СПМАП1
| СПМАП1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | SPMAP1 , открытая рамка считывания 98 хромосомы 17, белок 1, связанный с микротрубочками спермия | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | МГИ : 1919465 ; Гомологен : 19140 ; Генные карты : SPMAP1 ; ОМА : SPMAP1 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Белок 1, ассоциированный с микротрубочками спермы, представляет собой белок, который у человека кодируется геном SPMAP1 . Белок происходит из 17-й хромосомы человека разумного . [ 5 ] Ген C17orf98 состоит из последовательности из 6302 оснований. Его мРНК имеет три экзона и не имеет альтернативных сайтов сплайсинга. Белок содержит 154 аминокислоты, аномальных уровней аминокислот нет. [ 6 ] C17orf98 имеет домен неизвестной функции (DUF4542) и весит 17,6 кДа. [ 7 ] [ 8 ] C17orf98 не принадлежит ни к каким другим семействам и не имеет изоформ. [ 9 ] Белок имеет ортологи с высоким процентом сходства у млекопитающих и рептилий. У белка есть дополнительные отдаленные ортологи в царстве многоклеточных животных , кульминацией которого является семейство губок. [ 10 ]
Известно, что, как и большинство белков, C17orf98 высоко экспрессируется в семенниках. [ 11 ] Также известно, что белок имеет повышенный уровень при раке. [ 11 ] Было показано, что белок экспрессируется вблизи или внутри промежуточных филаментов и ядрышка . [ 11 ] Кроме того, c17orf98 содержит факторы транскрипции, которые также активны в гемопоэтических стволовых клетках , иммунной и сердечно-сосудистой системах , среди других. [ 12 ] Этот ген сверхэкспрессируется при многих типах рака, включая светлоклеточный рак почек и плоскоклеточный рак легких. [ 13 ] Анализ мотивов и факторов транскрипции указывает на то, что c17orf98 играет роль в пролиферации, особенно в пролиферации иммунных клеток.
Ген
[ редактировать ]Фон
[ редактировать ]Ген C17orf98 состоит из 6303 оснований. Он имеет три экзона и два больших интрона. Ген не имеет альтернативных сайтов сплайсинга. [ 14 ] Последовательность 5'- UTR C17orf98 высоко консервативна у приматов. Никаких совпадений 5' UTR, не относящихся к млекопитающим, определить не удалось. [ 15 ] [ 16 ] C17orf98 имеет 11 повторов Alu . [ 17 ]
Усилители
[ редактировать ]GeneCards определила, что C17orf98 имеет пять энхансерных последовательностей. Роль этих последовательностей может дать представление о функции C17orf98. Четыре из пяти энхансеров активны в тимусе. Все пять энхансеров активны в ЭСК H1. Кроме того, все пять энхансеров активны в iPS DF 19.11, полученном из фибробластов крайней плоти. [ 18 ]
Транскрипционные факторы
[ редактировать ]Промотор C17orf98 имеет множество сайтов связывания транскрипционных факторов. [ 19 ] Факторы транскрипции C17orf98 обычно обнаруживаются в гемопоэтических клетках, соединительной ткани, сердечно-сосудистой ткани и иммунной системе. Присутствие круппелеподобных факторов транскрипции предполагает роль c17orf 98 в пролиферации или апоптозе. Присутствие SMAD указывает на участие в пути TGF-β, тогда как присутствие факторов транскрипции, связанных с Myc, указывает на потенциальную пролиферационную функцию белка. Кроме того, другие факторы транскрипции C17orf98, такие как RBPJ-Kappa, участвуют в пролиферации и передаче сигналов.
Варианты
[ редактировать ]Многочисленные SNP были обнаружены в 5'-UTR, 3'-UTR и кодирующей области c17orf98. [ 20 ] Немногие SNP были обнаружены в высококонсервативных регионах. Всего в составе высококонсервативных аминокислот было обнаружено четыре SNP. Один SNP был обнаружен в последовательности стартового кодона. Из этих пяти трое имели SNP в третьей позиции кодона. Согласно гипотезе колебания , три из пяти SNP не окажут влияния на общую структуру белка.
мРНК
[ редактировать ]C17orf98 не имеет сайтов связывания микроРНК. [ 21 ] Его мРНК имеет низкую распространенность (0,44%). [ 22 ] Последовательность мРНК состоит из трех гексапетлей, ни одна из которых не является значимой. [ 23 ]
Белок
[ редактировать ]Первичная структура
[ редактировать ]C17orf98 представляет собой белок массой 17,6 кДа. [ 8 ] Отдаленные ортологи на 5–6 кДа больше, но некоторые расхождения происходят из-за добавленной последовательности NLS, которой нет у Homo sapiens . Кластеров положительного или отрицательного заряда нет. Трансмембранные компоненты отсутствуют. Изоэлектрическая точка составляет 9,80/17564,67 пи/МВт. [ 24 ] C17orf98 гидрофобен и растворим.

Вторичная и третичная структура
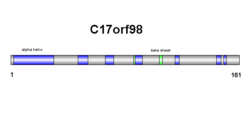
[ редактировать ]Вторичная структура c17orf98 состоит как из бета-листов, так и из альфа-спиралей (см. диаграмму справа). Результаты подтверждаются в третичной структуре, однако номера альфа-спирали и бета-листов немного различаются (см. диаграмму справа).
Мотивы и сайты связывания
[ редактировать ]N-концевые сигнальные пептиды отсутствуют. Мотивы спайности не обнаружены. Нет сигналов удерживания мембраны ЭР и сигнала пероксисомального нацеливания. SKL2 отсутствует, следовательно, сигнал вторичной пероксисомы отсутствует. Сигналы нацеливания на вакуоли отсутствуют. Не существует РНК-связывающих мотивов или актин-связывающих мотивов актининового типа. Паттерн N-миристоилирования или паттерн пренилирования отсутствует. [ 25 ]

Специалист по поиску киназ в Cuckoo определил сайты связывания киназы для c17orf98. Существует множество сайтов фосфорилирования серина/треонина и тирозинкиназы. [ 26 ] Сайты связывания серин- и треонин-киназы являются наиболее распространенными и превышают статистически значимый порог. Сайтов СУМОйляции нет. [ 27 ] Ген C17orf98 имеет шесть сайтов в последовательности возможных сайтов O-GlcNAc . [ 28 ] Высококонсервативными аминокислотными участками O-GlcNAc являются 24, 32, 117 и 142. Посттрансляционная модификация O-GlcNAc происходит на остатках Ser/Thr, особенно на онкогенах, опухолевых супрессорах и белках, участвующих в передаче сигналов факторов роста. [ 29 ]
C17orf98 имеет мотив Caspase3/7, при котором расщепляется либо каспаза 3, либо 7. [ 30 ] Это подтверждает идею о том, что C17orf98 участвует в пролиферации, поскольку проапоптотическая каспаза желает уничтожить любой белок, стимулирующий пролиферацию. Белок также имеет мотив, с которым связывается пептидил-пролил-цис-транс-изомераза NIMA 1 (Pin1). [ 30 ] Повышение регуляции Pin1 участвует в развитии рака и иммунных нарушений. [ 31 ] Это подтверждает утверждение о том, что C17orf98 участвует в развитии рака, иммунных клеток и, возможно, рака иммунной системы. Кроме того, белок C17orf98 имеет сайт IBM, с которым связываются ингибиторы апоптоза (IAP). [ 30 ] Это еще раз подтверждает идею о том, что C17orf98 участвует в ингибировании апоптоза и, логически, вызывает рак. Более того, C17orf98 имеет мотивы, с которыми GRB2 связывается домен SH2 . GRB2 представляет собой адаптерный белок, участвующий в сигнальном пути RAS, пути, дерегуляция которого приводит к неконтролируемой пролиферации.
Аминокислотная последовательность
[ редактировать ]Дублирование могло произойти в позициях 59–71.
Мудрый человек
MAYLSECRLRLEKGFILDGVAVSTAARAYGRSRPKLWSAIPPYNAQQDYHARSYFQ SHVVPPLLRVVPPLLRKTDQDHGGTGRDGWIVDYIHIFGQGQRYLNRRNWAGTGHS LQQVTGHDHYNADLKPIDGFNGRFGYRRNTPALRQSTSVFGEVTHFPLF
Связанные белки
[ редактировать ]Нет известных ассоциированных белков. [ 32 ] [ 33 ] [ 34 ] [ 35 ]
Выражение
[ редактировать ]Содержание белка в целом организме Homo sapiens довольно низкое. По другим видам данных нет. [ 36 ] Атлас мозга Аллена не дает атласа мозга для c17orf98. [ 37 ]
Субклеточная локализация
[ редактировать ]Было обнаружено, что белок C17orf98 экспрессируется в промежуточных нитях и ядрышках . [ 38 ] Антитело C17orf98 доступно от Sigma-Aldrich. [ 39 ] Кроме того, C17orf98 локализуется в цитоплазме. Дальнеродственные ортологи c17orf98 у таких организмов, как Macrostomum lignano и Amphimedon queenslandica, демонстрируют ядерную экспрессию. [ 40 ] Сигналы ядерной локализации присутствуют у отдаленно родственных организмов в неконсервативных участках. Результатом предсказания k-NN является цитоплазматическая локализация. [ 41 ] C17orf98 не является сигнальным пептидом. [ 42 ] Белок является растворимым. [ 43 ]
Салфетка
[ редактировать ]Как и большинство белков, белок C17orf98 высоко экспрессируется в семенниках. [ 44 ] Белок экспрессируется как в тканях взрослых, так и в тканях плода. Было обнаружено, что белок слабо экспрессируется в соединительной ткани. [ 45 ] Кроме того, экспрессия наблюдалась в сперме, эпителиальных клетках молочной железы и различных клетках иммунной системы. [ 46 ]
Клиническое значение
[ редактировать ]Рак
[ редактировать ]Экспрессия белка повышена у многих онкологических больных. В частности, было показано, что экспрессия белка высока в колоректальном кишечнике, молочной железе, простате и легких. [ 47 ] C17orf98 также экспрессируется при папиллярном раке щитовидной железы. [ 48 ] Кроме того, мутации в c17orf98 были обнаружены при раке эндометрия, желудка, колоратуры и почки. [ 49 ] Экспрессия C17orf98 повышена у онкологических больных с BRCA. У пациентов со светлоклеточной карциномой почки экспрессия c17orf98 резко снижается по сравнению с нераковым состоянием. [ 13 ] У 80% пациентов с хромофобной почечно-клеточной карциномой присутствовала по крайней мере одна дупликация гена c17orf98. [ 13 ]
Другие условия
[ редактировать ]Экспрессия белка ниже у мужчин с тератозооспермией по сравнению с мужчинами без нее. [ 50 ] С C17orf98 было проведено множество экспериментов по географическому профилю, однако ни один из них не дал данных, показывающих значительные изменения в экспрессии. [ 51 ]
Эволюция
[ редактировать ]C17orf98 — медленно мутирующий белок. Он напоминает цитохром c по скорости расхождения, определяемой уравнениями молекулярных часов. [ 52 ]

Паралоги
[ редактировать ]не известно . Homo sapiens Паралогов C17orf98 [ 53 ]
Ортологи
[ редактировать ]Белок C17orf98 имеет дополнительные отдаленные ортологи в царстве многоклеточных животных. Его самый дальний родственник принадлежит к семейству губок. У гребневиков, нематод, бактерий, грибов, растений и рыбок данио не существует известного ортолога. [ 10 ] Есть только две рыбы с геном C17orf 98. Модельные организмы, такие как Caenorhabditis elegans и Drosophila melanogaster , не имеют этого гена.
C17orf98 Ортологи [ 10 ]
| Последовательность # | Род и вид | Общее имя | Номер доступа | Длина белка | MYA Div | Идентификатор последовательности | Уверенность |
|---|---|---|---|---|---|---|---|
| 1 | Мудрый человек | Человек | НП_001073934 | 154 | 0 | 100% | уже |
| 2 | Дикий верблюд | Дикий двугорбый верблюд | XP_006176436 | 154 | 96 | 83% | 2.00Э-94 |
| 3 | Птеропус был выбран | Черная летучая лисица | XP_006924784 | 154 | 96 | 81% | 1.00Э-92 |
| 4 | Липоты вексилифер | Дельфин реки Янцзы | XP_007465208 | 154 | 96 | 81% | 6.00Э-89 |
| 5 | Кондилура кристат | Звездоносый крот | XP_004684322 | 154 | 96 | 75% | 5.00Э-78 |
| 6 | Миотис бренди | бита Брандта | EPQ05064 | 171 | 96 | 78% | 6.00Э-78 |
| 7 | Ворчать ворчать ворчать | Альпийский сурок | XP_015362150.1 | 154 | 90 | 81% | 3.00Э-94 |
| 8 | Октодон дегу | Чилийский грызун | XP_004633931 | 153 | 90 | 73% | 1.00Э-76 |
| 9 | Аллигатор китайский | Китайский аллигатор | XP_006022630 | 154 | 312 | 63% | 8.00Э-68 |
| 10 | Анолис Каролинский | Ящерица | XP_003222553 | 154 | 312 | 62% | 6.00Э-67 |
| 11 | Ксенопус левис | Африканская когтистая лягушка | XP_018090228 | 244 | 352 | 51% | 4.00Э-38 |
| 12 | Тип ринкодона | Китовая акула | XP_020388051.1 | 164 | 476 | 53% | 5.00Э-52 |
| 13 | Акантастер плачи | морская звезда | XP_022086463 | 209 | 684 | 48% | 1.00Э-37 |
| 14 | Мизухопектен дасоенсис | Гребешок | XP_021340301 | 275 | 797 | 45% | 5.00E-06 |
| 15 | Лоттия гигантская | Морская улитка | XP_009063876 | 173 | 797 | 45% | 2.00Э-37 |
| 16 | Утята | Корпус лампы | XP_013388744.1 | 211 | 797 | 43% | 2.00Э-35 |
| 17 | Биомфалярия гладкая | Пресноводная улитка | XP_013088317 | 198 | 797 | 41% | 6.00E-15 |
| 18 | Нематостелла вектенсис | Морской анемон | XP_001629616 | 173 | 824 | 48% | 2.00Э-35 |
| 19 | Стилофора пестилистая | Коралл | XP_022795125 | 226 | 824 | 46% | 3.00Э-38 |
| 20 | Макростонум лигнано | Плоский червь | ПАА73615 | 235 | 824 | 36% | 4.00E-25 |
| 21 | Амфимедона королевская | Губка | XP_003389909 | 275 | 951.8 | 32% | 2.00E-12 |
Ссылки
[ редактировать ]- ^ Jump up to: а б с ENSG00000276913 GRCh38: выпуск Ensembl 89: ENSG00000275489, ENSG00000276913 — Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000018543 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Зоди М.К., Гарбер М., Адамс Д.Д., Шарп Т., Харроу Дж., Лупски Дж.Р. и др. (апрель 2006 г.). «Последовательность ДНК 17-й хромосомы человека и анализ перестроек в линии человека» . Природа . 440 (7087): 1045–9. Бибкод : 2006Natur.440.1045Z . дои : 10.1038/nature04689 . ПМК 2610434 . ПМИД 16625196 .
- ^ Запись PSORT II на c17orf98 https://psort.hgc.jp/form2.html
- ^ Запись о консервативных доменах NCBI C17orf98
- ^ Jump up to: а б Запись ENMBL-EBI SAPS на c17orf98
- ^ «BLAST: базовый инструмент поиска локального выравнивания» . blast.ncbi.nlm.nih.gov . Проверено 2 мая 2018 г.
- ^ Jump up to: а б с «Protein BLAST: поиск по базам данных белков с помощью запроса белков» . blast.ncbi.nlm.nih.gov . Проверено 2 мая 2018 г.
- ^ Jump up to: а б с Запись в атласе белков человека по c17orf98
- ^ Genomatix El Derado etnry на c17orf98
- ^ Jump up to: а б с Запись TissGDB на c17orf98
- ^ Запись Acieview на c17orf98
- ^ Запись ClustalW на c17orf98 5 'UTR
- ^ Запись NCBI Blast на c17orf98 5' UTR https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blastho мне
- ^ Genomatix El Derado etnry на c17orf98 [ постоянная мертвая ссылка ]
- ^ База данных GeneCards Human Gene. «Ген C17orf98 - GeneCards - Белок CQ098 - Антитело CQ098» . www.genecards.org . Проверено 2 мая 2018 г.
- ^ «Genomatix El Derado etnry на c17orf98» . [ постоянная мертвая ссылка ]
- ^ Средство просмотра данных генома NCBI
- ^ Запись Target Scan на c17orf98 http://www.targetscan.org/cgibin/targetscan/vert_71/view_gene.cgi?rs=ENST00000398575.4&taxid=9606&showcnc=0&shownc=0&shownc_nc=&showncf1=&showncf2=&subset=1 [ постоянная мертвая ссылка ]
- ^ Запись Pax-db на c17orf98
- ^ «Запись mFold на c17orf98 5' UTR» . [ постоянная мертвая ссылка ]
- ^ Запись ExPASy pi/mW на c17orf98 https://web.expasy.org/cgi-bin/compute_pi/pi_tool [ постоянная мертвая ссылка ]
- ^ Запись PSort II на C17orf98 [ постоянная мертвая ссылка ]
- ^ GPS-запись Bio Cuckoo на C17orf98 http://gps.biocu [ постоянная мертвая ссылка ]
- ^ Запись GPS-сумо на c17orf98.
- ^ Запись ИньОян на c17orf98 http://www.cbs.dtu.dk/services/YinOYang/
- ^ Ганновер, Джон А.; Краузе, Майкл В.; С любовью, Дона К. (2010). «Сигнальный путь гексозамина: цикл O-GlcNAc во время праздника или голода» . Biochimica et Biophysical Acta (BBA) – Общие предметы . 1800 (2): 80–95. дои : 10.1016/j.bbagen.2009.07.017 . ПМК 2815088 . ПМИД 19647043 .
- ^ Jump up to: а б с Поиск эукариотического линейного мотива по аминокислотной последовательности c17orf98
- ^ Эно С., Браун Р.К., Шен З.Дж., Сян З., Хенингер Э., Лав Р.Б., Сандор М., Мальтер Дж.С. (февраль 2007 г.). «Pin1 модулирует иммунный ответ 1 типа» . ПЛОС ОДИН . 2 (2): е226. Бибкод : 2007PLoSO...2..226E . дои : 10.1371/journal.pone.0000226 . ПМК 1790862 . ПМИД 17311089 .
- ^ Запись BioGrid на c17orf98
- ^ Запись MINT на c17orf98
- ^ Запись STRING на C17orf98
- ^ PSICQUIC Посмотреть запись на c17orf98
- ^ запись pax-db на c17orf98 https://pax-db.org/protein/1858623#
- ^ «Данные микрочипа :: Атлас мозга Аллена: Человеческий мозг» . human.brain-map.org . Проверено 06 мая 2018 г.
- ^ Запись в Атласе белков человека (сигма) на c17orf98 https://www.proteinatlas.org/ENSG00000275489-C17orf98/cell [ постоянная мертвая ссылка ]
- ^ Запись Sigma Aldrich на c17orf98 https://www.sigmaaldrich.com/catalog/product/sigma/hpa051696?lang=en®ion=US
- ^ Запись PSORT II об аминокислотной последовательности c17orf98 https://psort.hgc.jp/form2.html
- ^ Запись PSort II на C17orf98 https://psort.hgc.jp/cgi-bin/runpsort.pl [ постоянная мертвая ссылка ]
- ^ Запись DTU по биоинформатике на c17orf98.
- ^ Запись Expasy Sosui на C17orf98
- ^ Запись в Атласе белков на c17orf98.
- ^ Запись NCBI Unigene на c17orf98 www.ncbi.nlm.nih.gov/UniGene/clust.cgi?UGID=169593&TAXID=9606&SEARCH=c17orf98
- ^ «Запись био-GPS на c17orf98» .
- ^ Запись в Атласе белков человека (сигма) на c17orf98 https://www.proteinatlas.org/ENSG00000275489-C17orf98/cell
- ^ Запись GeoProfiles NCBI на c17orf98 https://www.ncbi.nlm.nih.gov/geoprofiles
- ^ Запись о фосфосите на c17orf98 https://www.posite.org/proteinAction.action?id=5156341&showAllSites=true
- ^ «C17orf98 — Тератозооспермия (HG-U133 2.0)» .
- ^ «Запись GeoProfiles NCBI на c17orf98» .
- ^ «Молекулярные часы и оценка дивергенции видов — изучайте науку в Scitable» . www.nature.com . Проверено 2 мая 2018 г.
- ^ Взрывная запись о c17orf98 https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins