Pampaphoneus
| Pampaphoneus Временный диапазон: Гвадалупиан ( Вордиан ),
| |
|---|---|

| |
| Pampaphoneus biccai голотип черепа | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Synapsida |
| Клада : | Терапсида |
| Подотряд: | † Dinocephalia |
| Семья: | † anteosauridae |
| Подсемейство: | † syodontinae |
| Род: | † Pampaphoneus Cisneros et al. 2012 |
| Тип видов | |
| † Pampaphoneus biccai Cisneros et al. 2012
| |
Pampaphoneus является вымершим родом плотоядного диноцефальского TherapsId, принадлежащего к семейству Anteosauridae . Он жил от 268 до 265 миллионов лет назад в эпоху Гвадалупианского Фордиана ( в том , = среднего пермского ) периода что сейчас является Бразилии . Pampaphoneus известен почти полным черепом с нижней челюстью, все еще сформулированной, обнаруженной на землях фермы Бокирау, недалеко от города Сан -Габриэль , в штате Рио -Гранд -до Сул . В 2019 и 2020 годах был зарегистрирован второй образец из той же местности, но еще не был описан. Он состоит из черепа, связанного с посткраниальными останками. Это первый южноамериканский вид диномефальянского, который был описан. Группа была ранее известна в Южной Америке только несколькими изолированными зубами, а фрагмент челюсти сообщил в 2000 году в том же регионе Бразилии. циснеросом и проведенный , Филогенетический анализ коллегами Южная Америка и Восточная Европа , чем предполагалось ранее, способствуя реконструкции Continental Pangea B.
Этимология
[ редактировать ]Название рода происходит от Пампаса , области обширных равнин, типичная для Южной Южной Америки, из которой возникает образец, и от греческого телефона , что означает «убийца», в отношении хищных привычек животного. Конкретный эпитет чтит Хосе Бикку, землевладельца фермы, где было найдено ископаемое. [ 1 ]
Описание
[ редактировать ]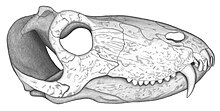
Голотип ( UFRGS PV386P ) PampAphoneus представляет собой череп длиной около 32 см. Как и во всех антеозаврах, вентральная маржа премаксиллы склонен вверх, а посторбитальный стержень сильно изогнут передневентрально, так что временная фенестра подрывает орбиту . Крыша черепа слегка подтолкнула . Временные фенестра широко открываются на крыше черепа , где зона вставки мускулатуры аддуктора в нижней челюсти распространяется на босс пинеще . Фронталы вносят вклад в зону вставки этой мускулатуры, а также участвуют в переднем крае босса шишкол. Эти характеристики указывают на то, что Pampaphoneus принадлежит к Clade Syodontinae . В рамках этой группы это с русскими видами Syodon Biarmicum , что Pampaphoneus разделяет наибольшее количество сходства. Как и последнее, он обладает крепко повторяющимися крючками крючками и очень низкими постканинами со скеррасами. Клыки голотипа имеют длину 7 см. Pampaphoneus , однако, отличается от Syodon его большим размером, более надежной морской, утолщенной Посторбитальный формирование надорбитального босса, и хорошо развитый гребень, простирающийся от босса пинеще до орбитального обода. Он также отличается от других антеозавров своими сквомозальными с помощью югального процесса , простирающегося за пределы переднего края временного фенестра. [ 1 ] Особенности голотипа, который, как считается, был диагностической особенностью, - это наличие всего четыре зуба на предчезгиллу (четвертый резчик , небольшой по размеру, однако, маскируется в боковом направлении ) и восемь постканинов. [ 1 ] Тем не менее, был сообщено второй череп (но неписанный), показывал некоторые различия с голотипом, такими как наличие пяти прачециллярных зубов и только семь постканинов. Поскольку другие черепные элементы очень похожи, эти различия интерпретируются как внутривидовые вариации. [ 2 ] [ 3 ] Несмотря на то, что, будучи Syodontinae, Pampaphoneus также делится несколькими персонажами с более полученными Anteosaurinae . Таким образом, он имеет хорошо развитый медиальный гребень на крыше черепа, в то время как ярко выраженное утолщение каждого посторбитала образует супраорбитальный босс, аналогичный устойчивому человеку из российского титанофона . Кроме того, угловая кость нижней челюсти имеет босса, как у Anteosaurus и двух видов титанофона . Этот угловой босс, однако, гораздо менее развит в Pampaphoneus . [ 1 ]
Палеоэкология
[ редактировать ]
Остатки Pampaphoneus поступают от члена Морро Пеладо формирования Rio Do Rasto , обнаружившись на землях фермы Бокирао в муниципалитете Сан -Габриэля (район Катуаба, штат Рио -Гранди -ду -Сул). Окаменелости сохраняются там в тонком розоватом песчанике и покрыты темной корочкой оксида железа. [ 4 ] Отложения члена Морро Пеладо были отложены в флювио-лакустрированных и дельтовых условиях, предполагающих аллювиальные условия с коалесцирующими поймыми. Эолианские фации (ископаемые дюны), присутствующие в верхней части члена Морро Пеладо, подтверждают прогрессирующую засущную экологию. Палеогеографически, юго-восточная Бразилия была расположена в середине труда между 35-м и 45-м параллельным югом , где теплый умеренный климат, вероятно, преобладал с длительным сухим сезоном. [ 5 ] Флора включала в себя леса Glossopteris Communis , которые занимали поймы и каналы из перевальщиков, в то время как более влажные биотопы были покрыты плотными ковриками эквизетидов, включая Equisetales ( Schizoneura и Phyllotheca ) и Sphenophylles ( Sphenophyllum Paranenes ). [ 5 ] Помимо Pampaphoneus , на ферме Boqueirão также содержится темппондиловая Коньхуковия Sangabrielensis [ 6 ] и маленький дицинодонт Растодон . [ 4 ] Другие населенные пункты члена Морро Пеладо также дали тетрапод останки. В Рио -Гранде -ду -Суле, несколько участков вокруг Aceguá в районе Минуано дали наиболее полные окаменелости Pareiasaur Perosaurus . [ 7 ] Местность фермы Fagundes (Сан -Габриэль, район Катуаба) дала фрагментарный череп Провелозавра , [ 8 ] Зубы неопределенных антеозавридов , титаносучид и тапиносефалидов диноцефалов, [ 9 ] и неопределенная амфибия. [ 10 ] В районе Тиараджу (также недалеко от Сан -Габриэля) было найдено базальное аномодонт Тиараджуденс [ 11 ] [ 12 ] От зоны весла (недалеко от , штат Парана ) Ортигейра , ест амфибийский австралирпетон , [ 10 ] неопределенные виды дицинодонта эндотиодона , [ 13 ] (Возможно, Э. Батистома [ 14 ] ), а также неопределенный тапиноцефалидный диноцефьянец, показывающий сходство с родами мосшопов и Moschognathus . [ 10 ] Тем не менее, существуют неопределенности относительно современности всех этих сайтов. Окаменелости позвоночных из формации Rio Do Rasto происходят в рассеянных, изолированных и прерывистых обнажениях из -за плотного растительного покрова, что затрудняет установление местных корреляций. Точное местоположение нескольких древних открытий является неопределенным, и несколько таксонов могут исходить от разных стратиграфических уровней в формировании Rio Do Rasto. Это случай окаменелостей Парана, где останки Тапиноцефалида и Эндотиона могут исходить от двух разных уровней. [ 10 ]
Возраст в Рио -до Расто Формирование
[ редактировать ]Оценка возраста формирования Rio Do Rasto уже давно полагалась исключительно на основе биостратиграфических корреляций. С 2018 года радиометрическое датирование позволило указать возраст части образования. [ 15 ] [ 16 ] Основываясь на конхостракане и двустворчатых фаунах, Хольц и его коллеги предположили, что осадочная преемственность этой формирования простиралась от Фордиана (среднего Гвадалупия ) до вучиапианского (основания лопинского ). [ 17 ] Среди тетрапод присутствие диномефальцев на участках Фагундес и Бокирао (штат Рио -Гранде -ду -Сул), а также в регионе Серра -Кадеаде (штат Парана) указывает на гвадалупийский век для этих населенных пунктов, учитывая, что эта синапсидная группа ограничен этим возрастом в остальном мире. На участке фермы Fagundes диномефальцы сосуществуют с Pareiasaur Provelosaurus . Последний присутствует в области ацегуеты примерно на 2 м над слоем толщиной 30 см бентонитового радиометрического, датированного 266 ± 5,4 миллиона лет. [ 16 ] [ 7 ] Широкая маржа ошибок в этом радиометрическом знакомстве помещает бразильские диноцефальные участки в промежуточном интервале времени в Нижнем Роудском - Среднем Капитан , что подтверждает гваадалупийский век этой части члена Морро Пеладо. [ 7 ] Другие тетраподы члена Морро Пеладо в Рио -Гранди -ду -Суле предполагают, что возраст, не превышающий Вордиан: Тапиноцефалиды известны только из вордианских и капитанских породов, базальный аномодонт связан южноафриканским аномоцефали с родом тиараджуденов тесно Простирается от покойного Фордиана до позднего капитан, а теммоспондил Конюковия , присутствующая на ферме Бокирао в Сан-Габриэле, также известна в восточной Европейской России в Страте, ограниченном интервалом Вордиан-Капитанского. [ 7 ] Вероятный возраст Ворджина члена Морро Пеладо также усиливается радиометрическим возрастом члена Серринья (нижняя часть формирования Рио -до Расто), полученной из слоя вулканического пепла , который дал возраст 270,61 + 1,76/ - 3,27 млн. Доудж (ранний гуадалупиан). [ 15 ]
Классификация
[ редактировать ]В описании Pampaphoneus , Cisneros et al. Представили несколько кладограмм, подтверждающих признание клад, Anteosaurinae и Syodontinae, установленных годом ранее Кристианом Каммерером. Во всех их анализах Pampaphoneus идентифицируется как самый базальный Syodontinae: [ 1 ]
Палеобиогеография
[ редактировать ]Во время перми большинство сухопутных массовых были объединены в один суперконтинент Pangea , который был примерно C-образной. Его северные ( Лауразию ) и южные ( Гондвана ) части были связаны с западом, но разделены на восток очень большой океанической бухтой - Tethys. [ 18 ] Длинная цепочка микроконтинтов, сгруппированная под именем Cimmeria , разделила тети на два: палео-тети на севере и нео-тетисы на юге. [ 19 ] В северном полушарии антеозавры известны из Восточной Европейской России , Казахстана и Китая , территории, соответствующие средним палеолатитудам между 30-м и 40-м параллельным севером . [ 20 ] В южном полушарии останки антеозавра известны в Южной Африке , Замбии и Зимбабве , территориях, которые расположены в высоких палеолатитудах, расположенных на уровне 60 -го параллельного юга . [ 20 ] [ 21 ] Бразильские населенные пункты находились немного дальше на север, между 35 -м и 45 -м параллельным югом . [ 5 ] Тесные отношения бразильского пампафонея с некоторыми южноафриканскими и российскими антеозаврами предполагают рассеивание через западную Пангею, а не по восточной панге через катайский мост. Последний включал в себя части северного и южного , Кореи и Индокитая и сыграл важную роль в рассеивании терапидов Китая в позднем Пермском и Триасе . Тем не менее, этот мост, вероятно, еще не был сформирован в средней пермской, а морские барьеры разбили различные катайзийские блоки, что затрудняет рассеяние животных из Катайсии в южную Африку через Восточную Европу . Циснерос и его коллеги предполагают, казалось бы, более простой маршрут миграции через Западную Пангею, предпочитая континентальную реконструкцию Пангеи B, где Южная Америка была сопоставлена с Аппалачи и, таким образом, была ближе к России, чем в Пангеи, реконструкция. В последнем горы Аппалачи и Мавританид были бы сложными для перехода. Напротив, в конфигурации Pangea B Бразилия была не только ближе к Восточной Европе, но и единственным горным барьером на этом пути был умеренно высокий европейский Герцинид . Антеозавры мигрировали бы из России (откуда приходят самые старые образцы в южную Африку, проходя через Восточную Европу и Западную Африку (обходя цепочку мавританидов), а затем через Бразилию. [ 1 ] Обнаружение вероятных следов антеозавра на юге Франции , [ N 1 ] который затем был расположен в низкой палеоластоте, на уровне 10-го параллельного севера , также подтверждает эту гипотезу, юго-западная Европа находится в этом миграционном коридоре. [ 22 ] [ 23 ] [ 24 ] [ 25 ]
Примечания
[ редактировать ]- ^ Эти следы, изначально названные Planipes Caudatus и P. brachydactylus , теперь сгруппированы под названием Brontopus antecursor (см. Ссылки Marchetti et al. 2019).
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в дюймовый и фон Cisneros, JC; Абдала, Ф.; Atayman-Güven, S.; Рубидж, BS; Шенгёр, AMC; Schultz, CL (2012). «Плотноядающий диномефан из среднего пермца Бразилии и рассеивания тетрапода в Пангее» (PDF) . Труды Национальной академии наук Соединенных Штатов Америки . 109 (5): 1584–1588. BIBCODE : 2012PNAS..109.1584C . doi : 10.1073/pnas.1115975109 . PMC 3277192 . PMID 22307615 .
- ^ Анильсон Коста Сантос, М.; Bulsing, KP; Сантос, в; Ferraz, JS; Oliveira, N.; Protti, SWL; Dos Santos Rodriguez, LA; Ксавье, пл; Из Кейроза, LC; Scineros, JC; Pinheiro, FL (2019). «Частичный скелет Dinocephalus ожидал Pampapophoneus biccai (Permian of Rio Grande do Sul)» . Труды XXVI Бразильского конгресса палеонтологии. Кампинас: Гало . 2 : 270–271.
- ^ Perval Paz, C.; Анильсон Коста Сантос, М.; Лима Пинхайро, Ф. (2020). «Первая запись Pampophoneus biccai (Anteosauridae) с посткраниальными костями » . Междисциплинарный журнал образования и окружающей среды . 1 (1): 133.
- ^ Jump up to: а беременный Бус, реклама; Kammerer, CF; Schultz, cl; Soares, MB; Ilha, Alr (2016). «Новый дицинодонт (Therapsida: Anomodontia) от Пермия Южной Бразилии и его последствия для бидентало -происхождения» . Plos один . 11 (5): E0155000. BIBCODE : 2016PLOSO..1155000B . doi : 10.1371/journal.pone.0155000 . PMC 4880204 . PMID 27224287 .
- ^ Jump up to: а беременный в Кунео, Н.Р. (1996). «Пермская фитогеография в Гондване». Палеогеография, палеоклиматология, палеоэкология . 125 (1–4): 75–104. Bibcode : 1996ppp ... 125 ... 75c . doi : 10.1016/s0031-0182 (96) 00025-9 .
- ^ Pacheco, CP; Eltink, E.; Мюллер, RT; Dias-Silva, In (2016). "Новая семья Аркесариды" Журнал . 15 (3): 241–2 doi : 10.1080/ 1472019.2016.1164763 S2CID 878600271 .
- ^ Jump up to: а беременный в дюймовый Cisneros, JC; Dentzien-Dias, P.; Фрэнсисчини, Х. (2021). «Бразильский пареайайавр пересмотрел» . Границы в экологии и эволюции . 9 : 758802. DOI : 10.3389/fevo.2021.758802 .
- ^ Cisneros, JC; Абдала, Ф.; Малабарба, MC (2005). «Pareiaurids из Rio Do Rasto Formation, Южная Бразилия: биостративные последствия для пермских фаун бассейна Парана » Revista Basileira de Palontology 8 (1): 13–2 Doi : 10.4072/ rbp.2005.1.0
- ^ Лангер, MC (2000). «Первая запись диномефальцев в Южной Америке: поздний Пермский (Rio Do Rasto Formation) бассейна Paraná, Бразилия». Новый ежегодный книга по геологии и палеонтологии - трактаты . 215 : 69–95. Doi : 10.1127/njgpa/215/2000/69 .
- ^ Jump up to: а беременный в дюймовый Бус, реклама; Kammerer, CF; Schultz, cl; Paes Neto, VD (2015). "Бассейн Параса, Бразилия): Американская земля 63 : 375–3 Bibcode : 2015jsaes . doi : 10.1016/j.jsames .
- ^ Cisneros, JC; Абдала, Ф.; Рубидж, BS; Dentzien-Dias, PC; Буэно, АО (2011). «Зубная окклюзия у 260-миллионного летнего терапида с саблями клыков из Пермия Бразилии». Наука . 331 (6024): 1603–1605. Bibcode : 2011sci ... 331.1603c . doi : 10.1126/science.1200305 . PMID 21436452 . S2CID 8178585 .
- ^ Cisneros, JC; Абдала, Ф.; Jashashvili, T.; Буэно, АО; Dentzien-Dias, P. (2015). « Tiarajudens Excentricus и Anomocephalus Africanus , два странных аномодонта (Synapsida, Therapsida) с зубной окклюзией от Пермия Гондвана» . Королевское общество открыто наука . 2 (7): 1–27. Bibcode : 2015rsos .... 250090c . doi : 10.1098/rsos.150090 . PMC 4632579 . PMID 26587266 .
- ^ Бус, реклама; Schultz, cl; Вега, CS; Aumond, JJ (2013). «О присутствии позднего Пермского Дицинодонта Эндотиодона в Бразилии». Палеонтология . 56 (4): 837–848. doi : 10.1111/pala.12020 . S2CID 129679835 .
- ^ Kammerer, CF; Ордоньес, Массачусетс (2021). «Дицинодонты (Therapsida: Anomodontia) Южной Америки». Журнал южноамериканских наук о Земле . 108 : 103171. Bibcode : 2021jsaes.10803171K . doi : 10.1016/j.jsames.2021.103171 . S2CID 233565963 .
- ^ Jump up to: а беременный Francischini, H.; Dentzien-Dias, P.; Guerra-Sommer, M.; Менегат, Р.; Сантос, Джос; Manfroi, J.; Schultz, CL (2018). «Средняя пермская (дорожная) эстивация легких рыбков из формации Рио-до Расто (бассейн Парана, Бразилия) и связанные с ними знакомства с U-PB». Палаис . 33 (2): 69–84. Bibcode : 2018palai..33 ... 69f . doi : 10.2110/palo.2017.050 . S2CID 134435628 .
- ^ Jump up to: а беременный Rocha-Campos, AC; Basei, mas; Натман, AP; Santos, Pr; Пассарелли, кр .; Canile, FM; Роза, OCR; Фернандес, MT; Санта -Ана, Х.; Верославский, Г. (2019). «U-PB Циркон датируется отложениями пепла из палеозойского бассейна Парана в Бразилии и Уругвае: переоценка стратиграфических корреляций». Журнал геологии . 127 (2): 167–182. Bibcode : 2019jg .... 127..167r . doi : 10.1086/701254 . HDL : 1885/202604 . S2CID 134671451 .
- ^ Holz, M.; Франса, AB; Souza, PA; Яннуцци, Р.; Рон Р. (2010). «Стратиграфическая карта поздней каменноугольной/пермской преемственности восточной границы бассейна Парана, Бразилия, Южная Америка». Журнал южноамериканских наук о Земле . 29 (2): 381–399. Bibcode : 2010jsaes..29..381h . doi : 10.1016/j.jsames.2009.04.004 .
- ^ McLoughlin, S. (2001). «История разрыва Гондваны и ее влияние на пре-кузозойский флористический провинциализм». Австралийский журнал ботаники . 49 (3): 271–300. doi : 10.1071/bt00023 .
- ^ Шенгёр, AMC (1987). «Тектоника тетизидов: орогенное развитие коллажа в столкновении». Ежегодный обзор земли и планетарных наук . 15 : 214–244. Bibcode : 1987Areps..15..213c . doi : 10.1146/annurev.ea.15.050187.001241 .
- ^ Jump up to: а беременный Кемп, Т.С. (2006). «Происхождение и раннее излучение рептилий, подобных млекопитающим,: палеобиологическая гипотеза» . Журнал эволюционной биологии . 19 (4): 1231–1247. doi : 10.1111/j.1420-9101.2005.01076.x . PMID 16780524 . S2CID 3184629 .
- ^ Bamford, MK (2016). «Ископаемые леса от верхнего карбона до нижнего бассейна Юрского периода Кару и их интерпретации окружающей среды». В Линоле, б.; де Вит, MJ (ред.). Происхождение и эволюция горов мыса и бассейна Кару . CHAM: региональные обзоры геологии, Springer. С. 159–167. ISBN 978-3-319-40858-3 .
- ^ Ганд, Г.; Завтра, Г.; Крестобал Ф. (1995). «Палихнофауна тетрапод верхней части Эстереля (Прованс, Франция)» Abteilung palaeontography a 4–6 (4–6): 97–139. два 10.1127/pala/235/1995/97: 247392880S2CID
- ^ Ганд, Г.; Гаррик, Дж.; Завтра, Г.; Элленбергер, П. (2000). «Палихнофауна тетрапод верхнего разрешения бассейна Лодева (Лангедок-Франс) » -переживание Палайер 29 (1): 1–82.
- ^ Marchetti, L.; Klein, H.; Бухвиц, М.; Рончи, А.; Смит, RMH; Де Клерк, WJ; Sciscio, L.; Groenewald, GH (2019). «Пермские триасовые следы позвоночных из Южной Африки: Ichnotaxonomy, продюсеры и биостратиграфия через два основных кризиса фауны» . Gondwana Research . 72 : 139–168. Bibcode : 2019gondr..72..139M . doi : 10.1016/j.gr.2019.03.009 . S2CID 133781923 .
- ^ Olroyd, Sl; Сидор, Калифорния (2017). «Обзор Гвадалупийской (средней пермской) глобальной тетрапод ископаемых записей» . Земля-наука обзоров . 171 : 583–597. Bibcode : 2017esrv..171..583o . doi : 10.1016/j.earscirev.2017.07.001 .

