .
- ^ Кольер, Т.; Чарльз Тейлор (июль 2004 г.). «Самоорганизация в сенсорных сетях» . J. Параллельные и распределенные вычисления . 64 (7): 866–873. дои : 10.1016/j.jpdc.2003.12.004 . Архивировано из оригинала (PDF) 16 мая 2008 г. Проверено 26 ноября 2007 г.
- ^ Перейти обратно: а б Фукс, А.; В.К. Йирса (2000). «Возвращение к модели HKB: как изменение степени симметрии контролирует динамику». Наука о движении человека . 19 (4): 425–449. дои : 10.1016/S0167-9457(00)00025-7 .
- ^ Келсо, Дж. А. Скотт; и др. (1988). «Генерация динамических паттернов в поведенческих и нейронных системах». Наука . 239 (4847): 1513–1520. Бибкод : 1988Sci...239.1513S . дои : 10.1126/science.3281253 . ПМИД 3281253 .
- ^ Перейти обратно: а б с д и Вернер, АГ; ВК Йирса (сентябрь 2007 г.). «Метастабильность, критичность и фазовые переходы в мозге и его моделях» (PDF) . Биосистемы . 90 (2): 496–508. Бибкод : 2007BiSys..90..496W . doi : 10.1016/j.biosystems.2006.12.001 . ПМИД 17316974 .
- ^ Йирса, ВК; А. Фукс; JAS Келсо (ноябрь 1998 г.). «Соединение корковой и поведенческой динамики: бимануальная координация». Нейронные вычисления . 10 (8): 2019–2045. дои : 10.1162/089976698300016954 . ПМИД 9804670 . S2CID 13605465 .
- ^ Тоньоли, Э; и др. (март 2007 г.). «Фи-комплекс как нейромаркер социальной координации человека» . ПНАС . 104 (19): 8190–8195. дои : 10.1073/pnas.0611453104 . ПМК 1859993 . ПМИД 17470821 .
- ^ Сет, А.; Б. Баарс (2005). «Нейронный дарвинизм и сознание». Сознание и познание . 14 (1): 140–168. дои : 10.1016/j.concog.2004.08.008 . ПМИД 15766895 . S2CID 6262356 .
- ^ Эдельман, Джеральд (1987). Нейронный дарвинизм: теория отбора групп нейронов . Нью-Йорк, Нью-Йорк: Основные книги. ISBN 978-0-19-286089-7 .
- ^ Ле Ван Куен, М. (2003). «Распутывание динамического ядра: исследовательская программа нейродинамики в крупном масштабе» . Биол. Рез . 36 (1): 67–88. дои : 10.4067/s0716-97602003000100006 . ПМИД 12795207 .
- ^ Баарс, Бернард (октябрь 2003 г.). «Обновление теории глобального рабочего пространства» . Обзор науки и сознания . Проверено 26 ноября 2007 г.
- ^ ДеХаен, С.; Л. Наккаш (2001). «На пути к когнитивной нейробиологии сознания: основные данные и рамки рабочего пространства». Познание . 79 (1): 1–37. дои : 10.1016/S0010-0277(00)00123-2 . ПМИД 11164022 . S2CID 1762431 .
- ^ Дж. А. Скотт Келсо (1991) Генерация поведенческих и нейронных паттернов: концепция нейроповеденческой динамической системы (NBDS). В: Koepchen HP (ред.) Кардиореспираторная и двигательная координация. Springer, Берлин, Гейдельберг, Нью-Йорк.
- ^ Бресслер С.Л., Келсо Дж.А. (2001). «Динамика корковой координации и познание». Тенденции Cogn Sci . 5 (1): 26–36. дои : 10.1016/s1364-6613(00)01564-3 . ПМИД 11164733 . S2CID 6010953 .
- ^ Каплан А.Я. (1998) Нестационарная ЭЭГ: методологический и экспериментальный анализ. Усп Физиол Наук (Успехи в физиологических науках) 29:35–55 (на русском языке).
- ^ Фингелькурц АнА Фингелькурц АлА (2001). «Оперативная архитектоника биопотенциального поля мозга человека: на пути к решению проблемы разум-мозг». Мозг и разум . 2 (3): 261–296. дои : 10.1023/А:1014427822738 . S2CID 16125062 .
- ^ Фингелькурц АнА Фингелькурц АлА (2004). «Упрощая сложность: многовариантность и метастабильность в мозге». Int J Neurosci . 114 (7): 843–862. дои : 10.1080/00207450490450046 . ПМИД 15204050 . S2CID 11376883 .
- ^ «Оперативная архитектоника» (PDF) . Архивировано из оригинала (PDF) 27 сентября 2007 г. Проверено 2 декабря 2007 г.
- ^ Фингелькурц АнА, Фингелькурц АлА (2005)Картирование оперативной архитектоники мозга. Глава 2. В: Chen FJ (ред.) Фокус на исследованиях картирования мозга. Nova Science Publishers, Inc. , стр. 59–98. URL = http://www.bm-science.com/team/chapt3.pdf . Архивировано 27 сентября 2007 г. на Wayback Machine.
- ^ Фингелькурц АнА Фингелькурц АнА (2006). «Время в познании и динамике мозга ЭЭГ: дискретность против непрерывности». Процесс познания . 7 (3): 135–162. дои : 10.1007/s10339-006-0035-0 . ПМИД 16832687 . S2CID 2228893 .
- ^ «Лаборатория человеческого мозга и поведения» . Центр сложных систем и наук о мозге — Атлантический университет Флориды . Архивировано из оригинала 23 сентября 2007 г. Проверено 26 ноября 2007 г.