Дополнительная моторная зона
| Дополнительная моторная зона | |
|---|---|
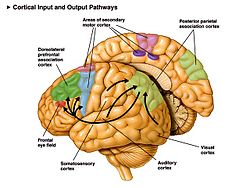 Некоторые двигательные области коры головного мозга человека. Дополнительная двигательная область показана розовым цветом. | |
 3D-визуализация дополнительной моторной коры головного мозга среднего человека | |
| Идентификаторы | |
| Нейроимена | 3176 |
| ФМА | 224858 |
| Анатомические термины нейроанатомии | |
Дополнительная двигательная область ( ДМА ) — часть моторной коры приматов, способствующая контролю движений. Он расположен на срединной поверхности полушария прямо перед (передним) представительством ноги первичной моторной коры . У обезьян СМА содержит приблизительную карту тела. У людей карта тела не выражена. Нейроны СМА проецируются непосредственно в спинной мозг и могут играть роль в прямом контроле движений. Возможные функции, приписываемые СМА, включают постуральную стабилизацию тела, координацию обеих сторон тела, например, при бимануальном действии, контроль движений, которые генерируются внутри, а не запускаются сенсорными событиями, и контроль последовательностей движений. . Все эти предложенные функции остаются гипотезами. Точная роль или роли SMA пока не известны.
Информацию об открытии СМА и ее связи с другими областями моторной коры см. в основной статье о моторной коре .
Субрегионы
[ редактировать ]По крайней мере шесть территорий теперь признаны внутри более крупного региона, который когда-то определялся как SMA. Эти подразделения наиболее подробно изучены в мозге обезьяны. Наиболее переднюю часть теперь обычно называют пре-СМА. [ 1 ] [ 2 ] [ 3 ] Он имеет редкие связи или вообще не имеет связей со спинным мозгом или первичной моторной корой и имеет обширные связи с префронтальными областями. [ 1 ] [ 4 ] [ 5 ] [ 6 ] [ 7 ]
Дополнительное поле глаза (SEF) представляет собой относительно переднюю часть SMA, которая при стимуляции вызывает движения головы и глаз и, возможно, движения конечностей и туловища. [ 8 ] [ 9 ] [ 10 ] [ 11 ]
Дум и веревка [ 5 ] На основании цитоархитектуры и связей со спинным мозгом выдвинули гипотезу, что часть СМА в поясной борозде в медиальной части полушария может быть разделена на три отдельные области, поясные двигательные области. Функции поясной двигательной области еще систематически не изучались, хотя могут быть вовлечены в эмоционально обусловленное поведение, такое как лимбический смех.
Собственно СМА у обезьян теперь ограничена областью короны полушария и частично распространяется на медиальную стенку, непосредственно перед первичным представительством двигательной ноги. Собственно СМА проецируется непосредственно на спинной мозг и, следовательно, является одной из основных зон вывода кортикальной двигательной системы. [ 5 ] [ 12 ] [ 13 ] [ 14 ] [ 15 ] [ 16 ]
Недавно Чжан и др. [ 17 ] исследовали функциональные подразделения медиальной ПФК на основе связей всего мозга, охарактеризованных на основе большого набора данных фМРТ в состоянии покоя. Помимо повторения границ между СМА и преСМА, текущие результаты подтверждают функциональные различия между задней и передней пре-СМА. В отличие от задней пре-СМА, передняя пре-СМА связана с большей частью префронтальных, но не соматомоторных областей. В целом, SMA тесно связана с таламусом и эпиталамусом , задняя пре-SMA с скорлупой , паллидумом и STN, а передняя пре-SMA с хвостатым ядром , при этом хвостатое ядро демонстрирует значительную асимметрию полушарий.
Функции
[ редактировать ]Пенфилд и Уэлч [ 18 ] в 1951 году впервые описал СМА в мозгу обезьяны и в мозгу человека как представительство тела на медиальной стенке полушария. Вулси и коллеги [ 19 ] в 1952 году подтвердил СМА в мозгу обезьяны, описав ее как грубую соматотопическую карту, где ноги расположены сзади, а лицо - спереди. Было обнаружено, что изображения различных частей тела сильно перекрываются. Стимуляция многих участков вызывала двусторонние движения, а иногда и движения всех четырех конечностей. Эта перекрывающаяся соматотопическая карта при СМА была подтверждена многими другими. [ 2 ] [ 13 ] [ 20 ] [ 21 ] [ 22 ]
Были предложены четыре основные гипотезы о функции СМА: контроль постуральной устойчивости во время стоянки или ходьбы, [ 18 ] координация временной последовательности действий, [ 23 ] [ 24 ] [ 25 ] [ 26 ] [ 27 ] [ 28 ] [ 29 ] [ 30 ] бимануальная координация, [ 31 ] [ 32 ] и инициирование внутренне генерируемых, а не стимулированных движений. [ 3 ] [ 29 ] [ 30 ] [ 33 ] Данные, однако, не подтверждают исключительную роль СМА в какой-либо из этих функций. Действительно, СМА явно активна во время непоследовательных, одноручных и управляемых стимулами движений. [ 34 ]
Было показано, что у людей SMA генерирует ранний компонент Bereitschaftspotential ( BP) или потенциала готовности BP1 или BPearly. [ 35 ] Роль SMA была дополнительно подтверждена Cunnington et al. 2003, [ 36 ] показывая, что собственно СМА и пре-СМА активны до произвольного движения или действия, а также поясная двигательная область (CMA) и передняя средняя поясная извилина (aMCC). Недавно путем интеграции одновременно полученных ЭЭГ и фМРТ было показано, что SMA и aMCC имеют сильные взаимные связи, которые поддерживают активность друг друга, и что это взаимодействие опосредовано во время подготовки к движению в соответствии с амплитудой потенциала Bereitschafts. [ 37 ]
СМА в мозгу обезьяны может усиливать движение, особенно сложное, такое как лазание или прыжки. [ 38 ] [ 39 ] [ 40 ] Это предположение было основано на исследованиях, в которых стимуляция в поведенчески значимом временном масштабе вызывала сложные движения всего тела, напоминающие лазание или прыжки. Эта гипотеза согласуется с предыдущими гипотезами, включая участие СМА в стабилизации позы, во внутренних движениях, в бимануальной координации и в планировании последовательностей движений, поскольку все эти функции активно задействованы в сложной локомоции. Гипотеза локомоции является примером интерпретации моторной коры с точки зрения основного поведенческого репертуара, из которого возникают абстрактные функции контроля, - подход, подчеркнутый Грациано и его коллегами. [ 38 ]
Дополнительные изображения
[ редактировать ]-
 Дополнительная моторная кора выделена зеленым на корональных изображениях Т1 МРТ.
Дополнительная моторная кора выделена зеленым на корональных изображениях Т1 МРТ. -
 Дополнительная моторная кора выделена зеленым на сагиттальных изображениях Т1 МРТ.
Дополнительная моторная кора выделена зеленым на сагиттальных изображениях Т1 МРТ. -
 Дополнительная моторная кора выделена зеленым на поперечных изображениях Т1 МРТ.
Дополнительная моторная кора выделена зеленым на поперечных изображениях Т1 МРТ.



Ссылки
[ редактировать ]- ^ Jump up to: а б Он, SQ, Дум, Р.П. и Стрик, PL (1995). «Топографическая организация кортикоспинальных проекций лобной доли: двигательные зоны на медиальной поверхности полушария» . Дж. Нейроски . 15 (5): 3284–3306. doi : 10.1523/JNEUROSCI.15-05-03284.1995 . ПМК 6578253 . ПМИД 7538558 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Луппино Г., Мателли М., Камарда Р.М., Галлезе В. и Риццолатти Г. (1991). «Множественные представления движений тела в мезиальной области 6 и прилегающей поясной извилине: исследование внутрикорковой микростимуляции на макаках». Дж. Комп. Нейрол . 311 (4): 463–482. дои : 10.1002/cne.903110403 . ПМИД 1757598 . S2CID 25297539 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Мацузака Ю., Айзава Х. и Танджи Дж. (1992). «Ростра моторной области к дополнительной моторной зоне (преддополнительная моторная область) у обезьяны: активность нейронов во время выученной двигательной задачи». Дж. Нейрофизиология . 68 (3): 653–662. дои : 10.1152/jn.1992.68.3.653 . ПМИД 1432040 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бейтс, Дж. Ф. и Голдман-Ракич, П. С. (1993). «Префронтальные соединения медиальных двигательных областей у макаки-резуса». Дж. Комп. Нейрол . 336 (2): 211–228. дои : 10.1002/cne.903360205 . ПМИД 7503997 . S2CID 35838076 .
- ^ Jump up to: а б с Дум, Р.П. и Стрик, PL (1991). «Происхождение кортикоспинальных проекций из премоторных областей лобной доли» . Дж. Нейроски . 11 (3): 667–689. doi : 10.1523/JNEUROSCI.11-03-00667.1991 . ПМК 6575356 . ПМИД 1705965 .
- ^ Лу, М.Т., Престон, Дж.Б. и Стрик, П.Л. (1994). «Взаимосвязь между префронтальной корой и премоторными областями лобной доли». Дж. Комп. Нейрол . 341 (3): 375–392. дои : 10.1002/cne.903410308 . ПМИД 7515081 . S2CID 13045173 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Луппино Г., Мателли М., Камарда Р. и Риццолатти Г. (1993). «Кортикокортикальные связи области F3 (собственно СМА) и области F6 (пре-СМА) у макаки». Дж. Комп. Нейрол . 338 (1): 114–140. дои : 10.1002/cne.903380109 . ПМИД 7507940 . S2CID 6171579 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Чен, Л.Л. и Уолтон, М.М. (2005). «Движение головы, вызванное электрической стимуляцией дополнительного поля глаза макаки-резуса». Дж. Нейрофизиология . 94 (6): 4502–4519. дои : 10.1152/jn.00510.2005 . ПМИД 16148273 .
- ^ Руссо, Г.С. и Брюс, СиДжей (2000). «Дополнительное поле глаза: представление саккад и взаимосвязь между полями нервных реакций и вызванными движениями глаз». Дж. Нейрофизиология . 84 (5): 2605–2621. дои : 10.1152/jn.2000.84.5.2605 . ПМИД 11068002 . S2CID 16208146 .
- ^ Шлаг Дж. и Шлаг-Рей М. (1987). «Доказательства дополнительного поля зрения». Дж. Нейрофизиология . 57 (1): 179–200. дои : 10.1152/jn.1987.57.1.179 . ПМИД 3559671 .
- ^ Теховник, Э.Дж. и Ли, К. (1993). «Дорсомедиальная лобная кора макаки-резуса: топографическое изображение саккад, вызываемых электрической стимуляцией». Эксп. Мозговой Рес . 96 (3): 430–442. дои : 10.1007/bf00234111 . ПМИД 8299745 . S2CID 11322416 .
- ^ Галеа, член парламента, и Дариан-Смит, я (1994). «Множественные популяции кортикоспинальных нейронов у макак характеризуются уникальным корковым происхождением, спинальными окончаниями и соединениями». Цереб. Кортекс . 4 (2): 166–194. дои : 10.1093/cercor/4.2.166 . ПМИД 8038567 .
- ^ Jump up to: а б Макферсон Дж., Марангоз К., Майлз Т.С. и Визендангер М. (1982). «Микростимуляция дополнительной двигательной области (SMA) у бодрствующей обезьяны». Эксп. Мозговой Рес . 45 (3): 410–416. дои : 10.1007/bf01208601 . ПМИД 7067775 . S2CID 10053006 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мюррей, Э.А. и Коултер, доктор юридических наук (1981). «Организация кортикоспинальных нейронов у обезьяны». Дж. Комп. Нейрол . 195 (2): 339–365. дои : 10.1002/cne.901950212 . ПМИД 7251930 . S2CID 20215391 .
- ^ Нудо, Р.Дж. и Мастертон, Р.Б. (1990). «Нисходящие пути спинного мозга, III: Места начала кортикоспинального тракта». Дж. Комп. Нейрол . 296 (4): 559–583. дои : 10.1002/cne.902960405 . ПМИД 2113540 . S2CID 20563396 .
- ^ Тоёсима, К. и Сакаи, Х. (1982). «Точная корковая протяженность происхождения кортикоспинального тракта (CST) и количественный вклад в CST в различных цитоархитектонических областях. Исследование пероксидазы хрена на обезьянах». Дж. Хирнфорш . 23 (3): 257–269. ПМИД 7130676 .
- ^ Чжан С., Иде Дж. С. и Ли К. С. (2012). «Функциональная связность медиальной верхней лобной коры в состоянии покоя» . Цереб. Кортекс . 22 (1): 99–111. дои : 10.1093/cercor/bhr088 . ПМЦ 3236794 . ПМИД 21572088 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Пенфилд, В. и Уэлч, К. (1951). «Дополнительная двигательная область коры головного мозга: клиническое и экспериментальное исследование». АМА Арх. Нейрол. Психиатрия . 66 (3): 289–317. doi : 10.1001/archneurpsyc.1951.02320090038004 . ПМИД 14867993 .
- ^ Вулси, К.Н., Сеттлейдж, П.Х., Мейер, Д.Р., Сенсер, В., Хамуи, Т.П. и Трэвис, А.М. (1952). «Схема локализации в прецентральных и «дополнительных» двигательных областях и их отношение к понятию премоторной области». Ассоциация исследований нервных и психических заболеваний . 30 . Нью-Йорк, штат Нью-Йорк: Raven Press: 238–264.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Гулд, Х.Дж. III, Кьюсик, К.Г., Понс, Т.П. и Каас, Дж.Х. (1996). «Взаимосвязь соединений мозолистого тела с картами электрической стимуляции моторики, дополнительной моторики и лобных полей глаз у совиных обезьян». Дж. Комп. Нейрол . 247 (3): 297–325. дои : 10.1002/cne.902470303 . ПМИД 3722441 . S2CID 21185898 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Муаккасса, К.Ф. и Стрик, PL (1979). «Вводы лобных долей в моторную кору приматов: свидетельства существования четырех соматотопически организованных «премоторных» областей». Мозговой Рес . 177 (1): 176–182. дои : 10.1016/0006-8993(79)90928-4 . ПМИД 115545 . S2CID 34047716 .
- ^ Митц, А.Р. и Уайз, СП (1987). «Соматотопическая организация дополнительной двигательной области: картирование внутрикорковой микростимуляции» . Дж. Нейроски . 7 (4): 1010–1021. doi : 10.1523/JNEUROSCI.07-04-01010.1987 . ПМК 6568999 . ПМИД 3572473 .
- ^ Гаймар Б., Пьеро=Дезейлиньи К. и Риво С. (1990). «Нарушение последовательностей саккад, управляемых памятью, после дополнительных поражений двигательной области». Анналы неврологии . 28 (5): 622–626. дои : 10.1002/ana.410280504 . ПМИД 2260848 . S2CID 31214017 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Герлофф К., Корвелл Б., Чен Р., Халлетт М. и Коэн Л.Г. (1997). «Стимуляция дополнительной двигательной области человека мешает организации будущих элементов в сложных двигательных последовательностях» . Мозг . 120 (9): 1587–1602. дои : 10.1093/мозг/120.9.1587 . ПМИД 9313642 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Дженкинс, И.Х., Брукс, Д.Д., Никсон, П.Д., Фраковяк, Р.С. и Пассингем, Р.Э. (1994). «Обучение двигательной последовательности: исследование с помощью позитронно-эмиссионной томографии» . Дж. Нейроски . 14 (6): 3775–3790. doi : 10.1523/JNEUROSCI.14-06-03775.1994 . ПМК 6576955 . ПМИД 8207487 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ли, Д. и Кесси, С. (2003). «Активность в дополнительной двигательной области, связанная с обучением и производительностью во время последовательного зрительно-моторного задания». Дж. Нейрофизиология . 89 (2): 1039–1056. дои : 10.1152/jn.00638.2002 . ПМИД 12574479 .
- ^ Мушиаке Х., Инасе М. и Танджи Дж. (1990). «Селективное кодирование моторной последовательности в дополнительной моторной области коры головного мозга обезьяны». Эксп. Мозговой Рес . 82 (1): 208–210. дои : 10.1007/bf00230853 . ПМИД 2257906 . S2CID 33522843 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Шима К. и Танджи Дж. (1998). «Как дополнительные, так и дополнительные двигательные области имеют решающее значение для временной организации множества движений». Дж. Нейрофизиология . 80 (6): 3247–3260. дои : 10.1152/jn.1998.80.6.3247 . ПМИД 9862919 .
- ^ Jump up to: а б Роланд П.Е., Ларсен Б., Лассен Н.А. и Скинхой Э. (1980). «Дополнительная двигательная область и другие области коры в организации произвольных движений человека». Дж. Нейрофизиология . 43 (1): 118–136. дои : 10.1152/jn.1980.43.1.118 . ПМИД 7351547 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Роланд П.Е., Скинхой Э., Лассен Н.А. и Ларсен Б. (1980). «Различные области коры человека в организации произвольных движений во внеличностном пространстве». Дж. Нейрофизиология . 43 (1): 137–150. дои : 10.1152/jn.1980.43.1.137 . ПМИД 7351548 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бринкман, К. (1981). «Поражения в дополнительной двигательной области мешают обезьяне выполнять задачу бимануальной координации». Неврология. Летт . 27 (3): 267–270. дои : 10.1016/0304-3940(81)90441-9 . ПМИД 7329632 . S2CID 41060226 .
- ^ Серриен, DJ, Стренс, Л.Х., Оливейро, А. и Браун, П. (2002). «Повторяющаяся транскраниальная магнитная стимуляция дополнительной двигательной области (SMA) ухудшает бимануальный контроль движений у людей». Неврология. Летт . 328 (2): 89–92. дои : 10.1016/s0304-3940(02)00499-8 . ПМИД 12133562 . S2CID 35568770 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Халсбанд У., Мацузака Ю. и Танджи Дж. (1994). «Нейрональная активность в дополнительной, преддополнительной и премоторной коре приматов во время последовательных движений, управляемых извне и внутри». Неврология. Рез . 20 (2): 149–155. дои : 10.1016/0168-0102(94)90032-9 . ПМИД 7808697 . S2CID 5930996 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Пикард, Н. и Стрик, PL (сентябрь 2003 г.). «Активация дополнительной двигательной области (ДМА) при выполнении зрительно управляемых движений» . Цереб. Кортекс . 13 (9): 977–986. дои : 10.1093/cercor/13.9.977 . ПМИД 12902397 .
- ^ Дикке Л., Корнхубер (1978). «дополнительная» моторная кора при произвольных движениях пальцев человека». Brain Res . 159 (2): 473–476. : 10.1016 /0006-8993(78)90561-9 . PMID 728816. . S2CID 43904948 doi
- ^ Каннингтон Р., Виндишбергер С., Дике Л., Мозер Э. (2003). «Подготовка и готовность к добровольному движению: высокопольное фМРТ-исследование реакции Bereitschafts-BOLD». НейроИмидж . 20 (1): 404–412. дои : 10.1016/s1053-8119(03)00291-x . ПМИД 14527600 . S2CID 13419573 .
- ^ Нгуен В.Т., Брейкспир М., Каннингтон Р. (2014). «Взаимные взаимодействия СМА и поясной извилины поддерживают преддвигательную активность для произвольных действий» . Дж. Нейроски . 34 (49): 16397–16407. doi : 10.1523/jneurosci.2571-14.2014 . ПМК 6608485 . ПМИД 25471577 .
- ^ Jump up to: а б Грациано, MSA (2008). Интеллектуальная машина движения . Оксфорд, Великобритания: Издательство Оксфордского университета.
- ^ Грациано, MSA и Афлало, Теннесси (2007). «Отображение поведенческого репертуара в коре головного мозга» (PDF) . Нейрон . 56 (2): 239–251. дои : 10.1016/j.neuron.2007.09.013 . ПМИД 17964243 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Грациано, MSA , Афлало, Теннесси и Кук, Д.Ф. (2005). «Движения рук, вызванные электрической стимуляцией моторной коры обезьян». Дж. Нейрофизиология . 94 (6): 4209–4223. дои : 10.1152/Jn.01303.2004 . ПМИД 16120657 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )
Дальнейшее чтение
[ редактировать ]- Принципы нейронауки (2000), 4-е изд., Кандель и др.
- Дебаер Ф., Вендерот Н., Сунаерт С., Ван-Хекке П., Суиннен С.П. (июль 2003 г.). «Внутренняя и внешняя генерация движений: дифференциальные нейронные пути, участвующие в бимануальной координации, выполняемые при наличии или отсутствии усиленной зрительной обратной связи». НейроИмидж . 19 (3): 764–76. дои : 10.1016/s1053-8119(03)00148-4 . ПМИД 12880805 . S2CID 12977852 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - Воробьев; и др. (1998). «Парцелляция мезиальной области 6 человека: цитоархитектонические данные для трех отдельных областей». Eur J Neurosci . 10 (6): 2199–203. дои : 10.1046/j.1460-9568.1998.00236.x . ПМИД 9753106 . S2CID 23787668 .