Синтетическая биологическая схема
| Часть серии статей о |
| Синтетическая биология |
|---|
| Синтетические биологические схемы |
| Редактирование генома |
| Искусственные клетки |
| Ксенобиология |
| Другие темы |

1 : РНК-полимераза, 2 : репрессор, 3 : промотор, 4 : оператор, 5 : лактоза, 6 : lacZ , 7 : lacY , 8 : lacA .
Синтетические биологические схемы — это применение синтетической биологии , где биологические части внутри клетки предназначены для выполнения логических функций, имитирующих те, которые наблюдаются в электронных схемах . Обычно эти цепи подразделяются на генетические цепи , цепи РНК или белковые цепи , в зависимости от типов биомолекул , которые взаимодействуют, создавая поведение цепи. Применение всех трех типов схем варьируется от простого стимулирования производства до добавления измеримого элемента, такого как зеленый флуоресцентный белок , в существующую естественную биологическую схему , до внедрения совершенно новых систем, состоящих из многих частей. [1]
Цель синтетической биологии — создать набор настраиваемых и охарактеризованных частей или модулей, с помощью которых можно легко спроектировать и реализовать любую желаемую синтетическую биологическую схему. [2] Эти схемы могут служить методом изменения клеточных функций, создания клеточных реакций на условия окружающей среды или влияния на клеточное развитие. Внедряя рациональные, управляемые логические элементы в клеточные системы, исследователи могут использовать живые системы в качестве сконструированных « биологических машин » для выполнения широкого спектра полезных функций. [1]
История
[ редактировать ]Первой природной генной цепью, подробно изученной, был lac-оперон . В исследованиях диауктического роста E. coli на средах с двумя сахарами Жак Моно и Франсуа Жакоб обнаружили, что E.coli преимущественно потребляет более легко перерабатываемую глюкозу, прежде чем переключиться на метаболизм лактозы . Они обнаружили, что механизм, контролирующий функцию метаболического «переключения», представляет собой двухчастный механизм управления lac-опероном. Когда в клетке присутствует лактоза, фермент β-галактозидаза, вырабатывается превращающий лактозу в глюкозу или галактозу . Когда лактоза отсутствует в клетке, lac-репрессор ингибирует выработку фермента β-галактозидазы, чтобы предотвратить любые неэффективные процессы внутри клетки.
Лак-оперон используется в биотехнологической промышленности для производства рекомбинантных белков для терапевтических целей. Ген или гены продукции экзогенного белка помещаются на плазмиду под контролем lac-промотора. Первоначально клетки выращивают в среде, не содержащей лактозу и другие сахара, поэтому новые гены не экспрессируются. Как только клетки достигают определенной точки своего роста, изопропил β-D-1-тиогалактопиранозид (ИПТГ) добавляют . IPTG, молекула, похожая на лактозу, но с серной связью, которая не гидролизуется и поэтому E. coli не переваривает ее, используется для активации или « индуцирования » производства нового белка. После индукции клеток трудно удалить IPTG из клеток и, следовательно, трудно остановить экспрессию.
Два ранних примера синтетических биологических цепей были опубликованы в журнале Nature в 2000 году. Один из них, написанный Тимом Гарднером, Чарльзом Кантором и Джимом Коллинзом , работающими в Бостонском университете , продемонстрировал «бистабильный» переключатель в E. coli . Переключатель включается нагреванием культуры бактерий и выключается добавлением ИПТГ. В качестве репортера для своей системы они использовали зеленый флуоресцентный белок. [3] Второй, авторы Майкла Эловица и Станисласа Лейблера , показали, что три гена-репрессора могут быть связаны, образуя петлю отрицательной обратной связи, называемую репрессилятором , которая вызывает самоподдерживающиеся колебания уровней белка в E. coli. [4]
В настоящее время синтетические схемы являются растущей областью исследований в системной биологии , и каждый год публикуется все больше публикаций с подробным описанием синтетических биологических цепей. [5] Также был проявлен значительный интерес к поощрению образования и информационно-просветительской деятельности: Международный конкурс генно-инженерных машин. [6] управляет созданием и стандартизацией деталей BioBrick как средства, позволяющего студентам и старшим школьникам разрабатывать свои собственные синтетические биологические схемы.
Интерес и цели
[ редактировать ]Существуют как непосредственные, так и долгосрочные применения синтетических биологических цепей, включая различные применения в метаболической инженерии и синтетической биологии . Среди успешных примеров — фармацевтическое производство, [7] и производство топлива. [8] Однако методы, включающие прямое генетическое введение, по своей сути неэффективны без использования основных принципов синтетических клеточных цепей. Например, каждая из этих успешных систем использует метод индукции или выражения по принципу «все или ничего». Это биологическая схема, в которую вводится простой репрессор или промотор для облегчения создания продукта или ингибирования конкурирующего пути. Однако из-за ограниченного понимания сотовых сетей и естественных схем реализация более надежных схем с более точным управлением и обратной связью затруднена. В этом и заключается непосредственный интерес к синтетическим сотовым схемам.
Развитие понимания клеточных схем может привести к появлению новых интересных модификаций, например, клеток, способных реагировать на стимулы окружающей среды. Например, можно было бы разработать клетки, которые сигнализируют о токсичном окружении и реагируют, активируя пути, используемые для разрушения воспринимаемого токсина. [9] Чтобы разработать такую клетку, необходимо создать сложную синтетическую клеточную цепь, которая сможет соответствующим образом реагировать на данный стимул.
Учитывая, что синтетические клеточные цепи представляют собой форму контроля клеточной активности, можно предположить, что при полном понимании клеточных путей принцип «подключи и работай» [1] можно создать клетки с четко определенной генетической схемой. Широко распространено мнение, что если создать правильный набор деталей, [10] синтетические клетки могут быть созданы, используя только те пути, которые необходимы для выживания и размножения клеток. Из этой клетки, которую можно считать минимальной клеткой генома , можно добавить части из набора инструментов, чтобы создать четко определенный путь с соответствующей синтетической схемой для эффективной системы обратной связи. Благодаря базовому методу построения с нуля и предлагаемой базе данных сопоставленных частей схемы, методы, повторяющие те, которые используются для моделирования компьютерных или электронных схем, могут использоваться для перепроектирования ячеек и моделирования ячеек для облегчения устранения неполадок, прогнозирования поведения и производительности.
Примеры схем
[ редактировать ]Осцилляторы
[ редактировать ]- Репрессилятор
- Перестраиваемый синтетический генератор млекопитающих
- Бактериальный перестраиваемый синтетический генератор
- Связанный бактериальный осциллятор
- Глобально связанный бактериальный осциллятор
Еловиц и др. и Фунг и др. создали колебательные контуры, которые используют несколько механизмов саморегуляции для создания зависимых от времени колебаний экспрессии генных продуктов. [11] [12]
Бистабильные переключатели
[ редактировать ]- Тумблер
Гарднер и др. использовали взаимное подавление между двумя блоками управления, чтобы создать тумблер, способный управлять клетками бистабильным образом: временные стимулы приводят к постоянным реакциям. [3]
Регуляция генов является важной частью процессов развития. В процессе развития гены включаются и выключаются в разных тканях, изменения регуляторных механизмов могут приводить к генетическим переключениям в бистабильной системе, генные переключатели служат сайтами связывания регуляторных молекул. Это белки, которые активируют транскрипцию, когда они попадают на генный переключатель, и тем самым экспрессируют ген, который, как ожидается, будет работать как устройство памяти, позволяя выбирать и поддерживать решения о судьбе клетки. [13]
Тумблер, который работает с использованием двух взаимно ингибирующих генов, каждый промотор ингибируется репрессором, который транскрибируется противоположным промотором. Конструкция тумблера: Индуктор 1 инактивирует репрессор 1, что означает, что вырабатывается репрессор 2. Репрессор 2, в свою очередь, останавливает транскрипцию гена репрессора 1 и репортерного гена. [14]
Логические операторы
[ редактировать ]
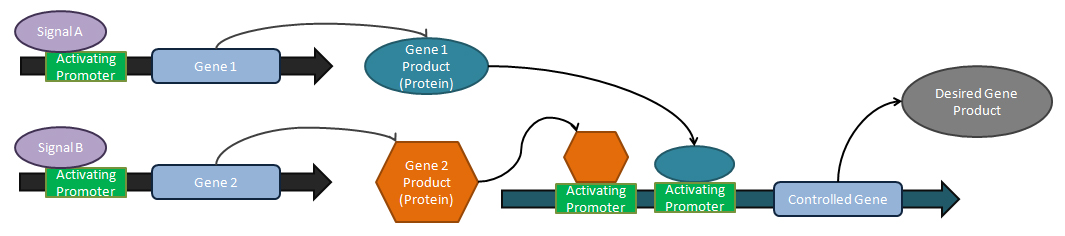

Аналоговые тюнеры
[ редактировать ]Используя отрицательную обратную связь и идентичные промоторы, генные цепи линеаризатора могут обеспечить однородную экспрессию генов, которая линейно зависит от концентрации внеклеточного химического индуктора. [17]
Контроллеры гетерогенности экспрессии генов
[ редактировать ]Синтетические генные цепи могут контролировать гетерогенность экспрессии генов, которую можно контролировать независимо от средства экспрессии генов. [18]
Другие инженерные системы
[ редактировать ]Инженерные системы являются результатом реализации комбинаций различных механизмов управления. Ограниченный механизм подсчета был реализован с помощью импульсно-управляемого генного каскада. [19] а применение логических элементов позволяет генетическое «программирование» клеток, как в исследовании Табора и др., Которые синтезировали светочувствительную программу обнаружения бактериальных границ. [20]
Схемотехника
[ редактировать ]
Недавние разработки в области искусственного синтеза генов и соответствующее усиление конкуренции внутри отрасли привели к значительному снижению цен и времени ожидания синтеза генов, а также помогли усовершенствовать методы, используемые при проектировании схем. [21] На данный момент схемотехника совершенствуется медленными темпами из-за недостаточной организации известных множественных взаимодействий генов и математических моделей. Эта проблема решается путем применения программного обеспечения для компьютерного проектирования (САПР) для обеспечения мультимедийного представления цепей с помощью изображений, текста и языка программирования, применяемого к биологическим цепям. [22] Некоторые из наиболее известных программ САПР включают GenoCAD, Clotho framework и j5. [23] [24] [25] GenoCAD использует грамматики, которые представляют собой «правила» с открытым исходным кодом или созданные пользователем, которые включают доступные гены и известные взаимодействия генов для клонирования организмов. Фреймворк Clotho использует стандартные правила Biobrick . [22]
Ссылки
[ редактировать ]- ^ Jump up to: а б с Кобаяши Х., Каэрн М., Араки М., Чанг К., Гарднер Т.С., Кантор Ч.Р., Коллинз Дж.Дж. (июнь 2004 г.). «Программируемые клетки: взаимодействие природных и инженерных генных сетей» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (22): 8414–8419. Бибкод : 2004PNAS..101.8414K . дои : 10.1073/pnas.0402940101 . ПМК 420408 . ПМИД 15159530 .
- ^ «Синтетическая биология: Часто задаваемые вопросы» . SyntheticBiology.org . Архивировано из оригинала 12 декабря 2002 года . Проверено 21 декабря 2011 г.
- ^ Jump up to: а б Гарднер Т.С., Кантор Ч.Р., Коллинз Дж.Дж. (январь 2000 г.). «Создание генетического тумблера в Escherichia coli». Природа . 403 (6767): 339–42. Бибкод : 2000Natur.403..339G . дои : 10.1038/35002131 . ПМИД 10659857 . S2CID 345059 .
- ^ Еловиц М.Б., Лейблер С. (январь 2000 г.). «Синтетическая колебательная сеть регуляторов транскрипции». Природа . 403 (6767): 335–338. Бибкод : 2000Natur.403..335E . дои : 10.1038/35002125 . ПМИД 10659856 . S2CID 41632754 .
- ^ Пурник П.Е., Вайс Р. (июнь 2009 г.). «Вторая волна синтетической биологии: от модулей к системам». Обзоры природы. Молекулярно-клеточная биология . 10 (6): 410–422. дои : 10.1038/nrm2698 . ПМИД 19461664 . S2CID 200495 .
- ^ Международные генно-инженерные машины (iGem) http://igem.org/Main_Page
- ^ Ро Д.К., Парадайз Э.М., Уэллет М., Фишер К.Дж., Ньюман К.Л., Ндунгу Дж.М. и др. (апрель 2006 г.). «Производство предшественника противомалярийного препарата артемизиновой кислоты в модифицированных дрожжах». Природа . 440 (7086): 940–943. Бибкод : 2006Natur.440..940R . дои : 10.1038/nature04640 . ПМИД 16612385 . S2CID 3199654 .
- ^ Фортман Дж.Л., Чхабра С., Мукхопадьяй А., Чоу Х., Ли Т.С., Стин Э., Кислинг Дж.Д. (июль 2008 г.). «Биотопливные альтернативы этанолу: прокачка микробной скважины» . Тенденции в биотехнологии . 26 (7): 375–381. дои : 10.1016/j.tibtech.2008.03.008 . ПМИД 18471913 . S2CID 205388761 .
- ^ Кислинг JD (январь 2008 г.). «Синтетическая биология для синтетической химии» . АКС Химическая биология . 3 (1): 64–76. дои : 10.1021/cb7002434 . ПМИД 18205292 .
- ^ Лакс Дж.Б., Ци Л, Уитакер В.Р., Аркин А.П. (декабрь 2008 г.). «На пути к масштабируемым семействам деталей для предсказуемого проектирования биологических схем». Современное мнение в микробиологии . 11 (6): 567–573. дои : 10.1016/j.mib.2008.10.002 . ПМИД 18983935 .
- ^ Еловиц М.Б., Лейблер С. (январь 2000 г.). «Синтетическая колебательная сеть регуляторов транскрипции». Природа . 403 (6767): 335–338. Бибкод : 2000Natur.403..335E . дои : 10.1038/35002125 . ПМИД 10659856 . S2CID 41632754 .
- ^ Фунг Э, Вонг В.В., Суен Дж.К., Балтер Т., Ли С.Г., Ляо Дж.К. (май 2005 г.). «Синтетический генно-метаболический осциллятор». Природа . 435 (7038): 118–122. Бибкод : 2005Natur.435..118F . дои : 10.1038/nature03508 . ПМИД 15875027 . S2CID 414371 .
- ^ Лугань Дж.Б., Соса Каррильо С., Кирх М., Кёлер А., Батт Г., Херсен П. (ноябрь 2017 г.). «Балансировка генетического тумблера с помощью управления с обратной связью в реальном времени и периодического воздействия» . Природные коммуникации . 8 (1): 1671. Бибкод : 2017NatCo...8.1671L . дои : 10.1038/s41467-017-01498-0 . ПМЦ 5693866 . ПМИД 29150615 .
- ^ Гарднер Т.С., Кантор Ч.Р., Коллинз Дж.Дж. (январь 2000 г.). «Создание генетического тумблера в Escherichia coli». Природа . 403 (6767): 339–342. Бибкод : 2000Natur.403..339G . дои : 10.1038/35002131 . ПМИД 10659857 . S2CID 345059 .
- ^ Jump up to: а б с Сильва-Роча Р., де Лоренцо В. (апрель 2008 г.). «Майнинг логических вентилей в прокариотических сетях регуляции транскрипции». Письма ФЭБС . 582 (8): 1237–1244. doi : 10.1016/j.febslet.2008.01.060 . ПМИД 18275855 . S2CID 45553956 .
- ^ Jump up to: а б с Бухлер Н.Е., Герланд У., Хва Т (апрель 2003 г.). «О схемах комбинаторной логики транскрипции» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (9): 5136–5141. Бибкод : 2003PNAS..100.5136B . дои : 10.1073/pnas.0930314100 . ПМК 404558 . ПМИД 12702751 .
- ^ Невожай Д., Адамс Р.М., Мерфи К.Ф., Йосич К., Балажши Г. (март 2009 г.). «Отрицательная ауторегуляция линеаризирует реакцию на дозу и подавляет гетерогенность экспрессии генов» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (13): 5123–5128. Бибкод : 2009PNAS..106.5123N . дои : 10.1073/pnas.0809901106 . ПМЦ 2654390 . ПМИД 19279212 .
- ^ Блейк В.Дж., Балажи Г., Кохански М.А., Исаакс Ф.Дж., Мерфи К.Ф., Куанг Ю. и др. (декабрь 2006 г.). «Фенотипические последствия транскрипционного шума, опосредованного промотором» . Молекулярная клетка . 24 (6): 853–865. doi : 10.1016/j.molcel.2006.11.003 . ПМИД 17189188 .
- ^ Фридланд А.Е., Лу Т.К., Ван X, Ши Д., Черч Дж., Коллинз Дж.Дж. (май 2009 г.). «Синтетические генные сети, которые имеют значение» . Наука . 324 (5931): 1199–1202. Бибкод : 2009Sci...324.1199F . дои : 10.1126/science.1172005 . ПМК 2690711 . ПМИД 19478183 .
- ^ Табор Дж.Дж., Салис Х.М., Симпсон З.Б., Шевалье А.А., Левская А., Маркотт Э.М. и др. (июнь 2009 г.). «Программа обнаружения синтетических генетических границ» . Клетка . 137 (7): 1272–1281. дои : 10.1016/j.cell.2009.04.048 . ПМЦ 2775486 . ПМИД 19563759 .
- ^ Ченг А.А., Лу Т.К. (1 января 2012 г.). «Синтетическая биология: новая инженерная дисциплина» . Ежегодный обзор биомедицинской инженерии . 14 (1): 155–178. doi : 10.1146/annurev-bioeng-071811-150118 . ПМИД 22577777 . S2CID 7319630 .
- ^ Jump up to: а б Блаби-Хаас CE, де Креси-Лагард V (апрель 2011 г.). «Извлечение экспериментальных данных с высокой пропускной способностью для связи гена и функции» . Тенденции в биотехнологии . 29 (4): 174–182. дои : 10.1016/j.tibtech.2011.01.001 . ПМЦ 3073767 . ПМИД 21310501 .
- ^ «GenoCAD: программное обеспечение САПР для синтетической биологии» . www.genocad.com . Архивировано из оригинала 5 февраля 2016 года . Проверено 21 октября 2015 г.
- ^ «Клото» . www.clothocad.org . Архивировано из оригинала 26 декабря 2014 года . Проверено 21 октября 2015 г.
- ^ «автоматическая сборка ДНК j5» . j5.jbei.org . Проверено 21 октября 2015 г.