Молекулярная антропология
Молекулярная антропология , также известная как генетическая антропология , представляет собой изучение того, как молекулярная биология способствовала пониманию эволюции человека. [ 1 ] Эта область антропологии исследует эволюционные связи между древними и современными человеческими популяциями, а также между современными видами. Как правило, сравнения проводятся между последовательностями, последовательностями ДНК или белка ; Однако ранние исследования использовали сравнительную серологию .
Изучая последовательности ДНК в разных популяциях, ученые могут определить близость взаимосвязи между популяциями (или внутри населения). Некоторые сходства в генетическом составе позволяют молекулярным антропологам определять, принадлежат ли разные группы людей одну и ту же гаплогруппу , и, таким образом, если они разделяют общее географическое происхождение. Это важно, потому что это позволяет антропологам проследить закономерности миграции и поселения , что дает полезное представление о том, как современные группы населения сформировались и прогрессировали с течением времени. [ 2 ]
Молекулярная антропология была чрезвычайно полезна для установления эволюционного дерева людей и других приматов , в том числе тесно связанных видов, таких как шимпанза и горилл. Несмотря на то, что между людьми и шимпанзе явно существует много морфологических сходств, определенные исследования также пришли к выводу, что между ДНК обоих видов существует примерно 98 процентов общности. [ Цитация необходима ] Тем не менее, более поздние исследования изменили общность 98 процентов до общности 94 процента, показав, что генетический разрыв между людьми и шимпанзе больше, чем первоначально думали. [ 3 ] Такая информация полезна при поиске общих предков и лучше понимать, как эволюционировали люди.
Гаплоидные локусы в молекулярной антропологии
[ редактировать ]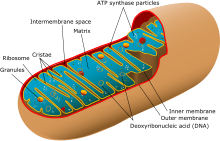
У людей есть две группы непрерывных связей , которые переносятся одним полом. Первая - Y -хромосома , которая передается от отца к сыну. Анатомические женщины несут Y -хромосому только в результате генетического дефекта. Другая группа сцепления - митохондриальная ДНК (мтДНК). МтДНК почти всегда передается только следующим поколениям женщинами, но при очень исключительных обстоятельствах мтДНК может передаваться через мужчин. [ нужно разъяснения ] Нерекомбинантная часть Y-хромосомы и мтДНК при нормальных обстоятельствах не подвергается продуктивной рекомбинации. Часть Y -хромосомы может подвергаться рекомбинации с Х -хромосомой, а в истории обезьяны граница изменилась. Такие рекомбинантные изменения в нерекомбинантной области Y чрезвычайно редки. [ Цитация необходима ]
Митохондриальная ДНК
[ редактировать ]
Митохондриальная ДНК стала области исследований в области филогенетики в конце 1970 -х годов. В отличие от геномной ДНК, она дала преимущества в том, что она не подвергалась рекомбинации. Процесс рекомбинации, если он достаточно частый, повреждает способность создавать экономные деревья из -за участков аминокислотных подчинений (SNP). [ нужно разъяснения ] При просмотре между отдаленно родственными видами рекомбинация является меньшей проблемой, поскольку рекомбинация между ветвями от общих предков предотвращается после возникновения истинного видообразования. При изучении тесно связанных видов или разветвления внутри видов рекомбинация создает большое количество «нерелевантных SNP» для кладистического анализа. МтДНК, благодаря процессу деления органелл, со временем стала клональной; Очень мало, или часто ни одного, от этой отцовской мтДНК проходит. Хотя рекомбинация может происходить в мтДНК, существует небольшой риск, что она будет передана следующему поколению. В результате мтДНК становятся клональными копиями друг друга, за исключением случаев, когда возникает новая мутация. В результате у мтДНК нет ловушек аутосомных локусов при изучении в межзообразных группах. Другое преимущество мтДНК заключается в том, что гипер-варизовые области развиваются очень быстро; Это показывает, что определенные области митохондриальной ДНК приближаются к нейтральности. Это позволило использовать митохондриальную ДНК определить, что относительный возраст человеческого населения был небольшим, что прошло через недавнее сужение около 150 000 лет назад (см. #CAUSE ошибок ).
Митохондриальная ДНК также использовалась для проверки близости шимпанзе к людям по сравнению с гориллами , а также для проверки взаимосвязи этих трех видов относительно орангутанов .

Совсем недавно, [ когда? ] Геном мтДНК использовался для оценки паттернов ветвления у народов по всему миру, например, когда Новый Свет был урегулирован и как. Проблема с этими исследованиями заключалась в том, что они в значительной степени полагаются на мутации в области кодирования. Исследователи все чаще обнаруживают, что по мере того, как люди переезжали из юго-восточных регионов Африки, что в регионе кодирования накопилось больше мутаций, чем ожидалось, и в прохождении в Новый Света считаются некоторые группы [ Цитация необходима ] Переход от азиатских тропиков в Сибири в древний земельный регион под названием Берингия и быстро мигрировал в Южную Америку. Многие из мтДНК имеют гораздо больше мутаций и в редко мутированных кодирующих сайтах относительно ожиданий нейтральных мутаций.
Митохондриальная ДНК предлагает еще одно преимущество перед аутосомной ДНК. Как правило, в каждой клетке имеется от 2 до 4 копий каждой хромосомы (от 1 до 2 от каждой родительской хромосомы). Для мтДНК в каждой клетке могут быть десятки до сотен. Это увеличивает количество каждого локуса мтДНК, по крайней мере, на величину. Для древней ДНК, в которой ДНК сильно разлагается, количество копий ДНК полезно для расширения и соединения коротких фрагментов вместе и уменьшает количество кости, извлеченной из очень ценных ископаемых/древних останков. В отличие от Y -хромосомы, как мужчин, так и женских останков несут мтДНК примерно в равных количествах.

Y -хромосома
[ редактировать ]
Y -хромосома обнаруживается в ядре нормальных клеток ( ядерная ДНК ). В отличие от мтДНК, он имеет мутации в нерекомбинантной части (NRY) хромосомы, расположенных широко распределенным, так далеко друг от друга, что обнаружение мутаций на новых Y-хромосомах является трудоемким по сравнению с мтДНК. Многие исследования полагаются на тандемные повторения; Тем не менее, тандемные повторения могут быстро расширяться и втягивать и в некоторых предсказуемых закономерностях. Y -хромосома только отслеживает мужские линии и не обнаруживается у женщин, тогда как мтДНК можно проследить у мужчин, даже если они не проходят на мтДНК. Кроме того, было подсчитано, что эффективные мужские популяции в доисторический период, как правило, были двумя женщинами на мужчине, и недавние исследования показывают, что культурная гегемония играет большую роль в прохождении Y. Это создало несоответствие между мужчинами и женщинами для того времени самому последнему общему предку (TMRCA). Оценки Y TMRCA варьируются от 1/4 до менее 1/2, а также MtDNA TMRCA. Неясно, связано ли это с высокими соотношениями мужчин и женских в прошлом в сочетании с повторными миграциями из Африки, в результате изменения мутационной скорости, или, как некоторые даже предположили, что женщины LCA между шимпанзе и людьми продолжали Передайте ДНК миллионы после того, как мужчины перестали пройти ДНК. В настоящее время наилучшие данные свидетельствуют о том, что при миграции соотношение мужчин и женщин у людей, возможно, снизилось, что привело к обрезке Y -разнообразия в нескольких случаях внутри и за пределами Африки.

Для молекулярной филогенетики и молекулярного сжима, Y-хромосома очень эффективна и создает вторую перспективу. Одним из аргументов, который возник, было то, что маори мтДНК, по -видимому, мигрировали из восточного Китая или Тайваня, Y -хромосомой из региона Папуа -Новой Гвинеи. Когда гаплотипы HLA использовались для оценки двух гипотез, было обнаружено, что оба были правы, что маори были смещенной популяцией. Такие добавки, по -видимому, являются распространенными в человеческой популяции, и, следовательно, использование одного гаплоидного локуса может дать предвзятую перспективу.
X-связанные исследования
[ редактировать ]X-хромосома также является формой ядерной ДНК. Поскольку он найден в 1 копии у мужчин и 2 недентичных хромосома у женщин, он имеет плоидность 1,5. Тем не менее, у людей эффективная плоидность несколько выше, ~ 1,7, поскольку женщины в размножающейся популяции имели тенденцию превзойти мужчины на 2: 1 во время большой части человеческой предыстории. Как и мтДНК, X-связанная ДНК имеет тенденцию чрезмерно подчеркивать историю женской популяции гораздо больше, чем мужчина. Было проведено несколько исследований локусов на Х -хромосоме, в общей сложности было исследовано 20 участков. К ним относятся PDHA1, PDHA1, XQ21.3, XQ13.3, ZFX , FIX, IL2RG, PLP, GK, IDS, ALAS2, RRM2P4, AMEIX, TNFSF5, LICAM и MSN. Время до самого последнего общего предка (TMRCA) варьируется от закрепленного до ~ 1,8 миллиона лет, со срединной около 700К. Эти исследования примерно запланированы к ожидаемому распределению аллелей фиксации с учетом неравновесного сцепления между соседними участками. Для некоторых аллелей точка происхождения неуловима, для других точка происхождения направлен на Африку к югу от Сахары. В SSA есть некоторые различия, которые предполагают меньшую область, но нет достаточно достаточного размера выборки и охвата, чтобы определить место самого последнего общего предка. TMRCA соответствует и расширяет узкое место, подразумеваемое мтДНК, уверенно до 500 000 лет.
Аутосомные локусы
[ редактировать ]
Изменение скорости
[ редактировать ]Этот раздел пуст. Вы можете помочь, добавив к этому . ( Июль 2010 г. ) |
Древнее секвенирование ДНК
[ редактировать ]Крингс -неандертальская мтДНК была секвенирована, а сходство последовательности указывает на столь же недавнее происхождение из небольшой популяции на неандертальской ветви покойных гоминидов . Ген MCR1 также был секвенирован, но результаты являются противоречивыми, причем одно исследование утверждает, что проблемы загрязнения не могут быть решены из неандертальных сходств человека. Критически, однако, не была получена последовательность ДНК из Homo erectus , Homo floresiensis или любого из других поздних гоминидов. Некоторые из полученных древних последовательностей имеют весьма вероятные ошибки и правильный контроль, чтобы избежать загрязнения.

Причины ошибок
[ редактировать ]Молекулярная филогенетика основана на количественных заменах, а затем сравнивая последовательность с другими видами, в процессе есть несколько точек, которые создают ошибки. Первая и самая большая проблема - найти «якоря», которые позволяют исследованию калибровать систему. В этом примере существует 10 мутаций между шимпанзе и людьми, но у исследователя нет известных окаменелостей, которые являются приятными наследственными, но не являются наследственными для следующего вида на дереве, горилла . Тем не менее, есть окаменелости, которые, как полагают, являются наследственными для орангутанов и людей, примерно 14 миллионов лет назад. Так, чтобы исследователь мог использовать орангутанское и человеческое сравнение и придумать разницу в 24. Используя это, он может оценить (24/(14*2, «2» предназначен для длины ветви для человека (14my) и Орангутана (14 My) от их последнего общего предка (LCA). 100 нт в длину, эта ставка составит 0,00857/нт за миллион лет.
Проблема калибровки
[ редактировать ]Есть несколько проблем, которые не видели в вышеперечисленном. Во -первых, мутации происходят как случайные события. Во -вторых, вероятность того, что любой сайт в геноме варьируется, отличается от следующего сайта, очень хороший пример - кодоны для аминокислот, первые два NT в кодоне могут мутировать на 1 на миллиард лет, но третий NT может мутировать 1 за миллион лет. Если ученый не изучит последовательность многих животных, особенно близких к изучению ветви, они, как правило, не знают, какая скорость мутации для данного сайта. Мутации действительно встречаются в 1 -й и 2 -й позициях кодонов, но в большинстве случаев эти мутации находятся под отрицательным отбором и поэтому удаляются из популяции в течение небольших периодов времени. При определении скорости эволюции в якоре есть проблема, которую создает случайная мутация. Например, скорость 0,005 или 0,010 также может объяснить 24 мутации в соответствии с распределением биномиальной вероятности . Некоторые из мутаций, которые произошли между ними, вернулись, скрывая изначально более высокую скорость. Выбор может сыграть в этом, редкая мутация может быть селективной в точке x во времени, но более поздний климат может измениться, или виды мигрируются, и он не более селективный, а давление оказывается на новые мутации, которые возвращают изменения, а иногда и возврат может возникнуть NT, тем больше расстояние между двумя видами, тем больше вероятность, что это произойдет. Кроме того, из этого наследственных видов оба вида могут случайным образом мутировать сайт с одним и тем же нуклеотидом. Много раз это может быть разрешено путем получения образцов ДНК от видов в ветвях, создав экономное дерево, в котором можно вывести порядок мутации, создавая диаграмму длины ветвления. Затем эта диаграмма будет провести более точную оценку мутаций между двумя видами. Статистически можно назначить дисперсию, основанную на проблеме случайности, мутаций задних и параллельных мутаций (гомоплазий) при создании диапазона ошибок.
Однако есть еще одна проблема в калибровке, которая бросила вызов статистическому анализу. Существует истинное/ложное обозначение ископаемого на наименее распространенный предок. В действительности, вероятность наличия наименее распространенного предка из двух существующих видов в качестве якоря, часто, часто, что ископаемое уже лежит в одной ветви (недооценивая возраст), лежит в третьей ветви (недооценивая возраст) или в случае существования В рамках видов LCA, возможно, было миллионы лет старше ветви. На сегодняшний день единственный способ оценить эту дисперсию - применять молекулярную филогенетику на виды, которые, как утверждают, являются ветвиными точками. Это только, однако определяет «отдаленные» точки привязки. И поскольку более вероятно, что более распространенные окаменелости моложе, чем точка ветви, отдаленные ископаемые могут быть просто редким старшим представителем. Эти неизвестные создают неопределенность, которая трудно количественно оценить, и часто не предпринимаются.
Недавние документы смогли оценить, примерно, дисперсию. Общая тенденция как новые окаменелости обнаружены, заключается в том, что более старые окаменелости недооценили возраст точки филиала. В дополнение к этой датировке окаменелости имели историю ошибок, и было много пересмотренных дат. Возраст, назначенный исследователями на некоторые крупные филиалы, почти удвоился за последние 30 лет. Отличным примером этого является дебаты о LM3 (озеро Mungo 3) в Австралии. Первоначально он был датируется примерно 30 ky при получении углерода, датирование углеродом имеет проблемы, однако, для отображения более 20 часов в возрасте и серьезных проблем для образцов в возрасте около 30 часов. Другое исследование рассмотрело ископаемое и оценило возраст в возрасте 62 тысяч.
В тот момент, когда кто-то имеет оценку скорости мутаций, учитывая вышеупомянутое, должно быть два источника дисперсии, которые необходимо перекрестно подготовить, чтобы генерировать общую дисперсию. Это редко делается в литературе.
Проблемы при оценке TMRCA
[ редактировать ]Время до последнего общего предка ( TMRCA ) объединяет ошибки в калибровке с ошибками при определении возраста местной ветви.
История
[ редактировать ]Протеиновая эра
[ редактировать ]
С ДНК, недавно обнаруженной в качестве генетического материала, в начале 1960 -х годов секвенирование белка начало взлетать. [ 4 ] Секвенирование белка началось на цитохроме C и гемоглобине. Герхард Браунитцер секвенировал гемоглобин и миоглобин , в общей сложности было выполнено более сотен последовательностей из широких видов. В 1967 году AC Wilson начал продвигать идею «молекулярных часов». К 1969 году молекулярное схему было применено к эволюции антропоидов, а В. Сарич и А.С. Уилсон обнаружили, что альбумин и гемоглобин имеют сопоставимые показатели эволюции, что указывает на то, что шимпанза и люди расщепляются примерно на 4-5 миллионов лет назад. [ 5 ] В 1970 году Луи Лики столкнулся с этим выводом, выступая за неправильную калибровку молекулярных часов. [ 6 ] К 1975 году секвенирование белка и сравнительная серология , объединенные, использовались для предположения, чтобы люди были самыми близкими живыми относительноми (как видами ), был шимпанзе. [ 7 ] Оглядываясь назад, последний общий предок (LCA) от людей и шимпанзе представляется старше, чем Сарича и Уилсон , но не так старый, как заявил Лики. Тем не менее, Лики был прав в дивергенции старых и новых обезьян, стоимость Сарича и Уилсона использовали значительную недооценку. Эта ошибка в возможности прогноза подчеркивает общую тему. (См. Причины ошибки )
Эра ДНК
[ редактировать ]
Гибридизация RLFP и ДНК
[ редактировать ]В 1979 году Wmbrown и Wilson начали смотреть на эволюцию митохондриальной ДНК у животных и обнаружили, что они быстро развиваются. [ 8 ] Техника, которую они использовали, представлял собой полиморфизм длины фрагмента ограничения ( RFLP ), который был более доступным в то время по сравнению с секвенированием. В 1980 году WM Brown, рассматривая относительные различия между человеческими и другими видами, признала недавнее сужение ( 180 000 лет назад) в человеческой популяции. [ 9 ] Год спустя Браун и Уилсон смотрели на фрагменты RFLP и определили, что население расширилось в последнее время, чем другие популяции обезьян. [ 10 ] В 1984 году была сделана первая последовательность ДНК от вымершего животного. [ 11 ] Sibley и Ahlquist применяют технологию гибридизации ДНК-ДНК к антропоидной филогении и видят раскол PAN/человека ближе, чем горилла/пан или горилла/человек, очень спорное утверждение. [ 12 ] [ 13 ] Однако в 1987 году они смогли поддерживать свои претензии. [ 14 ] В 1987 году Cann, Stoneking и Wilson предполагают, что RFLP -анализ митохондриальной ДНК человека, что люди эволюционировали из Африки в Африке одной женщины в небольшой популяции, ~ 10,00 человек, 200 000 лет назад. [ 15 ]
Эра ПЦР
[ редактировать ]
В 1987 году для определения последовательностей была впервые использована ПЦР-амплификация мтДНК. [ 16 ] В 1991 году Vigilante et al. Опубликована оригинальная работа по филогениям мтДНК, подразумевающей Африку к югу от Сахары как место самых последних общих предков людей для всех мтдн. [ 17 ] Война между внеафриканскими и мультирегионализмом, уже кипящая с критикой Аллана Темплтона, вскоре возросла с палеоантропологом, таким как Милфорд Вольпофф, вступив в воображение. [ 18 ] [ 19 ] [ 20 ] В 1995 году Ф. Айала опубликовал свою статью «Критическая наука» «Миф о Еве», которая опиралась на HLA-DR . последовательность [ 21 ] В то время, однако, Аяла не знала о быстрой эволюции локусов HLA посредством рекомбинационного процесса. В 1996 году Parham и Ohta опубликовали свои находки о быстрой эволюции HLA с помощью рекомбинации короткой дистанции («обращение генов» или «абортивную рекомбинацию»), ослабляя претензию Аялы (Пархам фактически написал обзор годом ранее, но это ушло незаметно). [ 22 ] [ 23 ] Поток документов следовал с обеих сторон, многие из которых с очень ошибочными методами и выборкой. Один из самых интересных [ Согласно кому? ] Был Харрис и Эй, 1998 год, что показало, что TMCRA (время до самого последнего общего предка) для гена PDHA1 значительно превышала 1 миллион лет. Учитывая плоидность в этом локусе в 1,5 (в 3 раза выше мтДНК), TMRCA была более чем вдвое превышает ожидание. В то время как это попадает в «кривую фиксации» 1,5 плюиды (в среднем 2 женщины и 1 мужчина), предполагаемый возраст 1,8 мой близок к значительно девиантному значению p для размера популяции, возможно, указывая на то, что человеческое население сократилось или раскололось Другое население. [ 24 ] Как ни странно, в следующих x-связанных локусах, которые они исследовали, фактор IX, показал TMRCA менее 300 000 лет. [ 25 ]

Древняя ДНК
[ редактировать ]Древнее секвенирование ДНК проводилось в ограниченном масштабе до конца 1990-х годов, когда персонал в Институте Макса Планка шокировал мир антропологии, секвенируя ДНК от около 40 000-летнего неандертальца . [ 26 ] [ 27 ] [ 28 ] Результатом этого эксперимента является то, что различия между людьми, живущими в Европе, многие из которых были получены из гаплогруппы H (CRS), неандертальцы разветвлялись от людей более чем за 300 000 лет до того, как гаплогруппа H достигла Европы. В то время как мтДНК и другие исследования продолжали поддерживать уникальное недавнее африканское происхождение, это новое исследование в основном ответило на критику с неандертальской стороны.
Геномное секвенирование
[ редактировать ]Значительный прогресс был достигнут в секвенировании генома с тех пор, как Ингман и его коллега опубликовали свои выводы о митохондриальном геноме. [ 29 ] Было опубликовано несколько статей по геномной мтДНК; Существует значительная изменчивость в ставке эволюции, а изменение и выбор ставок очевидны на многих участках. В 2007 году Gonder et al. предположил, что основная популяция людей, с наибольшим уровнем разнообразия и самым низким отбором, когда -то жила в регионе Танзании и проксимальных частях южной Африки, поскольку люди покинули эту часть Африки, митохондрии выборочно развивались в новые регионы. [ 30 ]
Критический прогресс
[ редактировать ]Критическая в истории молекулярной антропологии:
- Эта молекулярная филогенетика может конкурировать со сравнительной антропологией за определение близости видов к людям.
- Уилсон и Кинг осознали в 1975 году, что, хотя между уровнем молекулярной эволюции имелся равенство , разветвляется от шимпанзе до человека до предполагаемого LCA, в морфологической эволюции наблюдалось неравенство. Сравнительная морфология, основанная на окаменелостях, может быть смещена по различным показателям изменений. [ 7 ]
- Осознание того, что в ДНК есть несколько независимых сравнений. Два метода, мтДНК и гибридизация сходятся на одном ответе, шимпанза как вида наиболее тесно связаны с людьми.
- Способность разрешать размеры популяции на основе 2n правила, предложенного Кимурой в 1950 -х годах. [ 31 ] Использовать эту информацию для сравнения относительных размеров населения и прийти к выводу о изобилии, которое противопоставляло наблюдения на основе палеонтологической записи. В то время как человеческие окаменелости в раннем и среднем каменном веке гораздо более распространены, чем шимпанзе или горилла, мало однозначных ископаемых из шимпанзе или гориллы с того же периода
Локусы, которые использовались в молекулярной филогенетике:
- Цитохром c
- Сывороточный альбумин
- Hemoglobin - Braunitizer, 1960s, Harding et al. 1997
- Митохондриальная D -петля - Wilson Group, 1980, 1981, 1984, 1987, 1989, 1991 (посмертно) - TMRCA около 170 Kya.
- Y-хромосома
- HLA -DR - Ayala 1995 - TMRCA для локуса составляет 60 миллионов лет.
- CD4 (интрон) - Тишкофф, 1996 - Большая часть разнообразия находится в Африке.
- PDHA1 (X -связанный) Харрис и Эй - TMRCA для локуса более 1,5 миллионов лет.
Xlinked loci: pdha1, xq21.3, xq13.3, zfx , fix, il2rg, plp, gk, ids, alas2, rrm2p4, ameix, tnfsf5, licam и msn
Аутосомно: многочисленные.
Ссылки
[ редактировать ]- ^ Брэдли, Бренда Дж. (2008). «Реконструкция филогений и фенотипов: молекулярный взгляд на эволюцию человека» . Журнал анатомии . 212 (4): 337–353. doi : 10.1111/j.1469-7580.2007.00840.x . PMC 2409108 . PMID 18380860 .
- ^ Коттак, Конрад Филипп. Окна на человечестве . Нью-Йорк: McGraw-Hill, 2005.
- ^ «Люди и шимпанза: близко, но не так близко» . Scientific American. 2006-12-19. Архивировано из оригинала 2007-10-11 . Получено 2006-12-20 .
- ^ Acwilson и Nokaplan (1963) Ферменты и нуклеиновые кислоты в систематике. Труды XVI International Congress of Zoology Vol.4, стр.125-127.
- ^ Wilson AC, Sarich VM (август 1969 г.). «Молекулярная шкала времени для эволюции человека» . Прокурор Нат. Академический Наука США . 63 (4): 1088–93. Bibcode : 1969pnas ... 63.1088W . doi : 10.1073/pnas.63.4.1088 . PMC 223432 . PMID 4982244 .
- ^ Leakey LS (октябрь 1970 г.). «Отношения африканских обезьян, человек и обезьян Старого Света» . Прокурор Нат. Академический Наука США . 67 (2): 746–8. Bibcode : 1970pnas ... 67..746L . doi : 10.1073/pnas.67.2.746 . PMC 283268 . PMID 5002096 .
- ^ Jump up to: а беременный Кинг MC, Wilson AC (апрель 1975 г.). «Эволюция на двух уровнях у людей и шимпанзе». Наука . 188 (4184): 107–16. Bibcode : 1975sci ... 188..107K . doi : 10.1126/science.1090005 . PMID 1090005 .
- ^ Браун Wm, George M, Wilson AC (апрель 1979 г.). «Быстрая эволюция митохондриальной ДНК животных» . Прокурор Нат. Академический Наука США . 76 (4): 1967–71. Bibcode : 1979pnas ... 76.1967b . doi : 10.1073/pnas.76.4.1967 . PMC 383514 . PMID 109836 .
- ^ Браун WM (июнь 1980 г.). «Полиморфизм в митохондриальной ДНК людей, как выявлено с помощью анализа эндонуклеазы ограничения» . Прокурор Нат. Академический Наука США . 77 (6): 3605–9. Bibcode : 1980pnas ... 77.3605b . doi : 10.1073/pnas.77.6.3605 . PMC 349666 . PMID 6251473 .
- ^ Ferris SD, Brown WM, Davidson WS, Wilson AC (октябрь 1981 г.). «Обширный полиморфизм в митохондриальной ДНК обезьян» . Прокурор Нат. Академический Наука США . 78 (10): 6319–23. Bibcode : 1981pnas ... 78.6319f . doi : 10.1073/pnas.78.10.6319 . PMC 349030 . PMID 6273863 .
- ^ Higuchi R, Bowman B, Freiberger M, Ryder OA, Wilson AC (1984). «Последовательности ДНК из квагги, вымерший член семьи лошадей». Природа . 312 (5991): 282–4. Bibcode : 1984natur.312..282H . doi : 10.1038/312282A0 . PMID 6504142 . S2CID 4313241 .
- ^ Sibley CG, Ahlquist JE (1984). «Филогения гоминоидных приматов, о чем свидетельствует гибридизация ДНК-ДНК». J. Mol. Эвол . 20 (1): 2–15. Bibcode : 1984jmole..20 .... 2S . doi : 10.1007/bf02101980 . PMID 6429338 . S2CID 6658046 .
- ^ Templeton AR (сентябрь 1985 г.). «Филогения приматов гоминоидов: статистический анализ данных гибридизации ДНК-ДНК» . Мол Биол. Эвол . 2 (5): 420–33. doi : 10.1093/oxfordjournals.molbev.a040363 . PMID 3939706 .
- ^ Sibley CG, Ahlquist JE (1987). «Свидетельство гибридизации ДНК гоминоидной филогения: результаты расширенного набора данных». J. Mol. Эвол . 26 (1–2): 99–121. Bibcode : 1987jmole..26 ... 99S . doi : 10.1007/bf02111285 . PMID 3125341 . S2CID 40231451 .
- ^ Cann RL, Stoneking M, Wilson AC (1987). «Митохондриальная ДНК и эволюция человека». Природа . 325 (6099): 31–6. Bibcode : 1987natur.325 ... 31c . doi : 10.1038/325031A0 . PMID 3025745 . S2CID 4285418 .
- ^ Wrischnik LA, Higuchi RG, Stoneking M, Erlich HA, Arnheim N, Wilson AC (январь 1987 г.). «Мутации длины в митохондриальной ДНК человека: прямое секвенирование ферментативно амплифицированной ДНК» . Нуклеиновые кислоты Res . 15 (2): 529–42. doi : 10.1093/nar/15.2.529 . PMC 340450 . PMID 2881260 .
- ^ Бдительный Л., Стоункин М., Харпендинг Х, Хоукс К., Уилсон А.С. (сентябрь 1991 г.). «Африканские популяции и эволюция митохондриальной ДНК человека». Наука . 253 (5027): 1503–7. Bibcode : 1991sci ... 253.1503V . doi : 10.1126/science.1840702 . PMID 1840702 .
- ^ Templeton AR (1993). «Гипотеза« Ева »: генетическая критика и повторный анализ». Американский антрополог . 95 : 51–72. doi : 10.1525/aa.1993.95.1.02a00030 .
- ^ Thorne A и Wolpoff M. Многорегиональная эволюция людей . Scientific American (апрель) с. 28-33 (1992)
- ^ Wolpoff M и Thorne A. Дело против Евы. Новый ученый (1991) с. 37-41.
- ^ Айала Ф.Дж. (декабрь 1995 г.). «Миф о Еве: молекулярная биология и человеческое происхождение» . Наука . 270 (5244): 1930–6. Bibcode : 1995sci ... 270.1930a . doi : 10.1126/science.270.5244.1930 . PMID 8533083 .
- ^ Parham P, Ohta T (апрель 1996 г.). «Популяционная биология антигена презентации молекулами класса I MHC». Наука . 272 (5258): 67–74. Bibcode : 1996sci ... 272 ... 67p . doi : 10.1126/science.272.5258.67 . PMID 8600539 . S2CID 22209086 .
- ^ Parham P, Adams EJ, Arnett KL (февраль 1995 г.). «Происхождение HLA-A, B, C полиморфизм». Иммунол. Преподобный 143 : 141–80. doi : 10.1111/j.1600-065x.1995.tb00674.x . PMID 7558075 . S2CID 39486851 .
- ^ Харрис Э. Э. Эй, Дж. (Март 1999 г.). «Х -хромосома доказательства древних человеческих историй» . Прокурор Нат. Академический Наука США . 96 (6): 3320–4. Bibcode : 1999pnas ... 96.3320h . doi : 10.1073/pnas.96.6.3320 . PMC 15940 . PMID 10077682 .
- ^ Harris Ee, Hey J (май 2001 г.). «Человеческие популяции показывают уменьшенное изменение последовательности ДНК в локусе фактора IX» . Карт Биол . 11 (10): 774–8. Bibcode : 2001cbio ... 11..774H . doi : 10.1016/s0960-9822 (01) 00223-8 . PMID 11378388 .
- ^ Хандт О, Хосс М., Кингс М., Пябо С (июнь 1994 г.). «Древняя ДНК: методологические проблемы» Экспериментация 50 (6): 524–9 Doi : 10.1007/ bf01921720 8020612PMID 6742827S2CID
- ^ Хэндт О, Крингс М., Уорд Р.Х., Пябо С (август 1996 г.). «Извлечение древних последовательностей ДНК человека» . Являюсь. J. Hum. Генет . 59 (2): 368–76. PMC 1914746 . PMID 8755923 .
- ^ Крингс М., Стоун А., Шмитц Р.В., Краницки Х, Стоункин М., Пябо С (июль 1997 г.). «Неандертальские последовательности ДНК и происхождение современных людей». Клетка . 90 (1): 19–30. doi : 10.1016/s0092-8674 (00) 80310-4 . HDL : 11858/00-001M-0000-0025-0960-8 . PMID 9230299 . S2CID 13581775 .
- ^ Ингман М., Кассманн Х, Пябо С., Джилленстен У (декабрь 2000 г.). «Изменение митохондриального генома и происхождение современных людей». Природа . 408 (6813): 708–13. Bibcode : 2000natur.408..708i . doi : 10.1038/35047064 . PMID 11130070 . S2CID 52850476 .
- ^ Гондер М.К., Мортенсен Х.М., Рид Ф.А., де Соуза А., Тишкофф С.А. (март 2007 г.). «Анализ последовательности генома всего мтДНК древних африканских линий» . Мол Биол. Эвол . 24 (3): 757–68. doi : 10.1093/molbev/msl209 . PMID 17194802 .
- ^ Кимура М (май 1954). «Процесс, приводящий к квазификсации генов в естественных популяциях из-за случайных колебаний интенсивности отбора» . Генетика . 39 (3): 280–95. doi : 10.1093/Genetics/39.3.280 . PMC 1209652 . PMID 17247483 .