Рибозим молотоголового типа

Молекула РНК рибозима «головка молотка»
Рибозим «головка молотка» представляет собой мотив РНК , который катализирует обратимые реакции расщепления и лигирования в определенном сайте молекулы РНК. Это одна из нескольких каталитических РНК ( рибозимов ), встречающихся в природе. Он служит модельной системой для исследования структуры и свойств РНК и используется для целевых экспериментов по расщеплению РНК, некоторые из которых предлагаются для терапевтического применения. Назван в честь сходства ранних схем вторичной структуры с акулой-молотом . [1] Рибозимы «головки молотка» первоначально были обнаружены в двух классах растительных вирусоподобных РНК: сателлитных РНК и вироидах . Они известны также в некоторых классах ретротранспозонов , в том числе в ретрозимах . [2] Мотив рибозима «головка молота» повсеместно встречается в линиях на древе жизни. [3] [4]
Реакции саморасщепления, впервые описанные в 1986 г. [5] [6] являются частью механизма репликации по вращающемуся кругу . Последовательность молоточков достаточна для саморасщепления. [7] и действует путем формирования консервативной трехмерной третичной структуры.
Катализ
[ редактировать ]В естественном состоянии мотив РНК «головка молотка» представляет собой одну цепь РНК. Хотя расщепление происходит в отсутствие белковых ферментов , РНК «головка молотка» сама по себе не является катализатором в своем естественном состоянии, поскольку она потребляется реакцией (т. е. осуществляет саморасщепление) и, следовательно, не может катализировать множественные обороты.
Трансдействующие конструкции «головки-молота» можно сконструировать таким образом, чтобы они состояли из двух взаимодействующих цепей РНК, причем одна цепь образует рибозим «головка-молот», который расщепляет другую цепь. [8] [ нужна ссылка ] Расщепляемая нить может поступать в избытке, и можно продемонстрировать множественный оборот, подчиняющийся кинетике Михаэлиса-Ментен . [8] [ нужна ссылка ] типичная для белково-ферментной кинетики . Такие конструкции обычно используются для экспериментов in vitro , и термин «РНК-молот» стал на практике синонимом более часто используемого « рибозима в форме головки-молота ».
Как показано , минимальная транс-действующая последовательность рибозима «головка молотка», которая является каталитически активной, состоит из трех спаренных оснований, фланкирующих центральное ядро из 15 консервативных (в основном инвариантных) нуклеотидов . Консервативные центральные основания, за некоторыми исключениями, необходимы для каталитической активности рибозимов. Такие рибозимные конструкции типа «головка молотка» демонстрируют in vitro скорость оборота ( ) около 1 молекулы/минуту и Km kcat порядка 10 наномолей.
Рибозим «головка молотка», возможно, является наиболее изученным рибозимом. Его небольшой размер, тщательно изученная химия расщепления, известная кристаллическая структура и его биологическая значимость делают рибозим «головка молотка» особенно хорошо подходящим для биохимических и биофизических исследований фундаментальной природы катализа РНК.
Рибозимы «головки молотка» могут играть важную роль в качестве терапевтических агентов; в качестве ферментов, которые адаптируют определенные последовательности РНК, в качестве биосенсоров , а также для приложений в функциональной геномике и открытии генов. [9]
Распространение видов
[ редактировать ]| Рибозим «молоток» HH9 | |
|---|---|
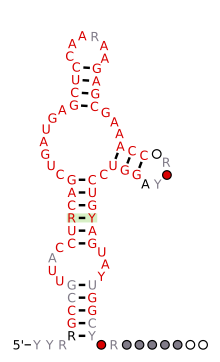 Предсказанная вторичная структура и консервативность последовательности рибозима HH9 обнаружены законсервированными от геномов ящерицы до человека | |
| Идентификаторы | |
| Символ | HH9 |
| Рфам | RF02275 |
| Другие данные | |
| РНК Тип | Джин ; рибозим |
| Домен(ы) | Эукариоты |
| ТАК | ТАК: 0000380 |
| PDB Структуры | ПДБе |
В 1986 году первые рибозимы «головки молотка» были обнаружены в РНК растительных патогенов, таких как вироиды и вирусные сателлиты. [5] [6] Год спустя рибозим «головка-молот» также был обнаружен в сателлитной ДНК геномов тритонов. [10] Новые примеры этого рибозима были затем обнаружены в геномах неродственных организмов, таких как шистосомы, [11] пещерные сверчки, [12] Арабидопсис Талиана [13] и несколько млекопитающих, таких как грызуны и утконосы. [14] В 2010 году было обнаружено, что рибозим «головка молотка» встречается в самых разных геномах бактерий и эукариот. [15] в том числе у человека. [16] Подобные сообщения подтвердили и расширили эти наблюдения. [17] [18] [19] открытие рибозима «головки молотка» как повсеместной каталитической РНК во всех жизненных царствах. [20]
Большинство эукариотических рибозимов типа «головка молотка» связаны с разновидностью коротких вкраплений ретроэлементов (SINE), называемых ретрозимами , которые выражаются в виде небольших кольцевых РНК. [21] Однако исключительную группу поразительно консервативных молотоголовок можно обнаружить в геномах всех амниот . [16] Эти рибозимы «головки молотка» (так называемые HH9 и HH10) встречаются в интронах нескольких специфических генов и указывают на сохранение биологической роли во время биосинтеза пре-мРНК. [22] В 2021 году было обнаружено, что новые геномы кольцевой РНК вируса гепатита D от различных животных кодируют рибозимы «головка молотка», аналогичные тем, которые присутствуют в вироидах растений и вирусных сателлитах. [23] Массовый биоинформационный поиск дельтавирусоподобных агентов по всему миру выявил сотни примеров геномов кольцевых РНК, несущих мотивы «головки молота», что указывает на то, что не только этот рибозим, но и небольшие кольцевые РНК с рибозимами являются повсеместными молекулами в биосфере. [24]
Химия катализа
[ редактировать ]Рибозим «головка молотка» осуществляет очень простую химическую реакцию, которая приводит к разрыву субстратной цепи РНК, особенно в С17, нуклеотиде сайта расщепления. Хотя расщепление РНК часто называют гидролизом , используемый механизм на самом деле не включает добавление воды . Скорее, реакция расщепления представляет собой просто изомеризацию , состоящую из перегруппировки связывающей фосфодиэфирной связи. С химической точки зрения это та же самая реакция, которая происходит при случайной деградации РНК, опосредованной основаниями , за исключением того, что она в высокой степени сайт-специфична, а ее скорость увеличивается в 10 000 раз и более.
Расщепление путем изомеризации фосфодиэфира
[ редактировать ]Реакция расщепления представляет собой фосфодиэфира реакцию изомеризации , которая инициируется отрывом 2'-гидроксильного протона рибозы в сайте расщепления от 2'-кислорода, который затем становится атакующим нуклеофилом в «линейном» или S N 2(P ) -подобная реакция, хотя неизвестно, удаляется ли этот протон до или во время химической стадии реакции расщепления типа «головка молота». (Реакция расщепления технически не является бимолекулярной , но ведет себя так же, как настоящая реакция S N 2(P); она претерпевает инверсию конфигурации после образования ассоциативного переходного состояния, состоящего из пентакоординированного оксифосфрана.) Атака и уход. Обе группы кислорода будут занимать два аксиальных положения в тригонально-бипирамидальной структуре переходного состояния, что требуется для S N механизма реакции, подобного 2 .
5'-продукт в результате этого механизма реакции расщепления обладает 2',3'-циклическим фосфатным концом, а 3'-продукт обладает 5'-ОН-концом, как и при неферментативном щелочном расщеплении РНК. Таким образом, реакция является обратимой, поскольку расщепляющийся фосфат остается фосфодиэфиром и, таким образом, может действовать как субстрат для лигирования, опосредованного РНК «головка молотка», без необходимости в АТФ или аналогичном экзогенном источнике энергии. [25] Реакция, катализируемая рибозимом «головки молотка», в отличие от формально идентичного неферментативного щелочного расщепления РНК, представляет собой высокоспецифичную последовательность реакцию расщепления с типичной скоростью оборота примерно 1 молекулы субстрата на молекулу фермента в минуту при pH 7,5 в 10 мМ. мг 2+ (так называемые «стандартные условия реакции» для минимальной последовательности РНК «головка молотка»), в зависимости от последовательности конкретной измеряемой конструкции рибозима «головка молотка». Это представляет собой увеличение скорости примерно в 10 000 раз по сравнению с неферментативным расщеплением РНК.
Потребность в ионах двухвалентных металлов
[ редактировать ]Первоначально считалось, что все рибозимы являются металлоферментами . Предполагалось, что ионы двухвалентных металлов типа Mg 2+ Считалось, что они выполняют две роли: способствуют правильному сворачиванию РНК и формируют каталитическое ядро. [26] Поскольку сама РНК не содержала достаточного разнообразия функциональных групп, считалось, что ионы металлов играют роль в активном центре, как это было известно о белках. Предполагаемый механизм действия иона Mg2+ заключался в следующем: депротонирование 2'-ОН-группы комплексом магния-аква-гидрокси, связанным кислородом про-R в сайте фосфатного расщепления, с последующей нуклеофильной атакой образовавшегося 2'-. алкаоксид на расщепленном фосфате, образуя промежуточный пентакоординатный фосфат. Последним этапом является уход уходящей 5'-группы с образованием 2',3'-циклического фосфата с инвертированной конфигурацией. [27]
Предполагалось, что гексагидратные магния ионы , находящиеся в равновесии с гидроксидом магния , могут играть роль общей кислоты и общего основания , аналогично тому, как два гистидина в РНКазе А. Была также предложена дополнительная роль ионов двухвалентных металлов в форме электростатической стабилизации переходного состояния .
Не металлофермент
[ редактировать ]В 1998 году было обнаружено [28] что рибозим «головка молотка», а также рибозим VS и рибозим «шпилька» не требуют присутствия ионов металлов для катализа, при условии, что присутствует достаточно высокая концентрация одновалентного катиона , позволяющая РНК сворачиваться. Это открытие показало, что сама РНК, вместо того, чтобы служить инертным, пассивным каркасом для связывания химически активных ионов двухвалентных металлов, вместо этого сама тесно участвует в химии катализа. Последние структурные результаты, описанные ниже, действительно подтверждают, что два инвариантных нуклеотида, G12 и G8, занимают соответствующую роль основного основания и основной кислоты в реакции расщепления «головки молотка».
Поэтому, строго говоря, рибозим «головка молотка» не может быть металлоферментом.
Первичная и вторичная структура
[ редактировать ]
Минимальный рибозим
[ редактировать ]Минимальная последовательность «головка молотка», необходимая для реакции саморасщепления, включает примерно 13 консервативных или инвариантных «ядерных» нуклеотидов, большинство из которых не участвуют в формировании канонических пар оснований Уотсона-Крика . Основная область окружена стеблями I, II и III, которые обычно состоят из канонических пар оснований Уотсона-Крика , но в остальном не ограничены в отношении последовательности. Скорость каталитического оборота минимальных рибозимов типа «головка молотка» составляет ~ 1/мин (обычно наблюдается диапазон от 0,1 до 10/мин, в зависимости от неконсервативных последовательностей и длин трех спиральных стеблей) в стандартных реакционных условиях с высоким содержанием Mg. 2+ (~10 мМ), pH 7,5 и 25 °C. Большая часть экспериментальных работ, проведенных с рибозимами «головки молотка», использовала минимальную конструкцию.
РНК типа I, типа II и типа III в виде головки молотка
[ редактировать ]| Рибозим «молоток» (тип I) | |
|---|---|
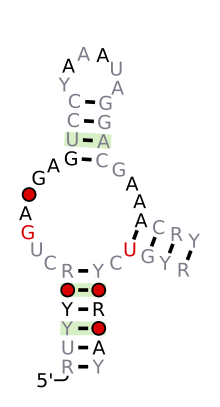 Предсказанная вторичная структура и сохранение последовательности Hammerhead_1 | |
| Идентификаторы | |
| Символ | Молот_1 |
| Рфам | RF00163 |
| Другие данные | |
| РНК Тип | Джин ; рибозим |
| Домен(ы) | Вироиды ; Эукариоты |
| ТАК | ТАК: 0000380 |
| PDB Структуры | ПДБе |
| Рибозим «молоток» (тип III) | |
|---|---|
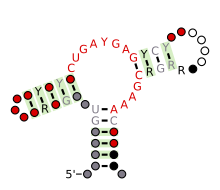 Прогнозируемая вторичная структура и консервативность последовательности рибозима Hammerhead (тип III) | |
| Идентификаторы | |
| Символ | Молот_3 |
| Рфам | RF00008 |
| Другие данные | |
| РНК Тип | Джин ; рибозим |
| Домен(ы) | Вироиды ; Эукариоты |
| ТАК | ТАК: 0000380 |
| PDB Структуры | ПДБе |
Структурно рибозим «головка молотка» состоит из трех спиралей, спаренных основаниями, разделенных короткими линкерами консервативных последовательностей. Эти спирали называются I, II и III. Рибозимы «головки-молота» можно разделить на три типа в зависимости от того, в какой спирали находятся 5'- и 3'-концы. Если 5'- и 3'-концы последовательности вносят вклад в стебель I, то это рибозим типа «головка-молот» I типа, в стебель II. представляет собой тип II, а стебель III - это рибозим типа III в форме головки молотка. Из трех возможных топологических типов тип I можно обнаружить в геномах прокариот, эукариот и РНК-патогенов растений, тогда как тип II описан только у прокариот. [18] [19] и тип III в основном встречаются у растений, фитопатогенов и прокариот. [20] [22]
Полноразмерный рибозим
[ редактировать ]Полноразмерный рибозим «головка молотка» состоит из дополнительных элементов последовательности в стеблях I и II, которые позволяют формировать дополнительные третичные контакты. Третичные взаимодействия стабилизируют активную конформацию рибозима, в результате чего скорость расщепления в 1000 раз превышает скорость расщепления соответствующих минимальных последовательностей в форме головки молотка. [29] [30]
Третичная структура
[ редактировать ]Минимальный
[ редактировать ]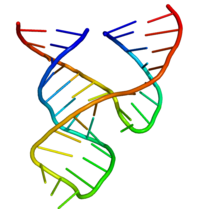
Минимальный рибозим «головка молотка» был тщательно изучен биохимиками и энзимологами, а также рентгеновскими кристаллографами, ЯМР-спектроскопистами и другими специалистами в области биофизических методов. Первая подробная трехмерная структурная информация о рибозиме «головка молотка» появилась в 1994 году в виде рентгеновской кристаллической структуры рибозима «головка молотка», связанного с аналогом субстрата ДНК, опубликованной в журнале Nature Плей, Флаэрти и Маккеем. [31] опубликовали минимальную структуру рибозима «головка молотка», состоящую из всех РНК Впоследствии в начале 1995 года Скотт, Финч и Клуг в журнале Cell . [32]
Минимальный рибозим «головка молотка» состоит из трех спиралей, спаренных основаниями, разделенных короткими линкерами консервативной последовательности, как показано на кристаллической структуре . [32] Эти спирали называются I, II и III. Консервативный поворот уридина соединяет спираль I со спиралью II и обычно содержит последовательность CUGA. Спирали II и III связаны последовательностью GAAA. Реакция расщепления происходит между спиралью III и I и обычно представляет собой C.
Структура полноразмерного рибозима показывает, что между петлей стебля II и стебля I существуют обширные взаимодействия. [33]
Подобные структуры наблюдаются и у других рибозимов, таких как рибозимы топорика .
Структура-функция
[ редактировать ]Несмотря на наблюдения катализа рибозимов «головка молотка» в кристалле минимальной последовательности «головка молота», в котором контакты упаковки кристаллической решетки по необходимости ограничивали глобальные положения дистальных концов всех трех фланкирующих спиральных стеблей, многие биохимические эксперименты были направлены на исследование взаимодействий в переходном состоянии и химия катализа оказалась несовместимой с кристаллическими структурами.
Например, инвариантные основные остатки G5, G8, G12 и C3 в минимальном рибозиме «головка молотка» оказались настолько хрупкими, что изменение даже одной экзоциклической функциональной группы на любом из этих нуклеотидов приводит к резкому снижению или отмене каталитической активности. , однако немногие из них, по-видимому, образуют водородные связи с участием граней Уотсона-Крика этих нуклеотидных оснований в любой из минимальных структур «головки молотка», за исключением взаимодействия G-5 в структуре продукта.
Особенно ярким и только недавно наблюдаемым примером были G8 и G12, которые были идентифицированы как возможные участники кислотно-основного катализа. Как только было продемонстрировано, что РНК «головка-молот» не требует ионов двухвалентных металлов для катализа, постепенно стало очевидно, что сама РНК, а не пассивно связанные ионы двухвалентных металлов, должна играть прямую химическую роль в любом кислотно-основном химическом процессе в рибозиме «головка-молот». активный сайт. Однако было совершенно неясно, как G12 и G8 могли добиться этого, учитывая исходные структуры минимального рибозима «головка молотка».
Другие опасения включали NOE между U4 и U7 расщепленного рибозима «головка молотка», который также наблюдался во время ЯМР- характеристики, который предполагал, что эти нуклеотидные основания должны сближаться друг с другом ближе, чем примерно на 6 Å, хотя близкое сближение U7 и U4, по-видимому, не вызывало опасений. возможно из кристаллической структуры. Наконец, как обсуждалось ранее, атакующий нуклеофил в исходных структурах, 2'-ОН C17, не находился в положении, поддающемся прямой атаке на соседний разрезаемый фосфат.
Возможно, наиболее тревожными были эксперименты, которые предположили, что А-9 и расщепляющиеся фосфаты должны находиться на расстоянии примерно 4 Å друг от друга в переходном состоянии, основанные на двойном замещении фосфортиоата и экспериментах по спасению ионов мягких металлов; Расстояние между этими фосфатами в минимальной кристаллической структуре «головка молота» составляло около 18 Å, при этом не было четкого механизма близкого сближения, если спирали A-формы Stem II и Stem I рассматривать как твердые тела. В совокупности эти результаты, по-видимому, позволяют предположить, что для достижения переходного состояния внутри минимальной структуры рибозима «головка молотка» должно было произойти довольно крупномасштабное конформационное изменение.
По этим причинам две серии экспериментов (биохимические и кристаллографические) оказались не только противоречащими друг другу, но и совершенно и безнадежно несовместимыми, порождая значительное количество разногласий в этой области. Никаких убедительных доказательств отклонения того или иного набора экспериментальных результатов так и не было получено, хотя в пользу каждого из них было высказано множество утверждений об обратном.
Полная длина
[ редактировать ]
В 2006 году была получена кристаллическая структура полноразмерного рибозима «головка молотка» с разрешением 2,2 Å. Эта новая структура (показанная справа), по-видимому, устраняет наиболее тревожные из предыдущих несоответствий. В частности, C17 теперь расположен для прямой атаки, а инвариантные остатки C3, G5, G8 и G12, по-видимому, участвуют в жизненно важных взаимодействиях, имеющих отношение к катализу. A9 и расщепляющиеся Более того, обнаружено, что фосфаты находятся на расстоянии 4,3 Å друг от друга, что согласуется с идеей о том, что при модификации эти фосфаты могут связывать один ион тиофильного металла. Структура также показывает, как два инвариантных остатка, G-12 и G-8, расположены внутри активного центра в соответствии с их ранее предполагаемой ролью в кислотно-основном катализе. G12 находится на расстоянии водородной связи с 2'-O C17, нуклеофила в реакции расщепления, а рибоза G8 образует водородные связи с уходящей группой 5'-O. (см. ниже), а нуклеотидное основание G8 образует пару Уотсона-Крика с инвариантом C3. Такое расположение позволяет предположить, что G12 является основным основанием в реакции расщепления и что G8 может действовать как обычная кислота, что согласуется с предыдущими биохимическими наблюдениями. Водородные связи G5 с кислородом фуранозы C17 помогают подготовить его к атаке в линию. U4 и U7, как следствие образования пары оснований между G8 и C3, теперь расположены так, что NOE между их основаниями легко объяснить.
Таким образом, кристаллическая структура полноразмерного рибозима с головкой-молотом ясно решает все основные проблемы, которые казались несовместимыми с предыдущими кристаллическими структурами минимального рибозима с головкой-молотом.
Структура и катализ
[ редактировать ]
Третичные взаимодействия в полноразмерном рибозиме «головка молотка» стабилизируют то, что явно является активной конформацией. Нуклеофил, 2'-кислород нуклеотида сайта расщепления, C17, почти идеально ориентирован для прямой атаки (реакция SN 2 (P)). G12 расположен на расстоянии водородной связи этого нуклеофила и, следовательно, сможет оторвать протон от 2'-кислорода, если сам G12 станет депротонированным. 2'-OH G8 образует водородную связь с кислородом 5'-уходящей группы и, следовательно, потенциально может поставлять протон, поскольку отрицательный заряд накапливается на 5'-кислороде рибозы A1.1.
Наиболее вероятное объяснение состоит в том, что G12 в депротонированной форме является основным основанием, а рибоза G8 представляет собой общую кислоту. Кажущаяся кинетическая pKa [ нужны разъяснения ] рибозима «головки молотка» составляет 8,5, тогда как рКа гуанозина составляет около 9,5. Возможно, что в молоточковом каталитическом ядре рКа G12 изменяется от 9,5 до 8,5; эта гипотеза в настоящее время является предметом интенсивного исследования.

Если инвариант G8 заменяется на C8, молоточковый катализ отменяется. Однако двойной мутант G8C + C3G, который сохраняет пару оснований G8-C3, обнаруженную в полноразмерной головке молотка, восстанавливает большую часть каталитической активности. Также было обнаружено, что 2'-ОН G8 важен для катализа; замена G8 на дезоксиG8 значительно снижает скорость катализа, что позволяет предположить, что 2'-ОН действительно имеет решающее значение для каталитического механизма.
Близкое сближение А9 и расщепляющихся фосфатов требует наличия высокой концентрации положительного заряда. Вероятно, это является источником наблюдения о том, что ионы двухвалентных металлов необходимы при низкой ионной силе, но без них можно обойтись при более высоких концентрациях одновалентных катионов.
Таким образом, реакция, вероятно, включает отрыв 2'-протона от C17 с последующей нуклеофильной атакой соседнего фосфата. Когда связь между расщепляющимся фосфором и уходящей группой 5'-O начинает разрываться, из рибозы G8 поступает протон, который затем, вероятно, репротонирует за счет наблюдаемой молекулы воды, образующей с ней водородную связь в кристаллической структуре. .
Терапевтическое применение
[ редактировать ]Модифицированные рибозимы «головки молотка» тестируются в качестве терапевтических средств. [34] Синтетические РНК, содержащие последовательности, комплементарные мутантной мРНК SOD1, и последовательности, необходимые для формирования каталитической структуры «головки молотка», изучаются в качестве возможной терапии бокового амиотрофического склероза . Также ведется работа по выяснению того, можно ли их использовать для создания ВИЧ линий Т-клеток, устойчивых к . Было показано, что модифицированные аденовирусы с рибозимом «головка молотка» эффективны в лечении рака как in vitro, так и in vivo. [35]
Терапевтическое использование транс -расщепляющих рибозимов «головки молотка» серьезно затруднено из-за их низкого уровня активности in vivo . Истинный каталитический потенциал транс -расщепляющих рибозимов «головки молотка» может быть восстановлен in vivo , и терапевтические производные, вероятно, будут дополнять другие терапевтические стратегии гибридизации нуклеиновых кислот. Уже существуют рибозимы типа «молоток», близкие к клиническому применению. [9]
Ссылки
[ редактировать ]- ^ Форстер AC, Саймонс Р.Х. (1987). «Саморасщепление плюсовых и минусовых РНК вирусоида и структурная модель активных центров». Клетка . 49 (2): 211–220. дои : 10.1016/0092-8674(87)90562-9 . ПМИД 2436805 . S2CID 33415709 .
- ^ Сервера, Амелия; Урбина, Денис; де ла Пенья, Маркос (2016). «Ретрозимы представляют собой уникальное семейство неавтономных ретротранспозонов с рибозимами в виде головки молотка, которые размножаются в растениях посредством кольцевых РНК» . Геномная биология . 17 (1): 135. дои : 10.1186/s13059-016-1002-4 . ISSN 1474-760X . ПМЦ 4918200 . ПМИД 27339130 .
- ^ де ла Пенья, Маркос; Гарсиа-Роблес, Инмакулада (2010). «Повсеместное присутствие рибозимного мотива «головка молотка» на древе жизни» . РНК . 16 (10): 1943–1950. дои : 10.1261/rna.2130310 . ISSN 1355-8382 . ПМК 2941103 . ПМИД 20705646 .
- ^ Хамманн, Кристиан; Луптак, Андрей; Перро, Джонатан; де ла Пенья, Маркос (2012). «Вездесущий рибозим-молот» . РНК . 18 (5): 871–885. дои : 10.1261/rna.031401.111 . ISSN 1355-8382 . ПМЦ 3334697 . ПМИД 22454536 .
- ^ Перейти обратно: а б Проди Г.А., Бакос Дж.Т., Бузаян Дж.М., Шнайдер И.Р., Брюнинг Г. (1986). «Автолитический процессинг димерной сателлитной РНК вируса растений». Наука . 231 (4745): 1577–1580. Бибкод : 1986Sci...231.1577P . дои : 10.1126/science.231.4745.1577 . ПМИД 17833317 . S2CID 21563490 .
- ^ Перейти обратно: а б Хатчинс С.Дж., Ратьен П.Д., Форстер А.К., Саймонс Р.Х. (1986). «Саморасщепление плюсовых и минусовых РНК-транскриптов вироида солнечной пятнистости авокадо» . Нуклеиновые кислоты Рез . 14 (9): 3627–3640. дои : 10.1093/нар/14.9.3627 . ПМК 339804 . ПМИД 3714492 .
- ^ Форстер AC, Саймонс Р.Х. (1987). «Саморасщепление вирусоидной РНК осуществляется предполагаемым активным центром из 55 нуклеотидов». Клетка . 50 (1): 9–16. дои : 10.1016/0092-8674(87)90657-X . ПМИД 3594567 . S2CID 7231363 .
- ^ Перейти обратно: а б Усман, Нассим; Бейгельман, Леонид; МакСвигген, Джеймс А. (1 августа 1996 г.). «Инженерия рибозимов-молотов». Современное мнение в области структурной биологии . 6 (4): 527–533. дои : 10.1016/S0959-440X(96)80119-9 . ПМИД 8794164 .
- ^ Перейти обратно: а б Хин Дж., Вайнберг М.С. (2008). «Возвращение к рибозиму-молоту: новые биологические идеи для разработки терапевтических агентов и приложений обратной геномики» . РНК и регуляция экспрессии генов: скрытый уровень сложности . Кайстер Академик Пресс. ISBN 978-1-904455-25-7 .
- ^ Эпштейн Л.М., Галл Дж.Г. (1987). «Саморасщепляющиеся транскрипты сателлитной ДНК тритона». Клетка . 48 (3): 535–543. дои : 10.1016/0092-8674(87)90204-2 . ПМИД 2433049 . S2CID 24110291 .
- ^ Фербейр Дж., Смит Дж.М., Седергрен Р. (1998). «Сателлитная ДНК шистосом кодирует активные рибозимы типа «молоток» . Мол. Клетка. Биол . 18 (7): 3880–3888. дои : 10.1128/MCB.18.7.3880 . ПМК 108972 . ПМИД 9632772 .
- ^ Рохас А.А., Васкес-Телло А, Фербейр Г, Венанцетти Ф, Бахманн Л, Пакуин Б, Сбордони В, Седергрен Р (2000). «Опосредованная молотком обработка спутниковых транскриптов семейства pDo500 пещерных сверчков Dolichopoda» . Нуклеиновые кислоты Рез . 28 (20): 4037–4043. дои : 10.1093/нар/28.20.4037 . ПМЦ 110794 . ПМИД 11024185 .
- ^ Пшибильски Р., Греф С., Лескут А., Неллен В., Вестхоф Е., Стегер Г., Хамманн С. (2005). «Функциональные рибозимы «головки молотка», естественно закодированные в геноме Arabidopsis thaliana» . Растительная клетка . 17 (7): 1877–1885. дои : 10.1105/tpc.105.032730 . ПМЦ 1167538 . ПМИД 15937227 .
- ^ Мартик М., Хоран Л.Х., Ноллер Х.Ф., Скотт В.Г. (2008). «Прерывистый рибозим в форме головы молотка, встроенный в информационную РНК млекопитающих» . Природа . 454 (7206): 899–902. Бибкод : 2008Natur.454..899M . дои : 10.1038/nature07117 . ПМК 2612532 . ПМИД 18615019 .
- ^ Де ла Пенья М., Гарсиа-Роблес I (2010). «Повсеместное присутствие рибозимного мотива «головка молотка» на древе жизни» . РНК . 16 (10): 1943–1950. дои : 10.1261/rna.2130310 . ПМК 2941103 . ПМИД 20705646 .
- ^ Перейти обратно: а б Де ла Пенья М., Гарсиа-Роблес I (2010). «Интронные рибозимы типа «головка молотка» ультраконсервативны в геноме человека» . Представитель ЭМБО . 11 (9): 711–716. дои : 10.1038/embor.2010.100 . ПМЦ 2933863 . ПМИД 20651741 .
- ^ Зеехафер С., Калвейт А., Стегер Г., Греф С., Хамманн С. (2011). «От альпаки до рыбок данио: рибозимы-молоты, куда ни глянь» . РНК . 17 (1): 21–26. дои : 10.1261/rna.2429911 . ПМК 3004062 . ПМИД 21081661 .
- ^ Перейти обратно: а б Хименес Р.М., Делварт Э., Луптак А. (2011). «Поиск на основе структуры обнаруживает рибозимы типа «молоток» в микробиоме человека» . J Биол Хим . 286 (10): 7737–7743. дои : 10.1074/jbc.C110.209288 . ПМК 3048661 . ПМИД 21257745 .
- ^ Перейти обратно: а б Перро Дж., Вайнберг З., Рот А., Попеску О., Шартран П., Фербейр Г., Брейкер Р.Р. (май 2011 г.). «Идентификация рибозимов «головки молотка» во всех сферах жизни выявляет новые структурные вариации» . PLOS Вычислительная биология . 7 (5): e1002031. Бибкод : 2011PLSCB...7E2031P . дои : 10.1371/journal.pcbi.1002031 . ПМК 3088659 . ПМИД 21573207 .
- ^ Перейти обратно: а б Хамманн С., Луптак А., Перро Дж., де ла Пенья М. (2012). «Вездесущий рибозим-молот» . РНК . 18 (5): 871–885. дои : 10.1261/rna.031401.111 . ПМЦ 3334697 . ПМИД 22454536 .
- ^ Сервера А, де ла Пенья М (2020). «Маленькие циркРНК с саморасщепляющимися рибозимами высоко экспрессируются в разнообразных транскриптомах многоклеточных животных» . Исследования нуклеиновых кислот . 48 (9): (9): 5054–5064. дои : 10.1093/nar/gkaa187 . ПМК 7229834 . ПМИД 32198887 .
- ^ Перейти обратно: а б Гарсиа-Роблес I, Санчес-Наварро Х, де ла Пенья М (2012). «Интронные рибозимы типа «молоток» в биогенезе мРНК». Биол хим . 393 (11): 1317–1326. дои : 10.1515/hsz-2012-0223 . hdl : 10251/34564 . ПМИД 23109545 . S2CID 16402212 .
- ^ де ла Пенья М., Сеприан Р., Кейси Дж., Сервера А. (2021). «Кольцевые РНК, подобные вирусу гепатита дельта, от различных многоклеточных животных, кодируют консервативные рибозимы «головки молотка»» . Эволюция вирусов . 7 (1): veab016. дои : 10.1093/ve/veab016 . ПМЦ 7936874 . ПМИД 33708415 .
- ^ Эдгар Р.К., Тейлор Дж., Лин В., Альтман Т., Барбера П., Мелешко Д., Лор Д., Новаковский Г., Бухфинк Б., Аль-Шайеб Б., Банфилд Дж.Ф., де ла Пенья М., Коробейников А., Чихи Р., Бабаян А. (2022) ). «Выравнивание последовательностей в масштабе петаоснований катализирует открытие вирусов» . Природа . 602 (7895): 142–147. Бибкод : 2022Natur.602..142E . дои : 10.1038/s41586-021-04332-2 . ПМИД 35082445 . S2CID 246297430 .
- ^ Кэнни, Мэриленд, Джакер Ф.М., Парди А. (2007). «Эффективное лигирование рибозима головки молотка шистосомы» . Биохимия . 46 (12): 3826–3834. дои : 10.1021/bi062077r . ПМК 3203546 . ПМИД 17319693 .
- ^ Лотт, Уильям Б.; Понтиус, Брайан В.; фон Хиппель, Питер Х. (20 января 1998 г.). «Механизм ионов двух металлов действует при расщеплении субстрата РНК, опосредованном рибозимом «головка молота»» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (2): 542–547. Бибкод : 1998PNAS...95..542L . дои : 10.1073/pnas.95.2.542 . ISSN 0027-8424 . ЧВК 18456 . ПМИД 9435228 .
- ^ Бирих, КР; Хитон, Пенсильвания; Экстайн, Ф. (1 апреля 1997 г.). «Структура, функции и применение рибозима «головка молотка»» . Европейский журнал биохимии . 245 (1): 1–16. дои : 10.1111/j.1432-1033.1997.t01-3-00001.x . ISSN 0014-2956 . ПМИД 9128718 .
- ^ Дж. Б. Мюррей; А.А. Сейхан; Н. Г. Уолтер; Дж. М. Берк; У.Г. Скотт (1998). «Рибозимы «головка молота», «шпилька» и VS каталитически эффективны только в отношении одновалентных катионов» (PDF) . Хим Биол . 5 (10): 587–595. дои : 10.1016/S1074-5521(98)90116-8 . ПМИД 9818150 . S2CID 17025877 .
- ^ Хворова А, Лескут А, Вестхоф Э, Джаясена СД (2003). «Элементы последовательности за пределами каталитического ядра рибозима «головка молотка» обеспечивают внутриклеточную активность». Структурная и молекулярная биология природы . 10 (9): 708–712. дои : 10.1038/nsb959 . ПМИД 12881719 . S2CID 52859654 .
- ^ Де ла Пенья М; Гаго С.; Флорес Р. (2003). «Периферические области природных рибозимов «головки молотка» значительно повышают их активность саморасщепления» . ЭМБО Дж . 22 (20): 5561–5570. дои : 10.1093/emboj/cdg530 . ПМК 213784 . ПМИД 14532128 .
- ^ Плей Х.В., Флаэрти К.М., Маккей Д.Б. (1994). «Трехмерная структура рибозима «головка молотка». Природа . 372 (6501): 68–74. Бибкод : 1994Natur.372...68P . дои : 10.1038/372068a0 . ПМИД 7969422 . S2CID 4333072 .
- ^ Перейти обратно: а б Скотт В.Г., Финч Дж.Т., Клуг А. (1995). «Кристаллическая структура рибозима с головкой молотка, состоящего из всех РНК: предлагаемый механизм каталитического расщепления РНК» (PDF) . Клетка . 81 (7): 991–1002. дои : 10.1016/S0092-8674(05)80004-2 . ПМИД 7541315 . S2CID 6965437 .
- ^ Мартик М., Скотт В.Г. (2006). «Третичные контакты, удаленные от активного центра, запускают рибозим для катализа» . Клетка . 126 (2): 309–320. дои : 10.1016/j.cell.2006.06.036 . ПМК 4447102 . ПМИД 16859740 .
- ^ Читти Л., Райнальди Дж. (2005). «Синтетические рибозимы «головки молотка» как терапевтические инструменты для контроля генов болезней». Современная генная терапия . 5 (1): 11–24. CiteSeerX 10.1.1.333.3872 . дои : 10.2174/1566523052997541 . ПМИД 15638708 .
- ^ Фей Ц, Чжан Х, Фу Л, Дай Икс, Гао Б, Ни М, Ге С, Ли Дж, Дин Икс, Кэ Ю, Яо Х, Чжу Дж (июнь 2008 г.). «Экспериментальная генная терапия рака с помощью множественных рибозимов «головка молотка» против выживаемости». Acta Biochimica et Biophysica Sinica . 40 (6): 466–77. дои : 10.1111/j.1745-7270.2008.00430.x . ПМИД 18535745 . S2CID 31532433 .
Внешние ссылки
[ редактировать ]- Страницы лаборатории Билла Скотта, посвященные рибозиму «головка-молот». Архивировано 29 сентября 2011 г. в Wayback Machine.
- Страница лаборатории Маркоса де ла Пенья, посвященная рибозиму «головка молота».
- Страница для Hammerhead типа I на Rfam
- Страница для Hammerhead типа II на Rfam
- Страница для Hammerhead типа III на Rfam
- Страница Hammerhead HH9 на Rfam
- Страница Hammerhead HH10 на Rfam