Дополнительная моторная зона
| Дополнительная моторная зона | |
|---|---|
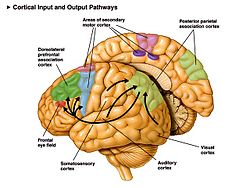 Некоторые двигательные области коры головного мозга человека. Дополнительная двигательная область показана розовым цветом. | |
 3D-визуализация дополнительной моторной коры головного мозга среднего человека | |
| Идентификаторы | |
| Нейроимена | 3176 |
| ФМА | 224858 |
| Анатомические термины нейроанатомии | |
Дополнительная двигательная область ( СМА ) — часть моторной коры приматов, способствующая контролю движений. Он расположен на срединной поверхности полушария прямо перед (передним) представительством ноги первичной моторной коры . У обезьян СМА содержит приблизительную карту тела. У людей карта тела не выражена. Нейроны СМА проецируются непосредственно в спинной мозг и могут играть роль в прямом контроле движений. Возможные функции, приписываемые СМА, включают постуральную стабилизацию тела, координацию обеих сторон тела, например, при бимануальном действии, контроль движений, которые генерируются внутри, а не запускаются сенсорными событиями, и контроль последовательностей движений. . Все эти предложенные функции остаются гипотезами. Точная роль или роли SMA пока не известны.
Информацию об открытии SMA и ее связи с другими моторными областями коры см. в основной статье о моторной коре .
Субрегионы
[ редактировать ]По крайней мере шесть территорий теперь признаны внутри более крупного региона, который когда-то определялся как SMA. Эти подразделения наиболее подробно изучены в мозге обезьяны. Наиболее переднюю часть теперь обычно называют пре-СМА. [1] [2] [3] Он имеет редкие связи или вообще не имеет связей со спинным мозгом или первичной моторной корой и имеет обширные связи с префронтальными областями. [1] [4] [5] [6] [7]
Дополнительное поле глаза (SEF) представляет собой относительно переднюю часть SMA, которая при стимуляции вызывает движения головы и глаз и, возможно, движения конечностей и туловища. [8] [9] [10] [11]
Дум и Стрик [5] На основании цитоархитектуры и связей со спинным мозгом выдвинули гипотезу, что часть СМА в поясной борозде в медиальной части полушария может быть разделена на три отдельные области, поясные двигательные области. Функции поясной двигательной области еще систематически не изучались, хотя могут быть вовлечены в эмоционально обусловленное поведение, такое как лимбический смех.
Собственно СМА у обезьян теперь ограничена областью короны полушария и частично распространяется на медиальную стенку, непосредственно перед первичным представительством двигательной ноги. Собственно СМА проецируется непосредственно на спинной мозг и, следовательно, является одной из основных зон вывода кортикальной двигательной системы. [5] [12] [13] [14] [15] [16]
Недавно Чжан и др. [17] исследовали функциональные подразделения медиальной ПФК на основе связей всего мозга, охарактеризованных на основе большого набора данных фМРТ в состоянии покоя. Помимо повторения границ между СМА и преСМА, текущие результаты подтверждают функциональные различия между задней и передней пре-СМА. В отличие от задней пре-СМА, передняя пре-СМА связана с большей частью префронтальных, но не соматомоторных областей. В целом, SMA тесно связана с таламусом и эпиталамусом , задняя пре-SMA с скорлупой , паллидумом и STN, а передняя пре-SMA с хвостатым ядром , при этом хвостатое ядро демонстрирует значительную асимметрию полушарий.
Функции
[ редактировать ]Пенфилд и Уэлч [18] в 1951 году впервые описал СМА в мозгу обезьяны и в мозгу человека как представительство тела на медиальной стенке полушария. Вулси и коллеги [19] в 1952 году подтвердил СМА в мозгу обезьяны, описав ее как грубую соматотопическую карту, на которой ноги расположены сзади, а лицо — спереди. Было обнаружено, что изображения различных частей тела сильно перекрываются. Стимуляция многих участков вызывала двусторонние движения, а иногда и движения всех четырех конечностей. Эта перекрывающаяся соматотопическая карта при СМА была подтверждена многими другими. [2] [13] [20] [21] [22]
Были предложены четыре основные гипотезы функции СМА: контроль постуральной устойчивости во время стоянки или ходьбы, [18] координация временной последовательности действий, [23] [24] [25] [26] [27] [28] [29] [30] бимануальная координация, [31] [32] и инициирование внутренне генерируемых, а не стимулированных движений. [3] [29] [30] [33] Данные, однако, не подтверждают исключительную роль СМА в какой-либо из этих функций. Действительно, СМА явно активна во время непоследовательных, одноручных и управляемых стимулами движений. [34]
Было показано, что у людей SMA генерирует ранний компонент Bereitschaftspotential ( BP) или потенциала готовности BP1 или BPearly. [35] Роль SMA была дополнительно подтверждена Cunnington et al. 2003, [36] показывая, что собственно СМА и пре-СМА активны до произвольного движения или действия, а также поясная двигательная область (CMA) и передняя средняя поясная извилина (aMCC). Недавно путем интеграции одновременно полученных ЭЭГ и фМРТ было показано, что SMA и aMCC имеют сильные взаимные связи, которые поддерживают активность друг друга, и что это взаимодействие опосредовано во время подготовки к движению в соответствии с амплитудой потенциала Bereitschafts. [37]
СМА в мозгу обезьяны может усиливать движение, особенно сложное, такое как лазание или прыжки. [38] [39] [40] Это предположение было основано на исследованиях, в которых стимуляция в поведенчески значимом временном масштабе вызывала сложные движения всего тела, напоминающие лазание или прыжки. Эта гипотеза согласуется с предыдущими гипотезами, включая участие СМА в стабилизации позы, во внутренних движениях, в бимануальной координации и в планировании последовательностей движений, поскольку все эти функции активно задействованы в сложной локомоции. Гипотеза локомоции является примером интерпретации моторной коры с точки зрения основного поведенческого репертуара, из которого возникают абстрактные функции контроля, - подход, подчеркнутый Грациано и его коллегами. [38]
Дополнительные изображения
[ редактировать ] Дополнительная моторная кора выделена зеленым на корональных изображениях Т1 МРТ.
Дополнительная моторная кора выделена зеленым на корональных изображениях Т1 МРТ. Дополнительная моторная кора выделена зеленым на сагиттальных изображениях Т1 МРТ.
Дополнительная моторная кора выделена зеленым на сагиттальных изображениях Т1 МРТ. Дополнительная моторная кора выделена зеленым на поперечных изображениях Т1 МРТ.
Дополнительная моторная кора выделена зеленым на поперечных изображениях Т1 МРТ.



Ссылки
[ редактировать ]- ^ Jump up to: а б Он, SQ, Дум, Р.П. и Стрик, PL (1995). «Топографическая организация кортикоспинальных проекций лобной доли: двигательные зоны на медиальной поверхности полушария» . Дж. Нейроски . 15 (5): 3284–3306. doi : 10.1523/JNEUROSCI.15-05-03284.1995 . ПМК 6578253 . ПМИД 7538558 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Луппино Г., Мателли М., Камарда Р.М., Галлезе В. и Риццолатти Г. (1991). «Множественные представления движений тела в мезиальной области 6 и прилегающей поясной извилине: исследование внутрикорковой микростимуляции на макаках». Дж. Комп. Нейрол . 311 (4): 463–482. дои : 10.1002/cne.903110403 . ПМИД 1757598 . S2CID 25297539 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Мацузака Ю., Айзава Х. и Танджи Дж. (1992). «Ростра моторной области к дополнительной моторной зоне (преддополнительная моторная область) у обезьяны: активность нейронов во время выученной двигательной задачи». Дж. Нейрофизиология . 68 (3): 653–662. дои : 10.1152/jn.1992.68.3.653 . ПМИД 1432040 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бейтс, Дж. Ф. и Голдман-Ракич, П. С. (1993). «Префронтальные соединения медиальных двигательных областей у макаки-резуса». Дж. Комп. Нейрол . 336 (2): 211–228. дои : 10.1002/cne.903360205 . ПМИД 7503997 . S2CID 35838076 .
- ^ Jump up to: а б с Дум, Р.П. и Стрик, PL (1991). «Происхождение кортикоспинальных проекций из премоторных областей лобной доли» . Дж. Нейроски . 11 (3): 667–689. doi : 10.1523/JNEUROSCI.11-03-00667.1991 . ПМК 6575356 . ПМИД 1705965 .
- ^ Лу, М.Т., Престон, Дж.Б. и Стрик, П.Л. (1994). «Взаимосвязь между префронтальной корой и премоторными областями лобной доли». Дж. Комп. Нейрол . 341 (3): 375–392. дои : 10.1002/cne.903410308 . ПМИД 7515081 . S2CID 13045173 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Луппино Г., Мателли М., Камарда Р. и Риццолатти Г. (1993). «Кортикокортикальные связи области F3 (собственно СМА) и области F6 (пре-СМА) у макаки». Дж. Комп. Нейрол . 338 (1): 114–140. дои : 10.1002/cne.903380109 . ПМИД 7507940 . S2CID 6171579 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Чен, Л.Л. и Уолтон, М.М. (2005). «Движение головы, вызванное электрической стимуляцией дополнительного поля глаза макаки-резуса». Дж. Нейрофизиология . 94 (6): 4502–4519. дои : 10.1152/jn.00510.2005 . ПМИД 16148273 .
- ^ Руссо, Г.С. и Брюс, СиДжей (2000). «Дополнительное поле глаза: представление саккад и взаимосвязь между полями нервных реакций и вызванными движениями глаз». Дж. Нейрофизиология . 84 (5): 2605–2621. дои : 10.1152/jn.2000.84.5.2605 . ПМИД 11068002 . S2CID 16208146 .
- ^ Шлаг Дж. и Шлаг-Рей М. (1987). «Доказательства дополнительного поля зрения». Дж. Нейрофизиология . 57 (1): 179–200. дои : 10.1152/jn.1987.57.1.179 . ПМИД 3559671 .
- ^ Теховник, Э.Дж. и Ли, К. (1993). «Дорсомедиальная лобная кора макаки-резуса: топографическое изображение саккад, вызываемых электрической стимуляцией». Эксп. Мозговой Рес . 96 (3): 430–442. дои : 10.1007/bf00234111 . ПМИД 8299745 . S2CID 11322416 .
- ^ Галеа, член парламента, и Дариан-Смит, я (1994). «Множественные популяции кортикоспинальных нейронов у макак характеризуются уникальным корковым происхождением, спинальными окончаниями и соединениями». Цереб. Кортекс . 4 (2): 166–194. дои : 10.1093/cercor/4.2.166 . ПМИД 8038567 .
- ^ Jump up to: а б Макферсон Дж., Марангоз К., Майлз Т.С. и Визендангер М. (1982). «Микростимуляция дополнительной двигательной области (SMA) у бодрствующей обезьяны». Эксп. Мозговой Рес . 45 (3): 410–416. дои : 10.1007/bf01208601 . ПМИД 7067775 . S2CID 10053006 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мюррей, Э.А. и Коултер, доктор юридических наук (1981). «Организация кортикоспинальных нейронов у обезьяны». Дж. Комп. Нейрол . 195 (2): 339–365. дои : 10.1002/cne.901950212 . ПМИД 7251930 . S2CID 20215391 .
- ^ Нудо, Р.Дж. и Мастертон, Р.Б. (1990). «Нисходящие пути спинного мозга, III: Места начала кортикоспинального тракта». Дж. Комп. Нейрол . 296 (4): 559–583. дои : 10.1002/cne.902960405 . ПМИД 2113540 . S2CID 20563396 .
- ^ Тоёсима, К. и Сакаи, Х. (1982). «Точная корковая протяженность происхождения кортикоспинального тракта (CST) и количественный вклад в CST в различных цитоархитектонических областях. Исследование пероксидазы хрена на обезьянах». Дж. Хирнфорш . 23 (3): 257–269. ПМИД 7130676 .
- ^ Чжан С., Иде Дж. С. и Ли К. С. (2012). «Функциональная связность медиальной верхней лобной коры в состоянии покоя» . Цереб. Кортекс . 22 (1): 99–111. дои : 10.1093/cercor/bhr088 . ПМЦ 3236794 . ПМИД 21572088 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Пенфилд, В. и Уэлч, К. (1951). «Дополнительная двигательная область коры головного мозга: клиническое и экспериментальное исследование». АМА Арх. Нейрол. Психиатрия . 66 (3): 289–317. doi : 10.1001/archneurpsyc.1951.02320090038004 . ПМИД 14867993 .
- ^ Вулси, К.Н., Сеттлейдж, П.Х., Мейер, Д.Р., Сенсер, В., Хамуи, Т.П. и Трэвис, А.М. (1952). «Схема локализации в прецентральных и «дополнительных» двигательных областях и их отношение к понятию премоторной области». Ассоциация исследований нервных и психических заболеваний . 30 . Нью-Йорк, штат Нью-Йорк: Raven Press: 238–264.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Гулд, Х.Дж. III, Кьюсик, К.Г., Понс, Т.П. и Каас, Дж.Х. (1996). «Взаимосвязь соединений мозолистого тела с картами электрической стимуляции моторики, дополнительной моторики и лобных полей глаз у совиных обезьян». Дж. Комп. Нейрол . 247 (3): 297–325. дои : 10.1002/cne.902470303 . ПМИД 3722441 . S2CID 21185898 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Муаккасса, К.Ф. и Стрик, PL (1979). «Вводы лобных долей в моторную кору приматов: свидетельства существования четырех соматотопически организованных «премоторных» областей». Мозговой Рес . 177 (1): 176–182. дои : 10.1016/0006-8993(79)90928-4 . ПМИД 115545 . S2CID 34047716 .
- ^ Митц, А.Р. и Уайз, СП (1987). «Соматотопическая организация дополнительной двигательной области: картирование внутрикорковой микростимуляции» . Дж. Нейроски . 7 (4): 1010–1021. doi : 10.1523/JNEUROSCI.07-04-01010.1987 . ПМК 6568999 . ПМИД 3572473 .
- ^ Гаймар Б., Пьеро=Дезейлиньи К. и Риво С. (1990). «Нарушение последовательностей саккад, управляемых памятью, после дополнительных поражений двигательной области». Анналы неврологии . 28 (5): 622–626. дои : 10.1002/ana.410280504 . ПМИД 2260848 . S2CID 31214017 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Герлофф К., Корвелл Б., Чен Р., Халлетт М. и Коэн Л.Г. (1997). «Стимуляция дополнительной двигательной области человека мешает организации будущих элементов в сложных двигательных последовательностях» . Мозг . 120 (9): 1587–1602. дои : 10.1093/мозг/120.9.1587 . ПМИД 9313642 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Дженкинс, И.Х., Брукс, Д.Д., Никсон, П.Д., Фраковяк, Р.С. и Пассингем, Р.Э. (1994). «Обучение двигательной последовательности: исследование с помощью позитронно-эмиссионной томографии» . Дж. Нейроски . 14 (6): 3775–3790. doi : 10.1523/JNEUROSCI.14-06-03775.1994 . ПМК 6576955 . ПМИД 8207487 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ли, Д. и Кесси, С. (2003). «Активность в дополнительной двигательной области, связанная с обучением и производительностью во время последовательного зрительно-моторного задания». Дж. Нейрофизиология . 89 (2): 1039–1056. дои : 10.1152/jn.00638.2002 . ПМИД 12574479 .
- ^ Мушиаке Х., Инасе М. и Танджи Дж. (1990). «Селективное кодирование моторной последовательности в дополнительной моторной области коры головного мозга обезьяны». Эксп. Мозговой Рес . 82 (1): 208–210. дои : 10.1007/bf00230853 . ПМИД 2257906 . S2CID 33522843 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Шима К. и Танджи Дж. (1998). «Как дополнительные, так и дополнительные двигательные области имеют решающее значение для временной организации множества движений». Дж. Нейрофизиология . 80 (6): 3247–3260. дои : 10.1152/jn.1998.80.6.3247 . ПМИД 9862919 .
- ^ Jump up to: а б Роланд П.Е., Ларсен Б., Лассен Н.А. и Скинхой Э. (1980). «Дополнительная двигательная область и другие области коры в организации произвольных движений человека». Дж. Нейрофизиология . 43 (1): 118–136. дои : 10.1152/jn.1980.43.1.118 . ПМИД 7351547 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Роланд П.Е., Скинхой Э., Лассен Н.А. и Ларсен Б. (1980). «Различные области коры человека в организации произвольных движений во внеличностном пространстве». Дж. Нейрофизиология . 43 (1): 137–150. дои : 10.1152/jn.1980.43.1.137 . ПМИД 7351548 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бринкман, К. (1981). «Поражения в дополнительной двигательной области мешают обезьяне выполнять задачу бимануальной координации». Неврология. Летт . 27 (3): 267–270. дои : 10.1016/0304-3940(81)90441-9 . ПМИД 7329632 . S2CID 41060226 .
- ^ Серриен, DJ, Стренс, Л.Х., Оливейро, А. и Браун, П. (2002). «Повторяющаяся транскраниальная магнитная стимуляция дополнительной двигательной области (SMA) ухудшает бимануальный контроль движений у людей». Неврология. Летт . 328 (2): 89–92. дои : 10.1016/s0304-3940(02)00499-8 . ПМИД 12133562 . S2CID 35568770 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Халсбанд У., Мацузака Ю. и Танджи Дж. (1994). «Нейрональная активность в дополнительной, преддополнительной и премоторной коре приматов во время последовательных движений, управляемых извне и внутри». Неврология. Рез . 20 (2): 149–155. дои : 10.1016/0168-0102(94)90032-9 . ПМИД 7808697 . S2CID 5930996 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Пикард, Н. и Стрик, PL (сентябрь 2003 г.). «Активация дополнительной двигательной области (ДМА) при выполнении зрительно-ориентированных движений» . Цереб. Кортекс . 13 (9): 977–986. дои : 10.1093/cercor/13.9.977 . ПМИД 12902397 .
- ^ Дикке Л., Корнхубер (1978). «дополнительная» моторная кора при произвольных движениях пальцев человека». Brain Res . 159 (2): 473–476. : 10.1016 /0006-8993(78)90561-9 . PMID 728816. . S2CID 43904948 doi
- ^ Каннингтон Р., Виндишбергер С., Дике Л., Мозер Э. (2003). «Подготовка и готовность к добровольному движению: высокопольное фМРТ-исследование реакции Bereitschafts-BOLD». НейроИмидж . 20 (1): 404–412. дои : 10.1016/s1053-8119(03)00291-x . ПМИД 14527600 . S2CID 13419573 .
- ^ Нгуен В.Т., Брейкспир М., Каннингтон Р. (2014). «Взаимные взаимодействия СМА и поясной извилины поддерживают преддвигательную активность для произвольных действий» . Дж. Нейроски . 34 (49): 16397–16407. doi : 10.1523/jneurosci.2571-14.2014 . ПМК 6608485 . ПМИД 25471577 .
- ^ Jump up to: а б Грациано, MSA (2008). Интеллектуальная машина движения . Оксфорд, Великобритания: Издательство Оксфордского университета.
- ^ Грациано, MSA и Афлало, Теннесси (2007). «Отображение поведенческого репертуара в коре головного мозга» (PDF) . Нейрон . 56 (2): 239–251. дои : 10.1016/j.neuron.2007.09.013 . ПМИД 17964243 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Грациано, MSA , Афлало, Теннесси и Кук, Д.Ф. (2005). «Движения рук, вызванные электрической стимуляцией моторной коры обезьян». Дж. Нейрофизиология . 94 (6): 4209–4223. дои : 10.1152/Jn.01303.2004 . ПМИД 16120657 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )
Дальнейшее чтение
[ редактировать ]- Принципы нейронауки (2000), 4-е изд., Кандель и др.
- Дебаер Ф., Вендерот Н., Сунаерт С., Ван-Хекке П., Суиннен С.П. (июль 2003 г.). «Внутренняя и внешняя генерация движений: дифференциальные нейронные пути, участвующие в бимануальной координации, выполняемые при наличии или отсутствии усиленной зрительной обратной связи». НейроИмидж . 19 (3): 764–76. дои : 10.1016/s1053-8119(03)00148-4 . ПМИД 12880805 . S2CID 12977852 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - Воробьев; и др. (1998). «Парцелляция мезиальной области 6 человека: цитоархитектонические данные для трех отдельных областей». Eur J Neurosci . 10 (6): 2199–203. дои : 10.1046/j.1460-9568.1998.00236.x . ПМИД 9753106 . S2CID 23787668 .