Дынеголовый кит
| Дынеголовый кит | |
|---|---|

| |

| |
| Размер по сравнению со средним человеком | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Парнокопытные |
| Инфрапорядок: | китообразные |
| Семья: | Дельфиниды |
| Род: | Пепоноцефала Нишиваки и Норрис, 1966 г. |
| Разновидность: | П. Электра
|
| Биномиальное имя | |
| Пепоноцефала электра ( Грей , 1846)
| |
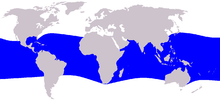
| |
| Ареал дынноголового кита | |
| Синонимы | |
|
Пепоноцефал электрус | |

Дынеголовый кит ( Peponocephalalectra ), также известный менее широко как электра , маленькая косатка или многозубая черная рыба , — зубатый кит семейства океанических дельфинов дельфин (Delphinidae). Общее название происходит от формы головы. Дынеголовые киты широко распространены в глубоких тропических и субтропических водах по всему миру, но в море их редко можно встретить. Они встречаются вблизи берега, в основном вокруг океанических островов, таких как Гавайи, Французская Полинезия и Филиппины.
Таксономия
[ редактировать ]Дынеголовый кит — единственный представитель рода Peponocephala . Впервые зарегистрированный по образцу, собранному на Гавайях в 1841 году, этот вид был первоначально описан как член семейства дельфинов и назван в 1846 году как Lagenorhynchus Electra. Джоном Эдвардом Греем Позже было установлено, что дынеголовый кит достаточно отличается от других Lagenorhynchus видов . быть отнесен к собственному роду. [ 3 ] Члены подсемейства Globicephalinae , дынеголовые киты тесно связаны с длинноплавниковыми и короткоплавниковыми гриндами ( Globicephala melas и G. macrorhynchus соответственно) и карликовой косаткой ( Feresa attenuata ). [ 4 ] [ 5 ] В совокупности эти виды (включая косаток Orcinus orca и ложных косаток Pseudorca crassidens ) известны под общим названием «черная рыба». Дынеголовые киты — один из самых мелких видов китообразных (после карликовых косаток), в общем названии которых есть слово «кит».
Описание
[ редактировать ]Дынеголовые киты имеют крепкое дельфиноподобное тело, сужающуюся коническую голову (треугольная форма головы, если смотреть сверху) без заметного клюва и относительно высокий серповидный (серповидный) спинной плавник , расположенный около середины назад. Окраска тела от угольно-серой до темно-серой. Темная «маска» лица простирается от глаза до передней части дыни, а у более крупных животных губы беловатые. У дынеголовых китов есть тёмная спинная накидка, которая начинается узко в передней части головы и опускается под крутым углом под спинной плавник. Граница между более темным мысом и окраской боков часто бывает слабой или размытой. И маска, и спинной плащ часто видны только при хорошем освещении. По сравнению с самками взрослые самцы имеют более округлую голову, более длинные ласты, более высокие спинные плавники, более широкие хвостовые плавники, а некоторые имеют выраженный брюшной киль позади анального отверстия. [ 6 ] [ 7 ] [ 8 ]
Дынеголовые киты вырастают до 2,75 м (9,0 футов) в длину и весят до 225 кг (496 фунтов), причем взрослые самцы немного крупнее самок. [ 7 ] [ 9 ] Длина при рождении составляет примерно 1 м (3,3 фута). [ 10 ] [ 11 ] Дынеголовые киты физически созревают в 13–15 лет и живут до 45 лет. [ 7 ]
Похожие виды
[ редактировать ]В море дынеголовых китов можно спутать с карликовыми косатками , которые очень похожи по внешнему виду и имеют почти идентичную среду обитания и ареал. [ 12 ] Форма головы, ласт и спинного мыса может быть полезным диагностическим признаком. У дынеголовых китов ласты с заостренными кончиками, тогда как у карликовых косаток кончики ласт закруглены, и, если смотреть сверху, форма головы более треугольная, чем закругленная голова карликовой косатки. [ 8 ] Спинной мыс дынеголовых китов закруглен и опускается намного ниже спинного плавника, чем у карликовых косаток, который опускается под относительно пологим углом и более резко разграничен по цвету между темным мысом и более светлыми боками. [ 13 ] Хотя у обоих видов белый цвет вокруг рта, у взрослых карликовых косаток он может распространяться на лицо. [ 14 ]
Многие из этих признаков трудно различить в сложных условиях моря и/или освещения, а поведение часто является полезным средством идентификации. Карликовые косатки двигаются медленнее (хотя дынеголовые киты часто неподвижно лежат на поверхности) и обычно встречаются гораздо меньшими группами, чем дынеголовые киты. Большие группы (более 100 особей) чаще всего представляют собой дынеголовые киты. [ 12 ] [ 9 ] Издалека дынеголовых китов также можно спутать с ложными косатками ( Pseudorca crassidens ), но у ложных китов гораздо больший размер — 5–6 м (16–20 футов) длина взрослой особи, длинное стройное тело и относительно меньший спинной плавник. косаток следует отличать их от дынеголовых китов.
Особей, выброшенных на мель или трупных особей, можно легко идентифицировать по количеству зубов: у дынеголовых китов имеется 20–25 пар тонких зубов (более похожих на зубы более мелких дельфинов, чем у других черных рыб) как в верхней, так и в нижней челюстях по сравнению с 8–25 парами тонких зубов (более похожих на зубы более мелких дельфинов, чем у других черных рыб). У карликовых косаток 13 пар крепких зубов как на верхней, так и на нижней челюсти. [ 13 ]
Географический ареал и распространение
[ редактировать ]
Дынеголовые киты встречаются в глубоких тропических и субтропических океанических водах, между 40° с.ш. и 35° ю.ш. [ 9 ] Хотя этот вид считается прибрежным пелагическим видом, в некоторых регионах существуют островные популяции (например, на Гавайях ), и их можно встретить недалеко от берега, связанного с океаническими островами и архипелагами, такими как Пальмира и Филиппины . [ 15 ] Наблюдения или выбросы на берег в крайних пределах их ареала, вероятно, связаны с распространением теплых течений. [ 9 ] Дынеголовые киты не являются мигрирующими.
Поведение
[ редактировать ]Собирательство
[ редактировать ]Дынеголовые киты питаются в основном пелагическими и мезопелагическими кальмарами и мелкой рыбой. [ 16 ] [ 17 ] Сообщалось также о ракообразных (креветках) из содержимого желудка. [ 8 ] [ 16 ] Мезопелагические виды кальмаров и рыб демонстрируют вертикальную миграцию , обитая на больших глубинах в течение дня и перемещаясь на сотни метров на более мелкие глубины после наступления сумерек, чтобы питаться планктоном. Дынеголовые киты кормятся ночью, когда их добыча находится в пределах верхних 400 м (1300 футов) водной толщи. [ 16 ] На Гавайских островах особи, оснащенные спутниковыми метками, передающими глубину, совершали ночные погружения в поисках пищи, средняя дальность которых составляла 219,5–247,5 м (720–812 футов), при этом максимальная зарегистрированная глубина погружения составляла 471,5 м (1547 футов). [ 16 ]
Социальные
[ редактировать ]Дынеголовые киты являются очень социальным видом и обычно путешествуют большими группами по 100–500 особей, время от времени встречаются стада численностью до 1000–2000 особей. [ 13 ] Большие стада, по-видимому, состоят из более мелких подгрупп, которые объединяются в более крупные группы. [ 18 ] Данные массовых выбрасываний на берег в Японии позволяют предположить, что дынеголовые киты могут иметь матрилинейную социальную структуру (т.е. связаны через родственников/группы самок, организованные вокруг старшей самки и их родственников); предвзятое соотношение полов (большее количество самок) в выброшенных на берег группах позволяет предположить, что зрелые самцы могут перемещаться между группами. [ 11 ] В то время как дынеголовые киты объединяются в большие группы (общая черта среди океанических дельфинов, в отличие от более мелких групп других видов черной рыбы), их социальная структура может быть более стабильной и промежуточной между более крупными черными рыбами (гринда, косатка и ложные косатки) и более мелкие океанические дельфины. [ 11 ] Однако генетические исследования дынеголовых китов в бассейнах Тихого, Индийского и Атлантического океанов позволяют предположить, что между популяциями существует относительно высокий уровень связи (межбридинга). [ 19 ] Это указывает на то, что дынеголовые киты могут не проявлять сильной преданности своей натальной группе (группе, в которой особь родилась) и что скорость перемещения особей между популяциями выше, чем у других видов черной рыбы. [ 19 ] Большие размеры групп могут усилить конкуренцию за ресурсы добычи, что потребует больших ареалов обитания и широкомасштабных перемещений в поисках пищи. [ 19 ]
Наблюдения за суточной активностью дынеголовых китов вблизи океанических островов позволяют предположить, что они проводят утро, отдыхая или копаясь в приповерхностных водах, после ночного кормления. [ 15 ] Поверхностная активность (например, хлопанье хвостом и шпионские прыжки) и вокализация, связанная с общением (коммуникационные свистки, а не эхолокационные щелчки, используемые для поиска пищи), увеличиваются во второй половине дня. [ 15 ] [ 20 ] Ежедневный образец поведения, наблюдаемый в островных популяциях, в сочетании с более крупными размерами групп дынеголовых китов (по сравнению с тем, что обычно наблюдается у других видов черной рыбы), больше похож на деления-слияния социальную структуру дельфинов-спиннеров ( Stenella longirostris ). ). [ 15 ] [ 19 ] Эти поведенческие черты могут быть связаны с избеганием хищников (более крупные группы обеспечивают некоторую защиту от крупных океанических акул) и привычками добывания пищи (оба вида являются ночными хищниками, которые охотятся на предсказуемых, относительно многочисленных мезопелагических кальмаров и рыб, которые совершают частые вертикальные миграции из глубоководных в поверхность).
Дынеголовые киты часто общаются с дельфинами Фрейзера ( Lagenodelphis hosei ), а также встречаются, хотя и реже, в смешанных стадах с другими видами дельфинов, такими как дельфины-спиннеры, обыкновенные афалины ( Tursiops truncatus ), зубастые дельфины ( Steno bredanensis). ), короткоплавниковые гринды и пантропические пятнистые дельфины ( Stenella attenuata ). [ 21 ] [ 22 ]
Уникальный случай межвидового усыновления между (предположительно) осиротевшим детёнышем дынноголового кита и матерью обыкновенной афалины был зафиксирован во Французской Полинезии. Впервые детеныш был замечен в 2014 году в возрасте менее одного месяца, когда он плавал вместе с самкой афалины и ее собственным биологическим потомством. [ 23 ] Детеныш дынноголового кита наблюдался вскармливающим грудью самку афалины, и до 2018 года его неоднократно видели вместе с приемной/приемной матерью. [ 23 ]
В августе 2017 года у острова Кауаи, Гавайи, был замечен гибрид дынноголового кита и грубозубого дельфина, путешествующий с дынеголовым китом среди группы зубастых дельфинов. [ 24 ] Гибрид внешне напоминал дынеголового кита, но более пристальное наблюдение показало, что он имел черты обоих видов, а также некоторые промежуточные черты между этими двумя видами, особенно в форме головы. Генетическое тестирование образца биопсии кожи подтвердило, что этот человек представлял собой гибрид самки дынноголового кита и самца грубозубого дельфина. [ 24 ]
Хищники
[ редактировать ]На дынеголовых китов могут охотиться крупные акулы и косатки. [ 25 ] [ 26 ] Шрамы и раны от несмертельных укусов акул-резаков ( Isitius brasiliensis ) наблюдались у животных, находящихся на свободном выгуле и на мели. [ 6 ] [ 25 ]
Разведение
[ редактировать ]О репродуктивном поведении дынеголовых китов известно мало. Наибольшую информацию дает анализ крупных групп, выброшенных на берег в водах Японии, где половая зрелость самок достигается в возрасте 7 лет. [ 6 ] Самки рожают одного теленка каждые 3–4 года после беременности продолжительностью примерно 12 месяцев. [ 10 ] [ 11 ] За пределами Японии сезон отела кажется продолжительным (с апреля по октябрь) без явного пика. [ 11 ] В гавайских водах новорожденных дынеголовых китов наблюдают во все месяцы, кроме декабря, что позволяет предположить, что роды происходят круглый год, но пик появления новорожденных приходится на март и июнь. [ 25 ] Новорожденных дынеголовых китов наблюдали в апреле и июне на Филиппинах. [ 9 ] В Южном полушарии отел также происходит в течение длительного периода, с августа по декабрь. [ 10 ]
Сплетения
[ редактировать ]Известно, что дынеголовые киты массово выбрасываются на берег, часто группами, насчитывающими сотни человек, что свидетельствует о сильных социальных связях внутри стад этого вида. [ 17 ] Массовые выбросы дынеголовых китов на берег были зарегистрированы на Гавайях, в восточной Японии, на Филиппинах, в северной Австралии, на Мадагаскаре, в Бразилии и на островах Зеленого Мыса. Два из этих случаев массового выброса на мель были связаны с антропогенным гидролокатором, связанным с военно-морской деятельностью на Гавайях, и высокочастотным многолучевым гидролокатором, используемым для разведки нефти и газа на Мадагаскаре. [ 27 ] [ 28 ] Массовое выбрасывание на мель в заливе Ханалей, Кауаи, Гавайи, точнее описывается как «почти» массовое выбрасывание на мель, поскольку группа из более чем 150 дынеголовых китов не смогла выброситься на мель благодаря вмешательству человека. [ 27 ] Животные находились на мелководье замкнутого залива более 28 часов, а затем были загнаны обратно в более глубокие воды сотрудниками службы реагирования и волонтерами, членами сообщества, властями штата и федеральными властями. [ 25 ] [ 27 ] Известно, что в этом случае погиб только один теленок. Частота массовых выбрасываний на берег дынеголовых китов, по-видимому, увеличилась за последние 30 с лишним лет. [ 29 ]
Движение
[ редактировать ]Дынеголовые киты — быстрые пловцы; они путешествуют большими, плотно упакованными группами и могут создавать много брызг при всплытии, часто превращаются в морских свиней (многократно выпрыгивая из поверхности воды под небольшим углом) при движении на скорости, а также известны шпионам, а также могут выпрыгивать из воды. вода. [ 12 ] Дынеголовые киты могут опасаться лодок, но в некоторых регионах подходят к лодкам и катаются на носу. [ 9 ] [ 15 ]
Статус населения
[ редактировать ]Численность населения мира неизвестна, но оценки численности для крупных регионов составляют примерно 45 000 особей в восточной тропической части Тихого океана . [ 30 ] 2235 в северной части Мексиканского залива [ 31 ] а на Филиппинах — 920 в восточной части моря Сулу и 1380 в проливе Таньон между островами Себу и Негрос . [ 22 ] На Гавайях известны две популяции: популяция численностью около 450 особей, проживающая на мелководье северо-западной части острова Гавайи («постоянная популяция Кохолы»), и гораздо более крупная популяция, насчитывающая около 8000 особей, которая перемещается между основными Гавайскими островами в более глубокие воды. [ 26 ]
Поскольку постоянное население острова Гавайи имеет ограниченный ареал (наблюдения были зарегистрированы только у северо-западной стороны острова Гавайи), и иногда большая часть или все постоянное население может быть вместе в одной группе, есть некоторые опасения, что это население может подвергаться риску из-за взаимодействия с рыболовством и воздействия антропогенного шума, [ 21 ] [ 25 ] особенно в свете деятельности ВМС США в регионе, учитывая потенциальную связь между гидролокатором и массовыми выбросами на мель. [ 27 ]
Взаимодействие с людьми
[ редактировать ]Рыболовный прилов
[ редактировать ]Мелкие китообразные, такие как дынеголовые киты, уязвимы для прилова при промысле и могут быть ранены или убиты в результате взаимодействия с промыслом или запутывания в потерянных или выброшенных сетях. Небольшое количество дынеголовых китов было случайно поймано во время ярусного промысла тунца и рыбы-меч у берегов Майотты . [ 32 ] и в дрифтерном рыболовстве на Филиппинах, [ 33 ] Шри-Ланка, [ 34 ] Гана [ 35 ] и Индия. [ 36 ] При промысле кошелькового тунца в восточной тропической части Тихого океана дынеголовые киты редко попадают в прилов. [ 8 ] и не были зарегистрированы при кошельковом промысле в юго-западной части Индийского океана. [ 32 ] У отдельных дынеголовых китов наблюдаются такие травмы, как шрамы на теле и уродства спинных плавников, вероятно, из-за взаимодействия с рыболовными промыслами на Гавайях. [ 26 ] и недалеко от Майотты на Нормандских островах Мозамбика. [ 37 ] Небольшое количество раненых особей, наблюдаемое вблизи Майотты, позволяет предположить, что либо взаимодействие этого вида с пелагическим ярусным промыслом в этом регионе редкое явление, либо особи чаще погибают, чем получают ранения. [ 37 ] Смертность также вероятна и в других странах, где ведется жаберный или дрифтерный промысел, однако данные о прилове во многих регионах скудны. [ 32 ] Животные, пойманные в качестве прилова, иногда используются в качестве наживки при рыболовстве. [ 38 ]
Охота
[ редактировать ]Особей ловят для наживки или потребления человеком при промысле мелких китообразных и гарпунном промысле в нескольких регионах, включая Шри-Ланку, [ 34 ] Карибский бассейн, [ 39 ] Филиппины [ 40 ] и Индонезия. [ 41 ] В порту Дикскоув в Гане дынеголовые киты являются третьим по величине видом китообразных, вылавливаемым кустарями в качестве «морского мяса диких животных» как за счет прилова в дрейфующих жаберных сетях, так и изредка направленного вылова. рыбаками- [ 35 ] В прошлом японский загонный промысел время от времени вылавливал стада дынеголовых китов. [ 7 ] В 2017/18 году Япония увеличила предлагаемую годовую квоту на вылов до 704 особей для загонного промысла в Тайцзи . [ 42 ]
Загрязнение
[ редактировать ]Загрязнения окружающей среды, образующиеся в результате пластикового мусора, разливов нефти и сброса промышленных отходов в море, а также сельскохозяйственных стоков из наземных источников, могут привести к биоаккумуляции в морских экосистемах и представлять угрозу для дынноголовых китов (как и для всех морских китов). млекопитающие и долгоживущие потребители высокого трофического уровня). Стойкие органические загрязнители (СОЗ) – включают загрязнители окружающей среды, такие как полихлорированные дифенилы (ПХД), хлорорганические пестициды, например, дихлордифенилтрихлорэтаны (ДДТ) и гексахлорциклогексаны (ГХГ), а также броморганические соединения , такие как полибромированные дифениловые эфиры (ПБДЭ), – являются липофильными (жирорастворимыми) и может накапливаться в жире морских млекопитающих. [ 43 ] В высоких концентрациях эти загрязнители могут влиять на общее состояние здоровья, уровень гормонов и влиять как на иммунную, так и на репродуктивную системы. [ 43 ] [ 44 ] Самки с высоким уровнем загрязнения могут передавать загрязняющие вещества через плаценту или через лактацию от матери к теленку, что приводит к смертности телят. [ 45 ] [ 46 ] Было обнаружено, что в образцах жира дынеголовых китов, выброшенных на берег в Японии и на Гавайях, концентрации ПХБ превышают пороговые значения, считающиеся токсичными. [ 47 ] [ 48 ] За пределами Японии уровни ПБДЭ и соединений, родственных хлордану (ХЛ), в ворвани увеличились в течение 1980–2000 годов. [ 49 ]
Шум
[ редактировать ]Дынеголовые киты могут быть уязвимы к воздействию антропогенного (антропогенного) шума, например, связанного с деятельностью военных гидролокаторов, сейсмическими исследованиями и работами мощных многолучевых эхолотов. [ 15 ] [ 50 ] Основываясь на предыдущих событиях выбрасывания на мель, связывающих массовые выбросы на мель с гидролокатором, [ 27 ] [ 28 ] Дынеголовые киты, по-видимому, являются одним из наиболее чувствительных видов к активным гидролокаторам средней частоты (от 1 до 10 кГц), используемым в военных операциях, и другим типам гидролокаторов. [ 50 ] Для популяций, связанных с островами, например, жителей Гавайского архипелага, [ 21 ] Атолл Пальмира и Маркизские острова , [ 15 ] воздействие антропогенного шума может привести к перемещению животных из важной среды обитания. [ 50 ]
Наблюдение за китами
[ редактировать ]Регионов, в которых можно достоверно увидеть дынеголовых китов, немного, однако Гавайи, Мальдивы, Филиппины и восточная часть Карибского бассейна, особенно вокруг Доминики, являются лучшими местами, чтобы их увидеть. [ 12 ] У Международной китобойной комиссии (IWC) есть рекомендации по наблюдению за китами, призванные обеспечить минимальное беспокойство дикой природы, но не каждый оператор их придерживается. [ 51 ]
Статус сохранения
[ редактировать ]Дынеголовый кит занесен в Красный список МСОП как вид, вызывающий наименьшее беспокойство . [ 1 ] Имеется мало информации о текущих уровнях прилова и коммерческой охоты, поэтому потенциальное воздействие на популяции дынеголовых китов не определено. [ 8 ] Текущая динамика численности населения неизвестна. [ 1 ]
Вид указан в Приложении II. [ 52 ] Конвенции о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения (СИТЕС). Дынеголовый кит включен в Конвенции о сохранении мигрирующих видов диких животных Меморандум о взаимопонимании в отношении сохранения ламантинов и мелких китообразных Западной Африки и Макаронезии (Меморандум о водных млекопитающих Западной Африки) и Меморандум о взаимопонимании по (CMS). Соглашение о сохранении китообразных и их среды обитания в регионе тихоокеанских островов (Меморандум о взаимопонимании по тихоокеанским китообразным). [ 52 ] Как и все другие виды морских млекопитающих, дынеголовый кит находится под защитой в водах США в соответствии с Законом о защите морских млекопитающих (MMPA). [ 53 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Кишка, Дж.; Браунелл-младший; РЛ (2019). « Пепоноцефала электра » . Красный список исчезающих видов МСОП . 2019 : e.T16564A50369125. doi : 10.2305/IUCN.UK.2019-2.RLTS.T16564A50369125.en . Проверено 19 ноября 2021 г.
- ^ «Приложения | СИТЕС» . сайт цитирует . Проверено 14 января 2022 г.
- ^ Нишиваки М. и К.С. Норрис (1966). «Новый род Peponocephala для зубатых китообразных ( Electra Electra )». Научные отчеты Института исследования китов . 20 :95–100.
- ^ Вилструп, Дж. Т.; и др. (2011). «Митогеномно-филогенетический анализ Delphinidae с акцентом на Globicephalinae» . Эволюционная биология BMC . 11 (1): 65. дои : 10.1186/1471-2148-11-65 . ПМК 3065423 . ПМИД 21392378 .
- ^ Макгоуэн, MR; и др. (2019). «Филогеномное разрешение древа жизни китообразных с использованием захвата целевой последовательности» . Систематическая биология . 69 (3): 479–501. дои : 10.1093/sysbio/syz068 . ПМК 7164366 . ПМИД 31633766 .
- ^ Перейти обратно: а б с Бест, П.Б. и П.Д. Шонесси (1981). «Первая запись дынноголового кита Peponocephala Electra из Южной Африки». Энн. Южная Африка. Муз . 83 : 33–47.
- ^ Перейти обратно: а б с д Миядзаки, Н.; Ю. Фудзисе; К. Ивата (1998). «Биологический анализ массового выбрасывания на берег дынеголовых китов ( Peponocephalalectra ) в Аошиме, Япония». Бюллетень Национального музея науки Токио, серия A. 24 : 31–60.
- ^ Перейти обратно: а б с д и Перриман, WL; К. Данил (2018). «Дыноголовый кит: Peponocephalalectra ». В В. Ф. Перрене; Б. Вюрсиг; Дж. Г. Тьювиссен (ред.). Энциклопедия морских млекопитающих (3-е изд.). Сан-Диего, Калифорния: Эльзевир. стр. 593–595.
- ^ Перейти обратно: а б с д и ж Перриман, WL; и др. (1994). С.Х. Риджуэй и Р.Дж. Харрисон (ред.). Дынеголовый кит Peponocephala Electra Grey, 1846 . Лондон: Академическая пресса. стр. 363–386.
{{cite book}}:|work=игнорируется ( помогите ) - ^ Перейти обратно: а б с Брайден, М.; Р. Харрисон; Р. Лир (1977). «Некоторые аспекты биологии Peponocephalalectra (Cetacea: Delphinidae). I. Общая и репродуктивная биология». Морские и пресноводные исследования . 28 (6): 703–715. дои : 10.1071/MF9770703 .
- ^ Перейти обратно: а б с д и Амано, М.; и др. (2014). «История жизни и групповой состав дынеголовых китов на основе массовых выбрасываний на берег в Японии». Наука о морских млекопитающих . 30 (2): 480–493. дои : 10.1111/mms.12050 .
- ^ Перейти обратно: а б с д Карвардин, М. (2017). Путеводитель Марка Карвардина по наблюдению за китами в Северной Америке . Лондон: Издательство Блумсбери.
- ^ Перейти обратно: а б с Джефферсон, штат Техас; М. А. Уэббер; Р. Л. Питман (2015). Морские млекопитающие мира: исчерпывающее руководство по их идентификации (2-е изд.). Лондон: Академическая пресса. п. 616.
- ^ Бэрд, RW (2010). «Карликовые косатки ( Feresa attenuata ) или ложные косатки ( Pseudorca crassidens )? Идентификация группы мелких китообразных, замеченных у берегов Эквадора в 2003 году». Водные млекопитающие . 36 (3): 326. doi : 10.1578/AM.36.3.2010.326 . S2CID 87999792 .
- ^ Перейти обратно: а б с д и ж г Браунелл, Р.Л. младший; и др. (2009). «Поведение дынеголовых китов Peponocephala Electrotra вблизи океанических островов» . Наука о морских млекопитающих . 25 (3): 639–658. дои : 10.1111/j.1748-7692.2009.00281.x . S2CID 2458209 .
- ^ Перейти обратно: а б с д Вест, КЛ; и др. (2018). «Содержимое желудка и нырятельное поведение дынеголовых китов ( Peponocephalalectra ) в гавайских водах». Наука о морских млекопитающих . 34 (4): 1082–1096. дои : 10.1111/mms.12507 .
- ^ Перейти обратно: а б Джефферсон, штат Техас; Н. Б. Баррос (1997). « Пепоноцефала электра » . Виды млекопитающих (553): 1–6. дои : 10.2307/3504200 . JSTOR 3504200 .
- ^ Маллин, К.Д.; и др. (1994). «Первые наблюдения дынеголовых китов ( Peponocephala Electra ) в Мексиканском заливе». Наука о морских млекопитающих . 10 (3): 342–348. дои : 10.1111/j.1748-7692.1994.tb00488.x .
- ^ Перейти обратно: а б с д Мартиен, КК; и др. (2017). «Неожиданные закономерности глобальной структуры популяций дынеголовых китов Peponocephala Electra » . Серия «Прогресс в области морской экологии» . 577 : 205–220. Бибкод : 2017MEPS..577..205M . дои : 10.3354/meps12203 .
- ^ Бауманн-Пикеринг, С.; и др. (2015). «Акустическое поведение дынеголовых китов варьируется в зависимости от суточного цикла» . Поведенческая экология и социобиология . 69 (9): 1553–1563. дои : 10.1007/s00265-015-1967-0 . ПМЦ 4534505 . ПМИД 26300583 .
- ^ Перейти обратно: а б с Аскеттино, Дж. М.; и др. (2012). «Структура популяции дынеголовых китов ( Peponocephalalectra ) на Гавайском архипелаге: свидетельства существования множественных популяций на основе фотоидентификации» . Наука о морских млекопитающих . 28 (4): 666–689. дои : 10.1111/j.1748-7692.2011.00517.x .
- ^ Перейти обратно: а б Долар, MLL; и др. (2023). «Численность и экология распространения китообразных на центральных Филиппинах» . Журнал исследований и управления китообразными . 8 (1): 93–111. дои : 10.47536/jcrm.v8i1.706 . S2CID 257452639 .
- ^ Перейти обратно: а б Карзон, П.; и др. (2019). «Межродовое усыновление дельфинид: один пример таксономического обсуждения». Этология . 125 (9): 669–676. дои : 10.1111/eth.12916 . S2CID 198254136 .
- ^ Перейти обратно: а б Бэрд, Р.; и др. (2018). «Исследования зубатых китов на Тихоокеанском ракетном полигоне в августе 2017 года: спутниковое мечение, фотоидентификация и пассивный акустический мониторинг». Подготовлен для командующего Тихоокеанским флотом США, Перл-Харбор, Гавайи .
- ^ Перейти обратно: а б с д и Бэрд, RW (2016). Жизнь дельфинов и китов Гавайев: естественная история и охрана . Гонолулу: Гавайский университет Press.
- ^ Перейти обратно: а б с Брэдфорд, Алабама; и др. (2017). «Оценки численности китообразных на основе линейного разреза в исключительной экономической зоне Гавайских островов США» . Рыболовный вестник . 115 (2): 129–142. дои : 10.7755/FB.115.2.1 .
- ^ Перейти обратно: а б с д и Саутхолл, БЛ; и др. (2006). «Массовое выбрасывание на берег гавайского дынноголового кита ( Peponocephala Electra ) 3-4 июля 2004 г.». Технический меморандум NOAA NMFS-OPR-31, Вашингтон, округ Колумбия: Национальное управление океанических и атмосферных исследований .
- ^ Перейти обратно: а б Саутхолл, БЛ; и др. (2013). «Итоговый отчет Независимой научной экспертной группы по исследованию потенциальных факторов, способствовавших массовому выбросу на берег дынеголовых китов ( Peponocephalalectra ) в Антсохихи, Мадагаскар, в 2008 году». Независимая научная экспертная группа .
- ^ Браунелл-младший, Р.; и др. (2006). «Массовое выбрасывание на берег дынеголовых китов Peponocephala Electra : мировой обзор». Документ Международной китобойной комиссии SC/58/SM . 8 .
- ^ Уэйд, П.Р. и Т. Герродетт (1993). «Оценки численности и распространения китообразных в восточной тропической части Тихого океана». Доклад Международной китобойной комиссии . 43 : 477–493.
- ^ Уоринг, GT; и др. (2012). «Оценка запасов морских млекопитающих в Атлантике и Мексиканском заливе США, 2012 г.». Технический меморандум NOAA NMFS-NE-2232013, Национальное управление океанических и атмосферных исследований, Национальная служба морского рыболовства, Северо-восточный научный центр рыболовства, Глостер, Массачусетс : 419.
- ^ Перейти обратно: а б с Кишка, Дж.; и др. (2009). «Прилов морских млекопитающих в юго-западной части Индийского океана: обзор и необходимость комплексной оценки состояния». Журнал морских наук Западной Индийского океана . 7 (2): 119–136.
- ^ Долар, MLL; и др. (1994). «Направленный промысел китообразных на Филиппинах». Отчеты Международной китобойной комиссии . 44 : 439–449.
- ^ Перейти обратно: а б Илангакун, А. (1997). «Видовой состав, сезонные вариации, соотношение полов и длина тела мелких китообразных, пойманных у западного, юго-западного и южного побережья Шри-Ланки». Журнал Бомбейского общества естествознания . 94 : 298–306.
- ^ Перейти обратно: а б Ван Веребек, К.; Дж. С. Дебра; ПК Офори-Дансон (2014). «Высадка китообразных в рыболовном порту Дикскоув, Гана, в 2013–2014 годах: предварительная оценка». Документ Научного комитета МКК SC/65b/SM17, Блед, Словения : 12–24.
- ^ Джейабаскаран, Р.; Э. Вивеканандан (2013). «Взаимодействие морских млекопитающих и рыболовства в Индийских морях». Региональный симпозиум по экосистемным подходам к морскому рыболовству и биоразнообразию: 27–30 октября, Кочи .
- ^ Перейти обратно: а б Кишка, Дж.; Д. Пелурдо; В. Риду (2008). «Шрамы на теле и уродства спинных плавников как индикаторы взаимодействия между мелкими китообразными и рыболовством вокруг острова Майотта в проливе Мозамбика». Журнал морских наук Западной Индийского океана . 7 (2).
- ^ Минцер, виджей; К. Диниз; Т. К. Фрейзер (2018). «Использование водных млекопитающих в качестве наживки в мировом рыболовстве» . Границы морской науки . 5 : 191. дои : 10.3389/fmars.2018.00191 .
- ^ Колдуэлл, Дания; MC Колдуэлл; Р. В. Уокер (1976). «Первые находки дельфина Фрейзера ( Lagenodelphis hosei ) в Атлантике и дынеголового кита ( Peponocephala Electra ) в западной Атлантике». Цетология . 25 : 1–4.
- ^ Долар, М. (1994). «Случайный вылов мелких китообразных при рыболовстве на Палаване, центральных Висайских островах и северном Минданао на Филиппинах». Доклад Международной китобойной комиссии . 15 : 355–363.
- ^ Мустика, ПЛК (2006). «Морские млекопитающие в море Саву (Индонезия): местные знания, анализ угроз и варианты управления». Магистерская диссертация. Университет Джеймса Кука, Таунсвилл, Квинсленд .
- ^ Международная китобойная комиссия (2018). «Отчет Научного комитета». Журнал исследований и управления китообразными . 19 (Приложение): 1–114.
- ^ Перейти обратно: а б Боссарт, Г. (2011). «Морские млекопитающие как виды-сторожа океанов и здоровья человека». Ветеринарная патология . 48 (3): 676–690. дои : 10.1177/0300985810388525 . ПМИД 21160025 .
- ^ Дефорж, Ж.-П.В.; и др. (2016). «Иммунотоксическое воздействие загрязнителей окружающей среды на морских млекопитающих». Интернационал окружающей среды . 86 : 126–139. дои : 10.1016/j.envint.2015.10.007 . ПМИД 26590481 .
- ^ Шваке, Л.Х.; и др. (2002). «Вероятностная оценка риска репродуктивного воздействия полихлорированных бифенилов на афалин ( Tursiops truncatus ) с юго-восточного побережья США» . Экологическая токсикология и химия . 21 (12): 2752–2764. дои : 10.1002/etc.5620211232 . ПМИД 12463575 . S2CID 17283029 .
- ^ Боррель, А.; Блох, Д.; Депортес, Г. (1995). «Возрастные тенденции и репродуктивная передача хлорорганических соединений у длинноплавниковых гринд с Фарерских островов». Загрязнение окружающей среды . 88 (3): 283–292. дои : 10.1016/0269-7491(95)93441-2 . ПМИД 15091540 .
- ^ Бахман, MJ; и др. (2014). «Постоянные концентрации органических загрязнителей в жире 16 видов китообразных, выброшенных на острова Тихого океана с 1997 по 2011 год». Наука об общей окружающей среде . 488 : 115–123. Бибкод : 2014ScTEn.488..115B . doi : 10.1016/j.scitotenv.2014.04.073 . ПМИД 24821437 .
- ^ Кадзивара, Н.; и др. (2006). «Географическое распространение полибромдифениловых эфиров (ПБДЭ) и хлорорганических соединений у мелких китообразных из вод Азии». Хемосфера . 64 (2): 287–295. Бибкод : 2006Chmsp..64..287K . doi : 10.1016/j.chemSphere.2005.12.013 . ПМИД 16439003 .
- ^ Кадзивара, Н.; и др. (2008). «Полибромдифениловые эфиры (ПБДЭ) и хлорорганические соединения у дынеголовых китов Peponocephalalectra , массово выброшенных на берег вдоль японского побережья: материнский перенос и временная тенденция». Загрязнение окружающей среды . 156 (1): 106–114. дои : 10.1016/j.envpol.2007.12.034 . ПМИД 18272274 .
- ^ Перейти обратно: а б с Форни, Калифорния; и др. (2017). «Деваться некуда: оценки воздействия шума на популяции морских млекопитающих с высокой точностью определения местоположения» . Исследования исчезающих видов . 32 : 391–413. дои : 10.3354/esr00820 .
- ^ «Общие принципы IWC по наблюдению за китами» . Международная китобойная комиссия. Архивировано из оригинала 31 декабря 2019 года . Проверено 20 ноября 2019 г.
- ^ Перейти обратно: а б ЮНЕП. «Вид+» . Веб-сайт Виды+. Найроби, Кения . Составлено UNEP-WCMC, Кембридж, Великобритания . Проверено 10 октября 2019 г.
- ^ Служба охраны рыбы и дикой природы США. «Закон о защите морских млекопитающих» . Международные дела . Проверено 10 октября 2019 г.
Дальнейшее чтение
[ редактировать ]- Карвардин, М., 2019. Справочник по китам, дельфинам и морским свиньям. Блумсбери, Лондон. 528 стр.
- Джефферсон, Т.А., М.А. Уэббер и Р.Л. Питман, 2015. Морские млекопитающие мира: комплексное руководство по их идентификации. 2-е изд., Лондон: Academic Press. 616 стр.
Внешние ссылки
[ редактировать ]- Мама-дельфин впервые усыновила китового детеныша
- Редкий гибрид дельфина и кита замечен возле Гавайев
- Звуки дынноголового кита
- Общество морской маммологии
- Каскадия Исследования
- НОАА по рыболовству
- IWC Ответственное наблюдение за китами/Справочник
- Меморандум о взаимопонимании относительно сохранения ламантинов и мелких китообразных Западной Африки и Макаронезии .
- Официальная веб-страница Меморандума о взаимопонимании по сохранению китообразных и их среды обитания в регионе островов Тихого океана.
- Виды, находящиеся в Красном списке МСОП, вызывающие наименьшее беспокойство
- Китообразные Атлантического океана
- Китообразные Индийского океана
- Китообразные Тихого океана
- Млекопитающие Доминиканской Республики
- Океанические дельфины
- Млекопитающие, описанные в 1846 г.
- Таксоны, названные Джоном Эдвардом Греем