Хорошо
| Имена | |
|---|---|
| Название ИЮПАК
1-(3,7,12,16,20,24-гексаметилпентакозил)-2,3,4-триметилбензол
| |
| Другие имена
хи,пси-каротан
| |
| Идентификаторы | |
3D model ( JSmol )
|
|
ПабХим CID
|
|
| Характеристики | |
| С 40 Ч 74 | |
| Молярная масса | 555.032 g·mol −1 |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа).
| |
Окенан , диагенетический конечный продукт окенона, является Chromatiaceae , биомаркером пурпурных серобактерий . [ 1 ] Эти аноксигенные фототрофы используют свет для получения энергии, а сульфид — в качестве донора электронов и источника серы . Обнаружение окенана в морских отложениях предполагает существование в прошлом эвксинной среды, где водные столбы были бескислородными и сульфидными. Это потенциально чрезвычайно важно для реконструкции прошлых условий океана, но до сих пор окенан был идентифицирован только в одном образце палеопротерозойских пород (возрастом 1,6 миллиарда лет) из Северной Австралии. [ 2 ] [ 3 ]
Фон
[ редактировать ]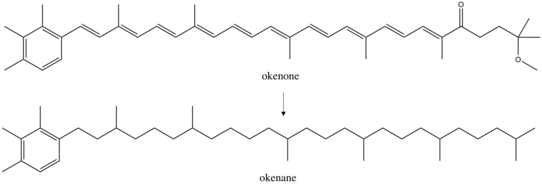

Окенон – каротиноид , [ 4 ] класс пигментов, широко распространённых в фотосинтезирующих организмах. Эти конъюгированные молекулы действуют как аксессуары в светособирающем комплексе . Известно более 600 каротиноидов, каждый из которых имеет множество функциональных групп , которые изменяют их спектр поглощения . Окенон, по-видимому, лучше всего адаптирован к желто-зеленому переходу (520 нм) видимого спектра , улавливая свет под морским планктоном в океане. Эта глубина варьируется в зависимости от структуры сообщества водной толщи. Исследование микробного цветения выявило Chromatiaceae на глубине от 1,5 до 24 метров, но более 75% из них наблюдались на глубине выше 12 метров. [ 5 ] Дальнейшие планктонные серные бактерии занимают другие ниши: зеленые серные бактерии Chlorobiaceae . , которые производят каротиноид хлоробактен , были обнаружены в наибольшем количестве на глубине выше 6 метров, тогда как зеленые серные бактерии, производящие изорениератен, преимущественно идентифицировались на глубине выше 17 метров Обнаружение любого из этих каротиноидов в древних породах может ограничить глубину перехода от кислородного к бескислородному, а также ограничить прошлую экологию . Окенан и хлоробактан, обнаруженные в австралийских палеопротерозойских образцах, позволили сделать вывод о временно неглубоком бескислородном переходе, вероятно, между 12 и 25 м. [ 2 ]
Окенон синтезируется у 12 видов Chromatiaceae, относящихся к восьми родам . Другие пурпурные серобактерии содержат ациклические каротиноидные пигменты, такие как ликопин и родопин . Однако геохимики в основном изучают окенон, поскольку он структурно уникален. Это единственный пигмент с паттерном замещения 2,3,4 триметиларила . Напротив, зеленые серобактерии продуцируют 2,3,6- триметиларилизопреноиды . [ 6 ] Синтез этих структур создает биологическую специфичность, которая может отличать экологию окружающей среды прошлого. Окенон, хлоробактен и изорениератен производятся серобактериями путем модификации ликопина . В океноне концевая группа ликопина образует χ-кольцо, а хлоробактен имеет φ-кольцо. [ 7 ] Первый этап биосинтеза этих двух пигментов аналогичен: образование β-кольца ферментом β- циклазой . Затем синтезы расходятся: фермент каротиндесатураза/ метилтрансфераза преобразует концевую группу β-кольца в χ-кольцо. Другие реакции завершают синтез до окенона: удлинение конъюгации, добавление метоксигруппы и вставка кетона . Однако биологически хорошо охарактеризованы только первые этапы синтеза.
Сохранение
[ редактировать ]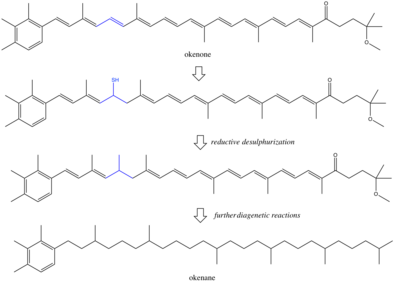
Пигменты и другие биомаркеры, вырабатываемые организмами, могут избежать микробного и химического разложения и сохраняться в осадочных породах . [ 8 ] В условиях консервации окружающая среда часто является бескислородной и восстанавливающей, что приводит к химической потере функциональных групп, таких как двойные связи и гидроксильные группы . Точные реакции во время диагенеза плохо изучены, хотя некоторые предполагают, что восстановительная десульфуризация является механизмом насыщения окенона до окенана. [ 9 ] [ 10 ] Всегда существует вероятность того, что окенан образуется в результате абиотических реакций, возможно, в результате метильных сдвигов в β-каротине . [ 11 ] Если бы эта реакция происходила, у окенана было бы несколько предшественников, и биологическая специфичность биомаркера была бы снижена. Однако маловероятно, что изомерспецифические перегруппировки двух метильных групп происходят без ферментативной активности. Большинство исследований пришли к выводу, что окенан является настоящим биомаркером пурпурных серобактерий. Однако есть и другие биологические аргументы против этой интерпретации. [ 12 ] Предыдущие организмы, синтезировавшие окенон, возможно, не были современными аналогами пурпурных серобактерий. В современном океане могут существовать и другие неохарактеризованные фотосинтезаторы, производящие окенон. Еще одним осложнением является горизонтальный перенос генов . [ 13 ] Если Chromatiaceae приобрели способность создавать окенон позже, чем в палеопротерозое, то окенан отслеживает не пурпурные серные бактерии, а скорее исходный донор гена. Эти неясности указывают на то, что интерпретация биомаркеров в горных породах возрастом в миллиарды лет будет ограничена пониманием древнего метаболизма .
Методы измерения
[ редактировать ]ГХ/МС
[ редактировать ]Перед анализом осадочных пород извлекают из органическое вещество . Обычно извлекается только менее одного процента из-за термической зрелости материнской породы. Органическое содержание часто разделяется на насыщенные , ароматические и полярные соединения . Газовая хроматография может быть объединена с масс-спектрометрией для анализа экстрагированной ароматической фракции. Соединения элюируются из колонки в зависимости от их отношения массы к заряду (M/Z) и отображаются на основе относительной интенсивности. Пики присваиваются соединениям на основе результатов поиска в библиотеке, стандартов и относительного времени удерживания . Некоторые молекулы имеют характерные пики, которые позволяют легко искать определенные отношения массы к заряду. Для триметиларилизопреноида окенана этот характерный пик наблюдается при M/Z 134.
Изотопные отношения
[ редактировать ]Соотношение изотопов углерода пурпурных и зеленых серобактерий существенно отличается от других фотосинтезирующих организмов. Биомасса пурпурных серобактерий Chromatiaceae часто обеднена δ 13 C по сравнению с типичными оксигенными фототрофами, в то время как зеленые серные бактерии Chlorobiaceae часто обогащаются. [ 14 ] Это дает дополнительную возможность определить экологические сообщества, сохранившиеся в осадочных породах. Для биомаркера окенана δ 13 C можно определить с помощью масс-спектрометра изотопного отношения .
Практический пример: Северная Австралия
[ редактировать ]В современных условиях пурпурные серобактерии процветают в меромиктических (постоянно стратифицированных) озерах. [ 15 ] и заросшие фьорды и встречаются в немногих морских экосистемах. Исключением являются гиперсоленые воды, такие как Черное море . [ 16 ] Однако миллиарды лет назад, когда океаны были бескислородными и сульфидными, фототрофные серные бактерии имели больше обитаемого пространства. Исследователи из Австралийского национального университета и Массачусетского технологического института исследовали горные породы возрастом 1,6 миллиарда лет, чтобы изучить химические условия палеопротерозойского океана . Многие полагают, что в это время были глубоко проникающие толщи кислородной воды из-за исчезновения полосчатых железных образований примерно 1,8 миллиарда лет назад. Другие, возглавляемые статьей Дональда Кэнфилда , опубликованной в 1998 году в журнале Nature , полагают, что воды были эвксинными. Исследование горных пород того времени выявило биомаркеры как пурпурных, так и зеленых серобактерий, что добавило доказательства в поддержку гипотезы Кэнфилдского океана . Проанализированное обнажение осадочных пород представляло собой формацию Барни-Крик из группы Макартур в северной Австралии. Анализ проб выявил как 2,3,6-триметиларилизопреноиды (хлорбактан) Chlorobiaceae, так и 2,3,4-триметиларилизопреноиды (окенан) Chromatiaceae. И хлоробактан, и окенан указывают на эвксиновый океан с сульфидной и бескислородной поверхностью на глубине ниже 12-25 метров. Авторы пришли к выводу, что хотя кислород и присутствовал в атмосфере, палеопротерозойские океаны не были полностью насыщены кислородом. [ 2 ]
См. также
[ редактировать ]- Аноксическое событие
- Аноксигенный фотосинтез
- Биомаркеры
- Каротиноиды
- Зеленые серные бактерии
- Пурпурные серобактерии
Ссылки
[ редактировать ]- ^ Имхофф, Йоханнес Ф. (1 января 1995 г.). «Таксономия и физиология фототрофных пурпурных бактерий и зеленых серных бактерий». В Бланкеншипе, Роберт Э .; Мэдиган, Майкл Т.; Бауэр, Карл Э. (ред.). Аноксигенные фотосинтезирующие бактерии . Достижения в области фотосинтеза и дыхания. Том. 2. Спрингер Нидерланды. стр. 1–15. дои : 10.1007/0-306-47954-0_1 . ISBN 9780792336815 .
- ^ Jump up to: а б с Брокс, Йохен Дж.; Шеффер, Филип (01 марта 2008 г.). «Окенан, биомаркер пурпурных серобактерий (Chromatiaceae) и других новых производных каротиноидов из формации Барни-Крик, образовавшейся 1640 млн лет назад». Акта геохимии и космохимии . 72 (5): 1396–1414. Бибкод : 2008GeCoA..72.1396B . дои : 10.1016/j.gca.2007.12.006 .
- ^ Брокс, Йохен Дж.; С любовью, Гордон Д.; Вызов, Роджер Э.; Нолл, Эндрю Х.; Логан, Грэм А.; Боуден, Стивен А. (2005). «Биомаркеры существования зеленых и пурпурных серных бактерий в стратифицированном палеопротерозойском море». Природа . 437 (7060): 866–870. Бибкод : 2005Natur.437..866B . дои : 10.1038/nature04068 . ПМИД 16208367 . S2CID 4427285 .
- ^ Шеффер, Филипп; Адам, Пьер; Верунг, Патрик; Альбрехт, Пьер (1 декабря 1997 г.). «Новые производные ароматических каротиноидов из серофотосинтезирующих бактерий в отложениях». Буквы тетраэдра . 38 (48): 8413–8416. дои : 10.1016/S0040-4039(97)10235-0 .
- ^ Гемерден, Ханс Ван; Мас, Хорди (1 января 1995 г.). Бланкеншип, Роберт Э.; Мэдиган, Майкл Т.; Бауэр, Карл Э. (ред.). Аноксигенные фотосинтезирующие бактерии . Достижения в области фотосинтеза и дыхания. Спрингер Нидерланды. стр. 49–85. дои : 10.1007/0-306-47954-0_4 . ISBN 9780792336815 .
- ^ Вызов, RE; Пауэлл, Т.Г. (1 марта 1987 г.). «Идентификация арилизопреноидов в нефтематеринских породах и сырой нефти: биологические маркеры зеленых серобактерий». Geochimica et Cosmochimica Acta . 51 (3): 557–566. Бибкод : 1987GeCoA..51..557S . дои : 10.1016/0016-7037(87)90069-X .
- ^ Фогль, К.; Брайант, окружной прокурор (01 мая 2012 г.). «Биосинтез биомаркера окенона: образование χ-кольца». Геобиология . 10 (3): 205–215. Бибкод : 2012Gbio...10..205V . дои : 10.1111/j.1472-4669.2011.00297.x . ISSN 1472-4669 . ПМИД 22070388 . S2CID 205627793 .
- ^ Брокс, Йохен Дж.; Грайс, Клити (1 января 2011 г.). Райтнер, Иоахим; Тиль, Волкер (ред.). Энциклопедия геобиологии . Серия Энциклопедия наук о Земле. Спрингер Нидерланды. стр. 147–167. дои : 10.1007/978-1-4020-9212-1_30 . ISBN 9781402092114 .
- ^ Хебтинг, Ю.; Шеффер, П.; Беренс, А.; Адам, П.; Шмитт, Г.; Шнекенбургер, П.; Бернаскони, С.М.; Альбрехт, П. (16 июня 2006 г.). «Биомаркерные доказательства основного пути сохранения осадочного органического углерода» . Наука . 312 (5780): 1627–1631. Бибкод : 2006Sci...312.1627H . дои : 10.1126/science.1126372 . ISSN 0036-8075 . ПМИД 16690819 . S2CID 45660326 .
- ^ Верне, Йозеф П.; Лайонс, Тимоти В.; Холландер, Дэвид Дж.; Схоутен, Стефан; Хопманс, Эллен К.; Синнингхе Дамсте, Яап С. (15 июля 2008 г.). «Исследование путей диагенетического сульфурирования органических веществ с использованием изотопного анализа серы для конкретных соединений». Geochimica et Cosmochimica Acta . 72 (14): 3489–3502. Бибкод : 2008GeCoA..72.3489W . дои : 10.1016/j.gca.2008.04.033 .
- ^ Купманс, Мартин П.; Схоутен, Стефан; Конен, Математик Э.Л.; Синнингхе Дамте, Яап С. (1 декабря 1996 г.). «Ограниченная полезность арилизопреноидов в качестве индикаторов аноксии фотозоны». Geochimica et Cosmochimica Acta . 60 (23): 4873–4876. Бибкод : 1996GeCoA..60.4873K . дои : 10.1016/S0016-7037(96)00303-1 . hdl : 1874/4427 .
- ^ Брокс, Йохен Дж.; Банфилд, Джиллиан (2009). «Разгадка древней истории микробов с помощью протеогеномики сообщества и геохимии липидов». Обзоры природы Микробиология . 7 (8): 601–609. дои : 10.1038/nrmicro2167 . ПМИД 19609261 . S2CID 20913191 .
- ^ Коббс, Кэссиди; Хит, Джереми; Стайрман III, Джон О.; Эббот, Патрик (01 августа 2013 г.). «Каротиноиды в неожиданных местах: галлицы, латеральный перенос генов и биосинтез каротиноидов у животных». Молекулярная филогенетика и эволюция . 68 (2): 221–228. дои : 10.1016/j.ympev.2013.03.012 . ПМИД 23542649 .
- ^ Зякун А.М.; Лунина, ОН; Прусакова Т.С.; Пименов, Н.В.; Иванов, М.В. (06.12.2009). «Фракционирование стабильных изотопов углерода фотоавтотрофно растущими аноксигенными пурпурными и зелеными серными бактериями». Микробиология . 78 (6): 757. doi : 10.1134/S0026261709060137 . ISSN 0026-2617 . S2CID 34430255 .
- ^ Оверманн, Йорг; Битти, Дж. Томас; Холл, Кен Дж.; Пфенниг, Норберт; Норткот, Том Г. (1 июля 1991 г.). «Характеристика плотного пурпурного слоя серобактерий в меромиктическом соленом озере» (PDF) . Лимнология и океанография . 36 (5): 846–859. Бибкод : 1991LimOc..36..846O . дои : 10.4319/lo.1991.36.5.0846 . ISSN 1939-5590 .
- ^ Хашва, Ф.А.; Трюпер, HG (1978). «Жизнеспособные фототрофные серобактерии дна Черного моря» . Гельголандские научные морские исследования . 31 (1–2): 249–253. Бибкод : 1978HWM....31..249H . дои : 10.1007/BF02297000 . ISSN 0017-9957 .