Иммунопреципитация метилированной ДНК
Иммунопреципитация метилированной ДНК (MeDIP или mDIP) — это крупномасштабный (по всей хромосоме или всему геному ) метод очистки в молекулярной биологии, который используется для обогащения метилированных последовательностей ДНК . Он заключается в выделении метилированных фрагментов ДНК с помощью антитела, выработанного против 5-метилцитозина (5mC). Этот метод был впервые описан Weber M. et al. [ 1 ] в 2005 году и помог проложить путь для жизнеспособных усилий по оценке уровня метилома , поскольку очищенная фракция метилированной ДНК может быть использована в высокопроизводительных методах обнаружения ДНК, таких как микрочипы ДНК высокого разрешения ( MeDIP-чип ) или секвенирование следующего поколения. (MeDIP-seq). Тем не менее, понимание метилома остается зачаточным; его изучение осложняется тем фактом, что, как и другие эпигенетические свойства, закономерности варьируются от типа клеток к типу клеток.
Фон
[ редактировать ]Метилирование ДНК , относящееся к обратимому метилированию 5-го положения цитозина метилтрансферазами , является основной эпигенетической модификацией в многоклеточных организмах. [ 2 ] У млекопитающих эта модификация в основном происходит в сайтах CpG , которые, в свою очередь, имеют тенденцию группироваться в регионах, называемых CpG-островками . [ 3 ] Существует небольшая часть CpG-островков, которые могут перекрываться или находиться в непосредственной близости от промоторных областей сайтов начала транскрипции. Модификация также может произойти на других сайтах, [ 4 ] но метилирование в любом из этих сайтов может подавлять экспрессию генов, либо вмешиваясь в связывание факторов транскрипции , либо модифицируя структуру хроматина до репрессивного состояния. [ 5 ]
Исследования заболеваний во многом способствовали усилиям по пониманию роли метилирования ДНК. В настоящее время основной исследовательский интерес заключается в изучении таких заболеваний, как рак, с целью выявления областей ДНК, которые претерпели значительные изменения метилирования. Гены, содержащиеся в этих регионах, представляют функциональный интерес, поскольку могут предложить механистическое объяснение основных генетических причин заболеваний. Например, аномальный характер метилирования раковых клеток. [ 6 ] [ 7 ] [ 8 ] Первоначально было показано, что это механизм, посредством которого опухолевым супрессорам , подавляются гены, подобные [ 9 ] хотя позже было замечено, что затрагивается гораздо более широкий диапазон типов генов. [ 10 ] [ 11 ] [ 12 ]
Другие технологии
[ редактировать ]Существует два подхода к анализу метилирования: технологии типирования и профилирования. Технологии типирования нацелены на небольшое количество локусов во многих образцах и включают использование таких методов, как ПЦР , ферменты рестрикции и масс-спектрометрия . Технологии профилирования, такие как MeDIP, нацелены на генома или метилома оценку метилирования на уровне всего ; это включает геномное сканирование ориентиров рестрикции (RLGS), [ 13 ] и методы, основанные на преобразовании бисульфита , которые основаны на обработке ДНК бисульфитом для преобразования неметилированных цитозина остатков в урацил . [ 14 ] [ 15 ] [ 16 ] [ 17 ]
Ограничения других технологий
[ редактировать ]Другие методы картирования и профилирования метилома оказались эффективными, но не лишены ограничений, которые могут повлиять на разрешение, уровень производительности или экспериментальные вариации. Например, RLGS ограничен количеством сайтов рестрикции в геноме, которые могут быть мишенями для фермента рестрикции; обычно можно оценить максимум ~4100 ориентиров. [ 18 ] Методы, основанные на бисульфитном секвенировании , несмотря на возможное разрешение отдельных нуклеотидов, имеют недостаток: превращение неметилированного цитозина в урацил может быть нестабильным. [ 19 ] Кроме того, когда преобразование бисульфита сочетается с микрочипами ДНК для обнаружения сайтов, преобразованных в бисульфит, проблемой становится снижение сложности последовательности ДНК. Микрочипы, способные всесторонне профилировать весь геном, становятся трудными для разработки, поскольку доступно меньше уникальных зондов. [ 20 ]
Методы
[ редактировать ]В следующих разделах описывается метод MeDIP в сочетании либо с гибридизацией массива высокого разрешения, либо с высокопроизводительным секвенированием. Каждый метод обнаружения ДНК также кратко описывает постлабораторную обработку и анализ. В зависимости от технологии, используемой для идентификации метилированных последовательностей, требуются различные постобработки необработанных данных. Это аналогично данным, генерируемым с помощью ChIP-chip и ChIP-seq .
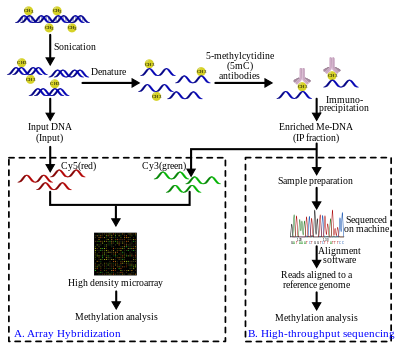
Иммунопреципитация метилированной ДНК (MeDIP)
[ редактировать ]Геномная ДНК извлекается ( экстракция ДНК ) из клеток и очищается. Затем очищенную ДНК подвергают обработке ультразвуком, чтобы разбить ее на случайные фрагменты. Этот процесс обработки ультразвуком является быстрым, простым и позволяет избежать ферментов рестрикции предвзятости . Полученные фрагменты имеют длину от 300 до 1000 пар оснований (п.н.), хотя обычно они составляют от 400 до 600 п.н. [ 21 ] Короткая длина этих фрагментов важна для получения адекватного разрешения, повышения эффективности последующего этапа иммунопреципитации и уменьшения эффектов или систематических ошибок, связанных с длиной фрагмента. Кроме того, размер фрагмента влияет на связывание антитела 5-метилцитидина (5mC), поскольку для эффективного связывания антителу требуется больше, чем просто один 5mC. [ 22 ] Для дальнейшего улучшения аффинности связывания антител фрагменты ДНК денатурируют с образованием одноцепочечной ДНК. После денатурации ДНК инкубируют с моноклональными антителами 5mC. классический метод иммунопреципитации Затем применяется магнитные шарики, конъюгированные с IgG : для связывания антител против 5mC используются мыши, а несвязанная ДНК удаляется в супернатанте. Для очистки ДНК добавляется протеиназа К , которая расщепляет антитела и высвобождает ДНК, которую можно собрать и подготовить для обнаружения ДНК.
Более подробную информацию об экспериментальных этапах см. [ 1 ] [ 19 ] [ 23 ] [ 24 ]
MeDIP и гибридизация на основе массива (MeDIP-чип)
[ редактировать ]Часть входной ДНК, полученная после описанного выше этапа обработки ультразвуком, помечена цианином -5 (Cy5; красный) дезоксицитозинтрифосфатом, тогда как метилированная ДНК, обогащенная после этапа иммунопреципитации, помечена цианином -3 (Cy3; зеленый). . Меченые образцы ДНК когибридизируются на двухканальном геномном микрочипе высокой плотности для проверки присутствия и относительных количеств. Целью этого сравнения является идентификация последовательностей, которые демонстрируют значительные различия в уровнях гибридизации, тем самым подтверждая, что интересующая последовательность обогащена. Идентификация последовательностей MeDIP на основе массива ограничена конструкцией массива. В результате разрешение ограничивается датчиками конструкции массива. При обработке сигналов требуются дополнительные стандартные шаги для устранения проблем гибридизации, таких как шум, как это имеет место в большинстве технологий массивов.
Видеть [ 23 ] [ 24 ] [ 25 ] для более подробной информации.
MeDIP и высокопроизводительное секвенирование (MeDIP-seq)
[ редактировать ]Подход MeDIP-seq, то есть сочетание MeDIP с технологиями секвенирования короткого считывания следующего поколения, такими как пиросеквенирование 454 или Illumina (Solexa), был впервые описан Down et al. в 2008 году. [ 20 ] Высокопроизводительное секвенирование метилированных фрагментов ДНК дает большое количество коротких прочтений (36-50 п.н.). [ 26 ] или 400 п.н., [ 27 ] в зависимости от технологии). Короткие чтения выравниваются по эталонному геному с помощью программного обеспечения для выравнивания, такого как Mapping and Assembly with Quality ( Maq ), которое использует байесовский подход, а также базовые и картографические качества для моделирования вероятностей ошибок для выравнивания. [ 28 ] Затем чтения можно расширить, чтобы представить фрагменты размером от ~ 400 до 700 п.н., полученные на этапе обработки ультразвуком. Охват этих расширенных чтений можно использовать для оценки уровня метилирования региона. геномный браузер , такой как Ensembl Для визуализации данных также можно использовать .
Валидацию подхода к оценке качества и точности данных можно провести с помощью количественной ПЦР . Это делается путем сравнения последовательности образца MeDIP с неметилированной контрольной последовательностью. Затем образцы помещают на гель и сравнивают интенсивности полос. [ 19 ] Относительная интенсивность служит ориентиром для определения обогащения. Результаты также можно сравнить с результатами чипа MeDIP, чтобы определить необходимое покрытие.
Последующий биоинформатический анализ
[ редактировать ]
Оценки уровня метилирования ДНК могут быть искажены из-за различной плотности метилированных сайтов CpG по всему геному при наблюдении данных, генерируемых MeDIP. Это может быть проблематично для анализа регионов с низким содержанием CpG (низкой плотности). Одной из причин этой проблемы плотности является ее влияние на эффективность иммунопреципитации. В своем исследовании Down et al. [ 20 ] разработали инструмент для оценки абсолютных уровней метилирования на основе данных, полученных с помощью MeDIP, путем моделирования плотности метилированных сайтов CpG. Этот инструмент называется байесовским инструментом для анализа метилирования (Бэтмен) . В исследовании сообщается о покрытии ~ 90% всех сайтов CpG в промоторах, областях кодирования генов, островках и регуляторных элементах, где можно оценить уровни метилирования; это почти в 20 раз лучшее покрытие, чем любые предыдущие методы.
Исследования с использованием MeDIP-seq или MeDIP-чипа представляют собой полногеномные подходы, имеющие общую цель - получить функциональное картирование метилома. Как только будут идентифицированы области метилирования ДНК, для ответа на определенные биологические вопросы можно будет применить ряд биоинформатических анализов. Одним из очевидных шагов является исследование генов, содержащихся в этих регионах, и изучение функционального значения их репрессии. Например, подавление генов-супрессоров опухолей при раке можно объяснить метилированием ДНК. [ 29 ] Выявив мутационные события, приводящие к гиперметилированию и последующей репрессии известных генов-супрессоров опухолей, можно более конкретно охарактеризовать факторы, способствующие причине заболевания. Альтернативно, можно идентифицировать гены, которые, как известно, обычно метилированы, но в результате какого-либо события мутации больше не подавляются.
Кроме того, можно попытаться исследовать и определить, был ли затронут какой-либо эпигенетический регулятор, такой как ДНК-метилтрансфераза (DNMT); [ 21 ] в этих случаях обогащение может быть более ограниченным.
Было показано, что анализ генного набора (например, с использованием таких инструментов, как DAVID и GoSeq) дает серьезные искажения при применении к данным высокопроизводительного метилирования (например, MeDIP-seq и MeDIP-ChIP); Было высказано предположение, что это можно исправить с помощью перестановок меток образцов или использования статистической модели для контроля различий в количестве зондов CpG/сайтов CpG, нацеленных на каждый ген. [ 30 ]
Ограничения MeDIP
[ редактировать ]Ограничения, которые следует учитывать при использовании MeDIP, являются типичными экспериментальными факторами. Сюда входит качество и перекрестная реактивность антител 5mC, используемых в процедуре. Более того, методы обнаружения ДНК (т.е. матричная гибридизация и высокопроизводительное секвенирование) обычно имеют четко установленные ограничения. Как упоминалось выше, в частности, для процедур на основе массива анализируемые последовательности ограничены конкретной используемой конструкцией массива.
Применяются наиболее типичные ограничения высокопроизводительного секвенирования следующего поколения. Проблема точности выравнивания повторяющихся участков генома приведет к менее точному анализу метилирования в этих участках. Кроме того, как упоминалось выше, короткие чтения (например, 36-50 пар оснований по данным анализатора генома Illumina ) представляют собой часть срезанного фрагмента при выравнивании с геномом; следовательно, точный сайт метилирования может находиться в любом месте окна, которое зависит от размера фрагмента. [ 19 ] В этом отношении бисульфитное секвенирование имеет гораздо более высокое разрешение (вплоть до одного сайта CpG; уровень одного нуклеотида). Однако этот уровень разрешения может не потребоваться для большинства приложений, поскольку было показано, что статус метилирования сайтов CpG в пределах <1000 п.н. значительно коррелирует. [ 20 ]
Применение MeDIP
[ редактировать ]- Вебер и др. 2005 г. [ 1 ] определили, что неактивная Х-хромосома у женщин гиперметилирована на уровне всей хромосомы с использованием MeDIP в сочетании с микрочипом.
- Кешет и др. 2006 г. [ 31 ] провели исследование на клетках рака толстой кишки и простаты с использованием MeDIP-чипа. Результатом стал полногеномный анализ генов, лежащих в гиперметилированных областях, а также вывод о том, что существует поучительный механизм метилирования de novo в раковых клетках.
- Чжан и др. 2006 г. [ 24 ] получили картирование метилома высокого разрешения у Arabidopsis с использованием MeDIP-чипа.
- Новак и др. 2006 г. [ 32 ] использовали подход MeDIP-чипа для исследования рака молочной железы человека на предмет молчания, связанного с метилированием, и наблюдали инактивацию кластера генов HOXA.
См. также
[ редактировать ]- Эпигенетика
- Иммунопреципитация
- Метилом
- Геномное сканирование рестрикционных ориентиров
- Бисульфитное секвенирование
Ссылки
[ редактировать ]- ^ Jump up to: а б с Вебер М., Дэвис Дж. Дж., Виттиг Д. и др. (август 2005 г.). «Общехромосомный и специфичный для промотора анализ выявляет участки дифференциального метилирования ДНК в нормальных и трансформированных клетках человека». Нат. Жене . 37 (8): 853–62. дои : 10.1038/ng1598 . ПМИД 16007088 . S2CID 14505320 .
- ^ Птица А (январь 2002 г.). «Схемы метилирования ДНК и эпигенетическая память» . Генс Дев . 16 (1): 6–21. дои : 10.1101/gad.947102 . ПМИД 11782440 .
- ^ Гардинер-Гарден М., Фроммер М. (июль 1987 г.). «CpG-островки в геномах позвоночных». Дж. Мол. Биол . 196 (2): 261–82. дои : 10.1016/0022-2836(87)90689-9 . ПМИД 3656447 .
- ^ Кларк С.Дж., Харрисон Дж., Фроммер М. (май 1995 г.). «Метилирование CpNpG в клетках млекопитающих». Нат. Жене . 10 (1): 20–7. дои : 10.1038/ng0595-20 . ПМИД 7647784 . S2CID 8857760 .
- ^ Джениш Р., Бёрд А (март 2003 г.). «Эпигенетическая регуляция экспрессии генов: как геном объединяет внутренние сигналы и сигналы окружающей среды». Нат. Жене . 33 (Приложение): 245–54. дои : 10.1038/ng1089 . ПМИД 12610534 . S2CID 17270515 .
- ^ Робертсон К.Д., Вольф А.П. (октябрь 2000 г.). «Метилирование ДНК в здоровье и болезни». Нат. Преподобный Жене . 1 (1): 11–9. дои : 10.1038/35049533 . ПМИД 11262868 . S2CID 1915808 .
- ^ Байлин С.Б. , Герман Дж.Г. (2000). «Гиперметилирование ДНК при онкогенезе: эпигенетика присоединяется к генетике» . Тенденции Жене . 16 (4): 268–274. дои : 10.1016/S0168-9525(99)01971-X . ПМИД 10729832 .
- ^ Джонс, Пенсильвания, Лэрд П.В. (февраль 1999 г.). «Эпигенетика рака достигает совершеннолетия». Нат. Жене . 21 (2): 163–7. дои : 10.1038/5947 . ПМИД 9988266 . S2CID 10328397 .
- ^ Джонс, Пенсильвания, Бэйлин С.Б. (июнь 2002 г.). «Фундаментальная роль эпигенетических событий при раке». Нат. Преподобный Жене . 3 (6): 415–28. дои : 10.1038/nrg816 . ПМИД 12042769 . S2CID 2122000 .
- ^ Костелло Дж.Ф., Фрювальд М.К., Смираглиа Д.Дж. и др. (февраль 2000 г.). «Аберрантное метилирование CpG-островков имеет неслучайные и специфичные для типа опухоли закономерности». Нат. Жене . 24 (2): 132–8. дои : 10.1038/72785 . ПМИД 10655057 . S2CID 6273676 .
- ^ Зардо Г., Тиирикайнен М.И., Хонг С. и др. (ноябрь 2002 г.). «Комплексный геномный и эпигеномный анализ выявляет инактивацию биаллельных генов в опухолях». Нат. Жене . 32 (3): 453–8. дои : 10.1038/ng1007 . ПМИД 12355068 . S2CID 23645606 .
- ^ Ю Л., Лю С., Вандейсен Дж. и др. (март 2005 г.). «Глобальная оценка метилирования промотора в мышиной модели рака идентифицирует ID4 как предполагаемый ген-супрессор опухолей при лейкемии человека». Нат. Жене . 37 (3): 265–74. дои : 10.1038/ng1521 . ПМИД 15723065 . S2CID 515916 .
- ^ Хатада И., Хаясидзаки Ю., Хироцунэ С., Комацубара Х., Мукаи Т. (ноябрь 1991 г.). «Метод геномного сканирования высших организмов с использованием сайтов рестрикции в качестве ориентиров» . Учеб. Натл. акад. наук. США . 88 (21): 9523–7. Бибкод : 1991PNAS...88.9523H . дои : 10.1073/pnas.88.21.9523 . ПМК 52750 . ПМИД 1946366 .
- ^ Ракян В.К., Хильдманн Т., Новик К.Л. и др. (декабрь 2004 г.). «Профиль метилирования ДНК главного комплекса гистосовместимости человека: пилотное исследование для проекта эпигенома человека» . ПЛОС Биол . 2 (12): е405. дои : 10.1371/journal.pbio.0020405 . ПМК 529316 . ПМИД 15550986 .

- ^ Гитан Р.С., Ши Х., Чен С.М., Ян П.С., Хуан Т.Х. (январь 2002 г.). «Микроматрица олигонуклеотидов, специфичных для метилирования: новый потенциал для высокопроизводительного анализа метилирования» . Геном Рез . 12 (1): 158–64. дои : 10.1101/гр.202801 . ПМК 155260 . ПМИД 11779841 .
- ^ Мейснер А., Гнирке А., Белл Г.В., Рамсахой Б., Ландер Э.С., Джениш Р. (2005). «Бисульфитное секвенирование с уменьшенным представлением для сравнительного анализа метилирования ДНК с высоким разрешением» . Нуклеиновые кислоты Рез . 33 (18): 5868–77. дои : 10.1093/nar/gki901 . ПМК 1258174 . ПМИД 16224102 .
- ^ Адорьян П., Дистлер Дж., Липшер Э. и др. (март 2002 г.). «Прогнозирование и обнаружение классов опухолей с помощью анализа метилирования ДНК на основе микрочипов» . Нуклеиновые кислоты Рез . 30 (5): 21д–21. дои : 10.1093/nar/30.5.e21 . ПМК 101257 . ПМИД 11861926 .
- ^ Дай З., Вейченхан Д., Ву Ю.З. и др. (октябрь 2002 г.). «Пограничная библиотека AscI для изучения генетических и эпигенетических изменений на островах CpG» . Геном Рез . 12 (10): 1591–8. дои : 10.1101/гр.197402 . ПМК 187524 . ПМИД 12368252 .
- ^ Jump up to: а б с д Помранинг К.Р., Смит К.М., Фрайтаг М. (март 2009 г.). «Полногеномный высокопроизводительный анализ метилирования ДНК у эукариот». Методы . 47 (3): 142–50. дои : 10.1016/j.ymeth.2008.09.022 . ПМИД 18950712 .
- ^ Jump up to: а б с д Даун Т.А., Ракян В.К., Тернер Д.Д. и др. (июль 2008 г.). «Стратегия байесовской деконволюции для анализа метиломов ДНК на основе иммунопреципитации» . Нат. Биотехнология . 26 (7): 779–85. дои : 10.1038/nbt1414 . ПМК 2644410 . ПМИД 18612301 .
- ^ Jump up to: а б Хасинто Ф.В., Баллестар Э, Эстеллер М (январь 2008 г.). «Иммунопреципитация метил-ДНК (MeDIP): поиск метилома ДНК» . БиоТехники . 44 (1): 35–43. дои : 10.2144/000112708 . ПМИД 18254377 .
- ^ Михан Р.Р., Льюис Дж.Д., Берд А.П. (октябрь 1992 г.). «Характеристика MeCP2, ДНК-связывающего белка позвоночных, обладающего сродством к метилированной ДНК» . Нуклеиновые кислоты Рез . 20 (19): 5085–92. дои : 10.1093/нар/20.19.5085 . ПМК 334288 . ПМИД 1408825 .
- ^ Jump up to: а б Уилсон И.М. и др. (2005). «Эпигеномика: картирование метилома» . Клеточный цикл . 5 (2): 155–8. дои : 10.4161/cc.5.2.2367 . ПМИД 16397413 .
- ^ Jump up to: а б с Чжан X, Язаки Дж, Сундаресан А и др. (сентябрь 2006 г.). «Полногеномное картирование с высоким разрешением и функциональный анализ метилирования ДНК арабидопсиса» . Клетка . 126 (6): 1189–201. дои : 10.1016/j.cell.2006.08.003 . ПМИД 16949657 .
- ^ Микрочип метилирования ДНК
- ^ «Illumina | Секвенирование и решения на основе массивов для генетических исследований» .
- ^ «Секвенирование, скрининг и анализ ДНК: приборы и услуги — 454 Life Sciences™» . Архивировано из оригинала 18 марта 2008 г. Проверено 18 марта 2008 г.
- ^ Ли Х, Руан Дж, Дурбин Р (ноябрь 2008 г.). «Картирование коротких прочтений секвенирования ДНК и вызов вариантов с использованием показателей качества картирования» . Геном Рез . 18 (11): 1851–8. дои : 10.1101/гр.078212.108 . ПМК 2577856 . ПМИД 18714091 .
- ^ Эстеллер М. (апрель 2007 г.). «Эпигенетическое молчание генов при раке: гиперметилом ДНК» . Хм. Мол. Жене . 16 (Спецификация № 1): 50–9 рэндов. дои : 10.1093/hmg/ddm018 . ПМИД 17613547 .
- ^ Гилихер П., Хартнетт Л., Иган Л.Дж., Голден А., Раджа Али Р.А., Сои С. (июнь 2013 г.). «Анализ набора генов сильно необъективен при применении к данным полногеномного метилирования» . Биоинформатика . 29 (15): 1851–7. doi : 10.1093/биоинформатика/btt311 . ПМИД 23732277 .
- ^ Кешет И., Шлезингер Ю., Фаркаш С. и др. (февраль 2006 г.). «Доказательства поучительного механизма метилирования de novo в раковых клетках». Нат. Жене . 38 (2): 149–53. дои : 10.1038/ng1719 . ПМИД 16444255 . S2CID 14405389 .
- ^ Новак П., Дженсен Т., Оширо М.М. и др. (ноябрь 2006 г.). «Эпигенетическая инактивация кластера генов HOXA при раке молочной железы» . Рак Рез . 66 (22): 10664–70. дои : 10.1158/0008-5472.CAN-06-2761 . ПМИД 17090521 .